
Extended Sentinel Monitoring of Helicoverpa zea Resistance to Cry and Vip3Aa Toxins in Bt Sweet Corn: Assessing Changes in Phenotypic and Allele Frequencies of Resistance
 , , , , ,
, , , , ,  , , , , , , , , , , , , , , add
Show full author list
, , , , , , , , , , , , , , add
Show full author list
Abstract
:Simple Summary
Abstract
1. Introduction
2. Materials and Methods
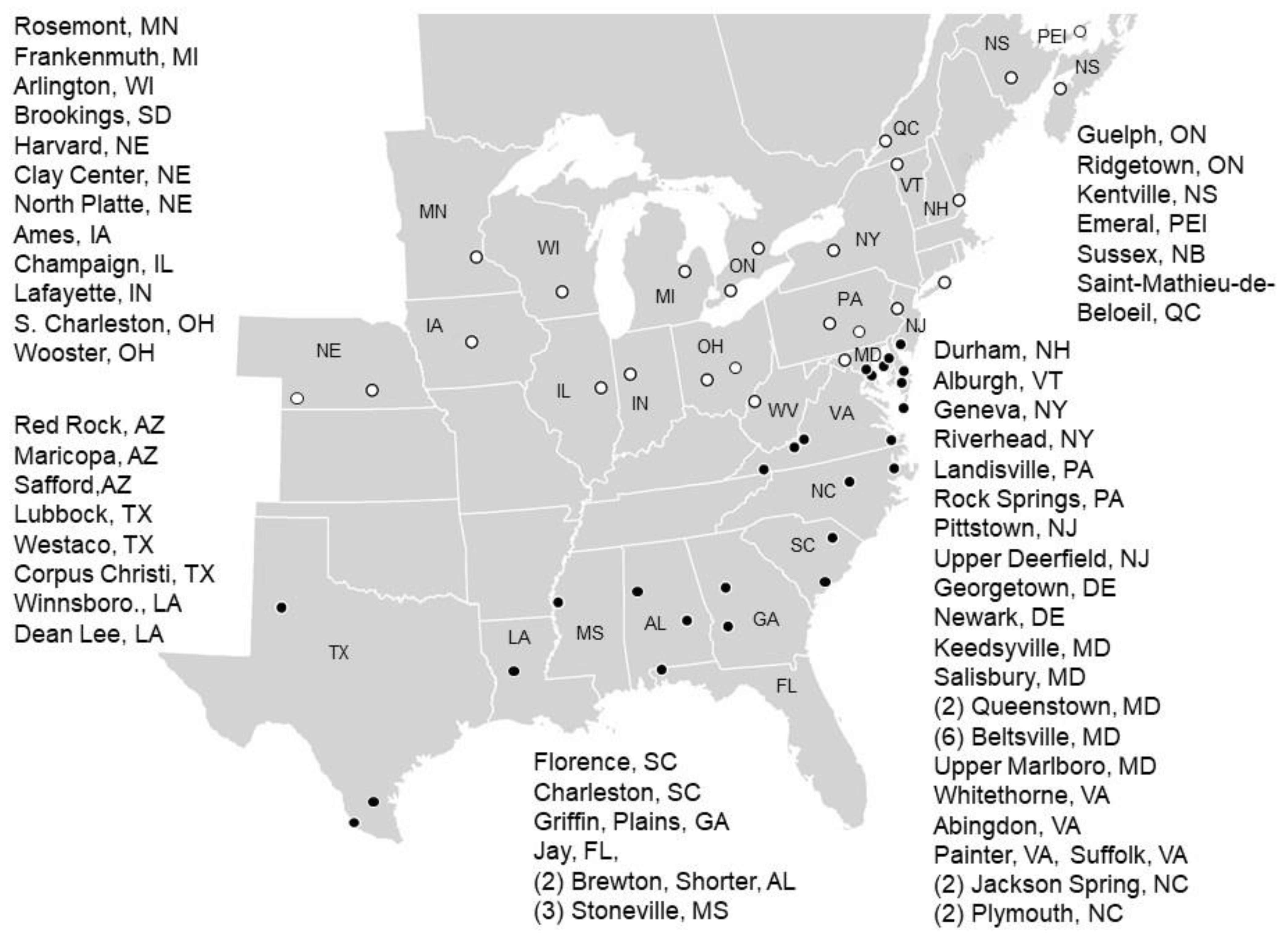
2.1. Sentinel Trial Locations
2.2. Hybrids, Planting Arrangement, and Plot Size
2.3. Ear Sampling
2.4. Recorded Data
2.5. Data Adjustments and Analysis
3. Results and Discussion
3.1. Occurrence and Infestation Levels of the Major Lepidopteran Larvae
3.2. Ear Damage, Larval Development, and Kernel Consumption by H. zea
3.3. Phenotypic Resistance and Estimated Range of Resistance Allele Frequency
4. Conclusions
Supplementary Materials
Author Contributions
Funding
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- USDA-ERS (U.S. Department of Agriculture, Economic Research Service). Adoption of Genetically Engineered Crops in the U.S. Recent Trends in GE Adoption. 2022. Available online: https://www.ers.usda.gov/data-products/adoption-of-genetically-engineered-crops-in-the-u-s/recent-trends-in-ge-adoption/ (accessed on 1 April 2023).
- Carrière, Y.; Ellers-Kirk, C.; Sisterson, M.; Antilla, L.; Whitlow, M.; Dennehy, T.J.; Tabashnik, B.E. Long-term regional suppression of pink bollworm by Bacillus thuringiensis cotton. Proc. Natl. Acad. Sci. USA 2003, 100, 1519–1523. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wu, K.M.; Lu, Y.H.; Feng, H.Q.; Jiang, Y.Y.; Zhao, J.Z. Suppression of cotton bollworm in multiple crops in China in areas with Bt toxin containing cotton. Science 2008, 321, 1676–1678. [Google Scholar] [CrossRef] [Green Version]
- Hutchison, W.D.; Burkness, E.C.; Mitchell, P.D.; Moon, R.D.; Leslie, T.W.; Fleischer, S.J.; Abrahamson, M.; Hamilton, K.L.; Steffey, K.L.; Gray, M.E.; et al. Areawide suppression of European corn borer with Bt maize reaps savings to non-Bt maize growers. Science 2010, 330, 222–225. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lu, Y.; Wu, K.; Jiang, Y.; Guo, Y.; Desneux, N. Widespread adoption of Bt cotton and insecticide decrease promotes biocontrol services. Nature 2012, 487, 362–365. [Google Scholar] [CrossRef] [PubMed]
- Edgerton, M.D.; Fridgen, J.; Anderson, J.R., Jr.; Ahlgrim, J.; Criswell, M.; Dhungana, P.; Gocken, T.; Li, Z.; Mariappan, S.; Pilcher, C.D.; et al. Transgenic insect resistance traits increase corn yield and yield stability. Nat. Biotechnol. 2012, 30, 493–496. [Google Scholar] [CrossRef] [PubMed]
- Klümper, W.; Qaim, M.A. Meta-analysis of the impacts of genetically modified crops. PLoS ONE 2014, 9, e111629. [Google Scholar] [CrossRef] [Green Version]
- Chen, G.; Hooks, C.R.R.; Patton, T.W.; Dively, G.P. Tolerance to stalk and ear-invading worms and yield performance of Bt and conventional corn hybrids. Agron. J. 2016, 108, 73–84. [Google Scholar] [CrossRef]
- Perry, E.D.; Ciliberto, F.; Hennessy, D.A.; Moschini, G. Genetically engineered crops and pesticide use in U.S. maize and soybeans. Sci. Adv. 2016, 2, e1600850. [Google Scholar] [CrossRef] [Green Version]
- Dively, G.P.; Venugopal, P.D.; Bean, D.; Whalen, J.; Holmstrom, K.; Kuhar, T.P.; Doughty, H.B.; Patton, T.; Cissel, W.; Hutchison, W.D. Regional pest suppression associated with widespread Bt maize adoption benefits vegetable growers. PNAS 2018, 115, 3320–3325. [Google Scholar] [CrossRef] [Green Version]
- Romeis, J.; Naranjo, S.E.; Meissle, M.; Shelton, A.M. Genetically engineered crops help support conservation biological control. Biol. Control. 2019, 130, 136–154. [Google Scholar]
- USEPA (U.S. Environmental Protection Agency). The Environmental Protection Agency’s White Paper on Bt Plant-Pesticide Resistance Management. 1998. Available online: https://nepis.epa.gov/Exe/ZyPURL.cgi?Dockey=20000TQB.txt (accessed on 2 April 2023).
- USEPA (U.S. Environmental Protection. Agency). Plant-Incorporated Protectants, Insect Resistance Management, 15 October, Biopesticides Registration Action Document. 2001. Available online: https://www3.epa.gov/pesticides/chem_search/reg_actions/pip/bt_brad2/4-irm.pdf (accessed on 2 April 2023).
- Alstad, D.; Andow, D.A. Managing the evolution of insect resistance to transgenic plants. Science 1995, 268, 1894–1896. [Google Scholar] [CrossRef] [PubMed]
- Roush, R.T. Bt-transgenic crops: Just another pretty insecticide or a chance for a new start in resistance management? Pestic. Sci. 1997, 51, 328–334. [Google Scholar] [CrossRef]
- Gould, F. Sustainability of transgenic insecticidal cultivars: Integrating pest genetics and ecology. Annu. Rev. Entomol. 1998, 43, 701–726. [Google Scholar] [CrossRef] [Green Version]
- Roush, R.T. Two-toxin strategies for management of insecticidal transgenic crops: Can pyramiding succeed where pesticide mixtures have not? Philos. Trans. R. Soc. Lond. 1998, 353, 1777–1786. [Google Scholar] [CrossRef] [Green Version]
- Carrière, Y.; Crickmore, N.; Tabashnik, B.E. Optimizing pyramided transgenic Bt crops for sustainable pest management. Nat. Biotechnol. 2015, 33, 161–168. [Google Scholar] [CrossRef]
- Santiago-González, J.C.; Kerns, D.L.; Head, G.P.; Yang, F. Effective dominance and redundant killing of single- and dual-gene resistant populations of Helicoverpa zea on pyramided Bt corn and cotton. Pest Manag. Sci. 2022, 78, 4333–4339. [Google Scholar] [CrossRef]
- Head, G.P.; Greenplate, J. The design and implementation of insect resistance management programs for Bt crops. GM Crop. Food 2012, 3, 144–153. [Google Scholar] [CrossRef] [Green Version]
- Tabashnik, B.E.; Mota-Sanchez, D.; Whalon, M.E.R.; Hollingworth, M.; Carrière, Y. Defining terms for proactive management of resistance to Bt crops and pesticides. J. Econ. Entomol. 2014, 107, 496–507. [Google Scholar] [CrossRef] [Green Version]
- Smith, J.L.; Farhan, Y.; Schaafsma, A.W. Practical Resistance of Ostrinia nubilalis (Lepidoptera: Crambidae) to Cry1F Bacillus thuringiensis maize discovered in Nova Scotia, Canada. Sci. Rep. 2019, 9, 18247. [Google Scholar] [CrossRef] [Green Version]
- Smith, J.L.; Farhan, Y. Monitoring resistance of Ostrinia nubilalis (Lepidoptera: Crambidae) in Canada to Cry toxins produced by Bt corn. J. Econ. Entomol. 2023, 116, toad046. [Google Scholar] [CrossRef]
- Huang, F.; Qureshi, J.A.; Meagher, R.L., Jr.; Reisig, D.D.; Head, G.P.; Andow, D.A.; Ni, X.Z.; Kerns, D.; Buntin, G.D.; Niu, Y.; et al. Cry1F resistance in fall armyworm Spodoptera frugiperda: Single gene versus pyramided Bt maize. PLoS ONE 2014, 9, e112958. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hardee, D.D.; Adams, L.C.; Solomon, W.L.; Sumerford, D.V. Tolerance to Cry1Ac in populations of Helicoverpa zea and Heliothis virescens (Lepidoptera: Noctuidae): Three-year summary. J. Agric. Urban Entomol. 2001, 18, 187–197. [Google Scholar]
- Ali, M.I.; Luttrell, R.G.; Abel, C. Monitoring Bt susceptibilities in Helicoverpa zea and Heliothis virescens: Results of 2006 studies. In Proceedings of the 2007 Beltwide Cotton Conferences, New Orleans, LA, USA, 11 January 2007; pp. 1062–1072. [Google Scholar]
- Ali, M.I.; Luttrell, R.G. Susceptibilities of bollworm and tobacco budworm (Lepidoptera: Noctuidae) to Cry2Ab2 insecticidal protein. J. Econ. Entomol. 2007, 100, 921–931. [Google Scholar] [CrossRef] [PubMed]
- Reisig, D.D.; Reay-Jones, F.P.F. Inhibition of Helicoverpa zea (Lepidoptera: Noctuidae) growth by transgenic corn expressing Bt toxins and development of resistance to Cry1Ab. Environ. Entomol. 2015, 44, 1275–1285. [Google Scholar] [CrossRef]
- Dively, G.P.; Venugopal, P.D.; Finkenbinder, C. Field-evolved resistance in corn earworm to Cry proteins expressed by transgenic sweet corn. PLoS ONE 2016, 11, e0169115. [Google Scholar] [CrossRef] [Green Version]
- Tabashnik, B.E.; Carrière, Y. Surge in insect resistance to transgenic crops and prospects for sustainability. Nat. Biotechnol. 2017, 35, 926–935. [Google Scholar] [CrossRef]
- Reisig, D.D.; Huseth, A.S.; Bacheler, J.S.; Aghaee, M.-A.; Braswell, L.; Burrack, H.J.; Flanders, K.; Greene, J.K.; Herbert, D.A.; Jacobson, A.; et al. Long-term empirical and observational evidence of practical Helicoverpa zea resistance to cotton with pyramided Bt toxins. J. Econ. Entomol. 2018, 111, 1824–1833. [Google Scholar] [CrossRef]
- Bilbo, T.R.; Reay-Jones, F.P.F.; Reisig, D.D.; Greene, J.K. Susceptibility of corn earworm (Lepidoptera: Noctuidae) to Cry1A.105 and Cry2Ab2 in North and South Carolina. J. Econ. Entomol. 2019, 112, 1845–1857. [Google Scholar] [CrossRef]
- Kaur, G.; Guo, J.G.; Brown, S.; Head, G.P.; Price, P.A.; Paula-Moraes, S.; Ni, X.Z.; Dimase, M.; Huang, F. Field-evolved resistance of Helicoverpa zea (Boddie) to transgenic maize expressing pyramided Cry1A.105/Cry2Ab2 proteins in northeast Louisiana, the United States. J. Invertebr. Pathol. 2019, 163, 11–20. [Google Scholar] [CrossRef]
- Dively, G.; Kuhar, T.; Taylor, S.; Doughty, H.; Holmstrom, K.; Gilrein, D.; Nault, B.; Ingerson-Mahar, J.; Whalen, J.; Reisig, D.; et al. Sweet corn sentinel monitoring for lepidopteran field-evolved resistance to Bt toxins. J. Econ. Entomol. 2021, 114, 307–319. [Google Scholar] [CrossRef]
- Yang, F.; Santiago González, J.C.; Head, G.P.; Price, P.A.; Kerns, D.L. Multiple and non-recessive resistance to Bt proteins in a Cry2Ab2-resistant population of Helicoverpa zea. Crop Prot. 2021, 145, 105650. [Google Scholar] [CrossRef]
- Yu, W.; Head, G.P.; Huang, F. Inheritance of Resistance to Cry1A.105 in Helicoverpa zea (Boddie) (Lepidoptera: Noctuidae). Academic 2022, 13, 875. [Google Scholar] [CrossRef]
- Lin, S.; Oyediran, I.; Niu, Y.; Brown, S.; Cook, D.; Ni, X.; Zhang, Y.; Reay-Jones, F.P.F.; Chen, J.S.; Wen, Z.; et al. Resistance allele frequency to Cry1Ab and Vip3Aa20 in Helicoverpa zea (Boddie) (Lepidoptera: Noctuidae) in Louisiana and three other Southeastern U.S. states. Toxins 2022, 14, 270. [Google Scholar] [CrossRef]
- Santiago-González, J.C.; Kerns, D.L.; Head, G.P.; Yang, F. A modified F2 screen for estimating Cry1Ac and Cry2Ab resistance allele frequencies in Helicoverpa zea (Lepidoptera: Noctuidae). J. Econ. Entomol. 2023, 116, 289–296. [Google Scholar] [CrossRef] [PubMed]
- Tabashnik, B.E.; Fabrick, J.A.; Carrière, Y. Global patterns of insect resistance to transgenic Bt crops: The first 25 years. J. Econ. Entomol. 2023, 116, 297–309. [Google Scholar] [CrossRef] [PubMed]
- Brévault, T.; Heuberger, S.; Zhang, M.; Ellers-Kirk, C.; Ni, X.; Masson, L.; Li, X.; Tabashnik, B.E.; Carrière, Y. Potential shortfall of pyramided transgenic cotton for insect resistance management. Proc. Natl. Acad. Sci. USA 2013, 110, 5806–5811. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Carrière, Y.; Degain, B.; Unnithan, G.C.; Harpold, V.S.; Li, X.; Tabashnik, B.E. Seasonal declines in Cry1Ac and Cry2Ab concentration in maturing cotton favor faster evolution of resistance to pyramided Bt cotton in Helicoverpa zea. J. Econ. Entomol. 2019, 112, 2907–2914. [Google Scholar] [CrossRef] [PubMed]
- Carrière, Y.; Degain, B.A.; Harpold, V.S.; Unnithan, G.C.; Tabashnik, B.E. Gene flow between Bt and non-Bt plants in a seed mixture increases dominance of resistance to pyramided Bt corn in Helicoverpa zea. J. Econ. Entomol. 2020, 113, 2041–2051. [Google Scholar] [CrossRef]
- Reisig, D.D. Factors associated with willingness to plant non-Bt maize refuge and suggestions for increasing refuge compliance. J. Integr. Pest Manag. 2017, 8, 9. [Google Scholar] [CrossRef] [Green Version]
- Reisig, D.D.; Kurtz, R. Bt resistance implications for Helicoverpa zea (Lepidoptera: Noctuidae) insecticide resistance management in the United States. Environ. Entomol. 2018, 47, 1357–1364. [Google Scholar] [CrossRef] [Green Version]
- US-EPA (U.S. Environmental Protection Agency). White Paper on Resistance in Lepidopteran Pests of Bacillus Thuringiensis (Bt) Plant-Incorporated Protectants in the United States. 2018. Available online: https://www.epa.gov/sites/production/files/2018-07/documents/position_paper_07132018.pdf (accessed on 2 April 2023).
- Reisig, D.D.; DiFonzo, C.; Dively, G.; Farhan, Y.; Gore, J.; Smith, J. Best management practices to delay the evolution of Bt resistance in lepidopteran pests without high susceptibility to Bt toxins in North America. J. Econ. Entomol. 2021, 115, 10–25. [Google Scholar] [CrossRef] [PubMed]
- Welch, K.L.; Unnithan, G.C.; Degain, B.A.; Wei, J.; Zhang, J.; Li, X.; Tabashnik, B.E.; Carrière, Y. Cross-resistance to toxins used in pyramided Bt crops and resistance to Bt sprays in Helicoverpa zea. J. Invertebr. Pathol. 2015, 132, 149–156. [Google Scholar] [CrossRef] [PubMed]
- Burkness, E.C.; Dively, G.; Patton, T.; Morey, A.C.; Hutchison, W.D. Novel Vip3A Bacillus thuringiensis (Bt) maize approaches high-dose efficacy against Helicoverpa zea (Lepidoptera: Noctuidae) under field conditions: Implications for resistance management. GM Crop. 2010, 1, 337–343. [Google Scholar] [CrossRef] [PubMed]
- Yang, F.; Kerns, D.L.; Little, N.; Brown, S.A.; Stewart, S.D.; Catchot, A.L.; Cook, D.R.; Gore, J.; Crow, W.D.; Lorenz, G.M.; et al. Practical resistance to Cry toxins and efficacy of Vip3Aa in Bt cotton against Helicoverpa zea. Pest Manag. Sci. 2022, 78, 5234–5242. [Google Scholar] [CrossRef] [PubMed]
- Yang, F.; Santiago González, J.C.; Williams, J.; Cook, D.C.; Gilreath, R.T.; Kerns, D.L. Occurrence and ear damage of Helicoverpa zea on transgenic Bacillus thuringiensis maize in the field in Texas, U.S. and Its Susceptibility to Vip3A Protein. Toxins 2019, 11, 102. [Google Scholar] [CrossRef] [Green Version]
- Yang, F.; Santiago González, J.C.; Little, N.; Reisig, D.; Payne, G.; Dos Santos, R.F.; Jurat-Fuentes, J.L.; Kurtz, R.; Kerns, D.L. First documentation of major Vip3Aa resistance alleles in field populations of Helicoverpa zea (Boddie) (Lepidoptera: Noctuidae) in Texas, USA. Sci. Rep. 2020, 10, 5867. [Google Scholar] [CrossRef] [Green Version]
- Yang, F.; Kerns, D.L.; Little, N.S.; Santiago González, J.C.; Tabashnik, B.E. Early Warning of Resistance to Bt Toxin Vip3Aa in Helicoverpa zea. Toxins 2021, 13, 618. [Google Scholar] [CrossRef]
- Santiago-González, J.C.; Kerns, D.L.; Yang, F. Resistance Allele Frequency of Helicoverpa zea to Vip3Aa Bacillus thuringiensis Protein in the Southeastern U.S. Insects 2023, 14, 161. [Google Scholar] [CrossRef]
- Huang, F. Detection and monitoring of insect resistance to transgenic Bt crops. Insect Sci. 2006, 13, 73–84. [Google Scholar] [CrossRef]
- Fritz, M.L. Utility and challenges of using whole genome resequencing to detect emerging insect and mite resistance in agro-ecosystems. Evol. Appl. 2022, 15, 1505–1520. [Google Scholar] [CrossRef]
- Hawthorne, D.; Siegfried, B.; Shelton, T.; Hellmich, R. Monitoring for Resistance Alleles: A Report from an Advisory Panel on Insect Resistance Monitoring Methods for Bt Corn. In Agricultural Biotechnology Stewardship Committee Report; Agricultural Biotechnology Stewardship Technical Committee: Washington, DC, USA, 2001. [Google Scholar]
- US-EPA (U.S. Environmental Protection Agency). EPA Needs Better Data, Plans and Tools to Manage Insect Resistance to Genetically Engineered Corn; 16-P-0194; Office of the Inspector General: Washington, DC, USA, 2016; p. 22. [Google Scholar]
- US-EPA (U.S. Environmental Protection Agency). Resistance in Lepidopteran Pests to Bacillus Thuringiensis (Bt) Plant Incorporated Protectants (PIPs) in the United States. In July 2018 Scientific Advisory Panel; U.S. Environmental Protection Agency: Washington, DC, USA, 2018. Available online: https://www.regulations.gov/document?D=EPA-HQ-OPP-2017-0617-0078 (accessed on 2 April 2023).
- USEPA (U.S. Environmental Protection Agency). EPA Draft Proposal to Address Resistance Risks to Lepidopteran Pests of Bt Following the July 2018 FIFRA Scientific Advisory Panel Recommendation. Memorandum EPA-HQ-OPP-2019-0682-0007. 2020. Available online: https://www.regulations.gov/document?D=EPA-HQ-OPP-2019-0682-0007 (accessed on 2 April 2023).
- Venette, R.C.; Hutchison, W.D.; Andow, D.A. An in-field screen for early detection and monitoring of insect resistance to Bacillus thuringiensis in transgenic crops. J. Econ. Entomol. 2000, 93, 1055–1064. [Google Scholar] [CrossRef] [PubMed]
- Moar, W.; Roush, R.; Shelton, A.; Ferré, J.; MacIntosh, S.; Leonard, B.R.; Abel, C. Field-evolved resistance to Bt toxins. Nat. Biotechnol. 2008, 26, 1072–1074. [Google Scholar] [CrossRef] [PubMed]
- Jurat-Fuentes, J.L.; Heckel, D.G.; Ferré, J. Mechanisms of resistance to insecticidal proteins from Bacillus thuringiensis. Annu. Rev. Entomol. 2021, 66, 121–140. [Google Scholar] [CrossRef] [PubMed]
- Yang, F.; Santiago González, J.C.; Sword, G.A.; Kerns, D.L. Genetic basis of resistance to the Vip3Aa Bt protein in Helicoverpa zea. Pest Manag. Sci. 2021, 77, 1530–1535. [Google Scholar] [CrossRef]
- Siegfried, B.D.; Rangasamy, M.; Wang, H.; Spencer, T.; Haridas, C.V.; Tenhumberg, B.; Sumerford, D.V.; Storer, N.P. Estimating the frequency of Cry1F resistance in field populations of the European corn borer (Lepidoptera: Crambidae). Pest Manag. Sci. 2014, 70, 725–733. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Andow, D.A.; Olson, D.M.; Hellmich, R.L.; Alstad, D.N.; Hutchison, W.D. Frequency of resistance to Bacillus thuringiensis toxin Cry1Ab in an Iowa population of European corn borer (Lepidoptera: Crambidae). J. Econ. Entomol. 2000, 93, 26–30. [Google Scholar] [CrossRef] [Green Version]
- Siegfried, B.D.; Hellmich, R.L. Understanding successful resistance management: The European corn borer and Bt corn in the United States. GM Crop. Food 2012, 3, 184–193. [Google Scholar] [CrossRef] [Green Version]
- Gassmann, A.J.; Reisig, D.D. Management of insect pests with Bt crops in the United States. Annu. Rev. Entomol. 2023, 68, 31–49. [Google Scholar] [CrossRef]
- Horner, T.A.; Dively, G.P.; Herbert, D.A. Development, survival and fitness performance of Helicoverpa zea (Lepidoptera: Noctuidae) in MON810 Bt field corn. J. Econ. Entomol. 2003, 96, 914–924. [Google Scholar] [CrossRef] [PubMed]
- Lynch, R.E.; Wiseman, B.R.; Sumner, H.R.; Plaisted, D.; Warnick, D. Management of corn earworm and fall armyworm (Lepidoptera: Noctuidae) injury on a sweet corn hybrid expressing a cryIA(b) gene. J. Econ. Entomol. 1999, 92, 1217–1222. [Google Scholar] [CrossRef]
- Siegfried, B.D.; Spencer, T.; Nearman, J. Baseline susceptibility of the corn earworm (Lepidoptera: Noctuidae) to the Cry1Ab toxin from Bacillus thuringiensis. J. Econ. Entomol. 2000, 93, 265–1268. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Buntin, G.D.; Flanders, K.L.; Lynch, R.E. Assessment of experimental Bt events against fall armyworm and corn earworm in field corn. J. Econ. Entomol. 2004, 97, 259–264. [Google Scholar] [CrossRef] [PubMed]
- Pan, Z.; Onstad, D.; Crain, P.; Crespo, A.; Hutchison, W.; Buntin, D.; Porter, P.; Catchot, A.; Cook, D.; Pilcher, C.; et al. Evolution of Resistance by Helicoverpa zea (Lepidoptera: Noctuidae) Infesting Insecticidal Crops in the Southern United States. J. Econ. Entomol. 2016, 109, 821–831. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Yu, W.; Lin, S.; Dimase, M.; Niu, Y.; Brown, S.; Head, G.P.; Price, P.A.; Reay-Jones, F.P.F.; Cook, D.; Reisig, D.; et al. Extended investigation of field-evolved resistance of the corn earworm Helicoverpa zea (Lepidoptera: Noctuidae) to Bacillus thuringiensis Cry1A.105 and Cry2Ab2 proteins in the southeastern United States. J. Invertebr. Path. 2021, 183, 107560. [Google Scholar] [CrossRef]
- Niu, Y.; Oyediran, I.; Yu, W.; Lin, S.; Dimase, M.; Brown, S.; Reay-Jones, F.P.; Cook, D.; Reisig, D.; Thrash, B.; et al. Populations of Helicoverpa zea (Boddie) in the Southeastern United States are Commonly Resistant to Cry1Ab, but Still Susceptible to Vip3Aa20 Expressed in MIR 162 Corn. Toxins 2021, 13, 63. [Google Scholar] [CrossRef]
- Storer, N.P.; Gould, F.; Kennedy, G.G.; Van Duyn, J.W. Ecology and biology of cotton bollworm in reference to modeling Bt resistance development in a Bt cotton/Bt corn system. In Proceedings of the 1999 Beltwide Cotton Conference, Orlando, FL, USA, 3–7 January 1999; National Cotton Council of America: New Orleans, LA, USA; pp. 949–952. [Google Scholar]
- Horner, T.A.; Dively, G.P. Effect of MON810 Bt field corn on Helicoverpa zea (Lepidoptera: Noctuidae) cannibalism and its implications to resistance development. J. Econ. Entomol. 2003, 96, 931–934. [Google Scholar] [CrossRef]
- Chilcutt, C.F. Cannibalism of Helicoverpa zea (Lepidoptera: Noctuidae) from Bacillus thuringiensis (Bt) transgenic corn versus non-Bt corn. J. Econ. Entomol. 2006, 99, 728–732. [Google Scholar] [CrossRef] [Green Version]
- Wei, J.; Guo, Y.; Liang, G.; Wu, K.; Zhang, J.; Tabashnik, B.E.; Li, X. Cross-resistance and interactions between Bt toxins Cry1Ac and Cry2Ab against the cotton bollworm. Sci. Rep. 2015, 5, 7714. [Google Scholar] [CrossRef] [Green Version]
- US-EPA (U.S. Environmental Protection Agency). Biopesticide Registration Action Document: Bacillus Thuringiensis Cry1A.105 and Cry2Ab2 Insecticidal Proteins and the Genetic Material Necessary for Their Production in Corn. 2010. Available online: https://www3.epa.gov/pesticides/chem_search/reg_actions/pip/mon-89034-brad.pdf (accessed on 2 April 2023).
- Siebert, M.W.; Nolting, S.P.; Hendrix, W.H.; Dhavala, S.; Craig, C.; Leonard, B.R.; Stewart, S.D.; All, J.; Musser, F.; Buntin, G.D.; et al. Evaluation of corn hybrids expressing Cry1F, Cry1A.105, Cry2Ab2, Cry34Ab1/Cry35Ab1, and Cry3Bb1 against southern United States insect pests. J. Econ. Entomol. 2012, 105, 1825–1834. [Google Scholar] [CrossRef]
- Rule, D.M.; Nolting, S.P.; Prasifka, P.L.; Storer, N.P.; Hopkins, B.W.; Scherder, E.F.; Siebert, M.W.; Hendrix, W.H., III. Efficacy of pyramided Bt proteins Cry1F, Cry1A.105, and Cry2Ab2 expressed in SmartStax corn hybrids against lepidopteran insect pests in the northern United States. J. Econ. Entomol. 2014, 107, 403–409. [Google Scholar] [CrossRef] [Green Version]
- Yang, F.; Head, G.P.; Price, P.A.; Santiago González, J.C.; Kerns, D.L. Inheritance of Bacillus thuringiensis Cry2Ab2 protein resistance in Helicoverpa zea (Lepidoptera: Noctuidae). Pest Manag. Sci. 2020, 76, 3676–3684. [Google Scholar] [CrossRef] [PubMed]
- Arends, B.R.; Reisig, D.D.; Gundry, S.; Greene, J.K.; Kennedy, G.G.; Reay-Jones, F.P.F.; Huseth, A.S. Helicoverpa zea (Lepidoptera: Noctuidae) feeding incidence and survival on Bt maize in relation to maize in the landscape. Pest Manag. Sci. 2022, 78, 2309–2315. [Google Scholar] [CrossRef] [PubMed]
- Reisig, D.; Buntin, G.D.; Greene, J.K.; Paula-Moraes, S.V.; Reay-Jones, F.; Roberts, P.; Smith, R.; Taylor, S.V. Magnitude and Extent of Helicoverpa zea Resistance Levels to Cry1Ac and Cry2Ab2 across the Southeastern USA. Insects 2023, 14, 262. [Google Scholar] [CrossRef] [PubMed]
- USEPA (U.S. Environmental Protection Agency). Review of Data Submission to Support Continued Use of Natural Refuge for Managing Insect Resistance in Cotton. 2022. Available online: https://www.regulations.gov/document/EPA-HQ-OPP-2011-0922-0056 (accessed on 2 April 2023).


{kind=link}
| Hybrid (Bt Trait) | Year [No. of Trials] | % Ears Damaged | Number of Larvae per Ear | Kernel Consumption per Damaged Ear (cm2) | Proportion of Late Instars |
|---|---|---|---|---|---|
| Obsession I (Non-Bt isoline to Obsession II) | 2020 (41) | 78.9 ± 30.3 | 1.13 ± 0.68 | 6.5 ± 3.8 | 74.0 ± 24.2 |
| 2021 (42) | 70.2 ± 31.9 | 0.88 ± 0.63 | 6.3 ± 3.5 | 78.0 ± 22.1 | |
| 2022 (44) | 80.9 ± 30.5 | 1.20 ± 0.68 | 7.3 ± 4.0 | 79.1 ± 20.1 | |
| Obsession II (Cry1A.105 + Cry2Ab2) | 2020 (41) | 68.7 ± 29.6 # | 0.92 ± 0.68 # | 4.1 ± 2.8 # | 56.0 ± 28.1 # |
| 2021 (46) | 57.5 ± 36.8 # | 0.72 ± 0.61 # | 4.5 ± 3.3 # | 57.6 ± 32.1 # | |
| 2022 (45) | 68.5 ± 33.0 # | 0.97 ± 0.72 # | 4.7 ± 3.2 # | 59.1 ± 32.5 # | |
| Providence (Non-Bt isoline to BC0805 and Remedy) | 2020 (41) | 80.0 ± 27.7 | 1.18 ± 0.71 | 7.0 ± 2.7 | 82.1 ± 17.5 |
| 2021 (50) | 70.6 ± 33.6 | 0.95 ± 0.68 | 6.2 ± 3.4 | 77.1 ± 22.7 | |
| 2022 (55) | 80.1 ± 27.5 | 1.23 ± 0.70 | 7.3 ± 4.3 | 78.1 ± 18.3 | |
| BC0805 (Cry1Ab) | 2020 (41) | 77.1 ± 17.9 | 1.23 ± 1.02 | 5.4 ± 2.6 * | 66.0 ± 27.7 * |
| 2021 (41) | 66.7 ± 33.8 | 0.94 ± 0.70 | 5.6 ± 3.5 | 69.7 ± 24.9 | |
| 2022 (45) | 74.9 ± 30.8 | 1.26 ± 1.01 | 6.0 ± 3.7 | 67.4 ± 23.3 * | |
| Remedy (Cry1Ab + Vip3Aa) | 2020 (41) | 1.44 ± 4.09 | 0.004 ± 0.02 | 0.33 ± 0.88 | 0.0 ± 0.0 |
| 2021 (52) | 0.82 ± 2.18 | 0.006 ± 0.02 | 0.96 ± 2.05 | 10.1 ± 26.8 | |
| 2022 (53) | 0.37 ± 0.99 | 0.004 ± 0.02 | 0.49 ± 1.27 | 6.9 ± 24.8 |
| Hybrid (Bt Toxins Expressed) | Year | No. of Trials b | Phenotypic Frequency of Resistance | 95% Confidence Limits |
|---|---|---|---|---|
| BC0805 (Cry1Ab) | 2020 | 31 | 0.97 | 0.85–1.09 |
| 2021 | 28 | 0.98 | 0.84–1.07 | |
| 2022 | 33 | 1.02 | 0.95–1.09 | |
| Obsession II (Cry1A.105 + Cry2Ab2) | 2020 | 23 | 0.88 | 0.77–0.98 |
| 2021 | 23 | 0.93 | 0.78–1.08 | |
| 2022 | 31 | 0.82 | 0.75–0.90 | |
| Remedy (Cry1Ab + Vip3Aa) | 2020 | 36 | 0.002 | −0.0014–0.0048 |
| 2021 | 37 | 0.009 | 0.0022–0.0153 | |
| 2022 | 43 | 0.002 | 0.0003–0.0048 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Dively, G.P.; Kuhar, T.P.; Taylor, S.V.; Doughty, H.; Holmstrom, K.; Gilrein, D.O.; Nault, B.A.; Ingerson-Mahar, J.; Huseth, A.; Reisig, D.; et al. Extended Sentinel Monitoring of Helicoverpa zea Resistance to Cry and Vip3Aa Toxins in Bt Sweet Corn: Assessing Changes in Phenotypic and Allele Frequencies of Resistance. Insects 2023, 14, 577. https://doi.org/10.3390/insects14070577
Dively GP, Kuhar TP, Taylor SV, Doughty H, Holmstrom K, Gilrein DO, Nault BA, Ingerson-Mahar J, Huseth A, Reisig D, et al. Extended Sentinel Monitoring of Helicoverpa zea Resistance to Cry and Vip3Aa Toxins in Bt Sweet Corn: Assessing Changes in Phenotypic and Allele Frequencies of Resistance. Insects. 2023; 14(7):577. https://doi.org/10.3390/insects14070577
Chicago/Turabian StyleDively, Galen P., Tom P. Kuhar, Sally V. Taylor, Helene Doughty, Kristian Holmstrom, Daniel O. Gilrein, Brian A. Nault, Joseph Ingerson-Mahar, Anders Huseth, Dominic Reisig, and et al. 2023. "Extended Sentinel Monitoring of Helicoverpa zea Resistance to Cry and Vip3Aa Toxins in Bt Sweet Corn: Assessing Changes in Phenotypic and Allele Frequencies of Resistance" Insects 14, no. 7: 577. https://doi.org/10.3390/insects14070577