1. Introduction
Entomopathogenic fungi are important natural regulators of insect populations, and are widely used as environmentally friendly biological control alternatives to synthetic chemical insecticides [
1,
2,
3,
4]. In biological studies, measurements of fungal growth over time on different defined media serve as a standard measure to assess performance and evaluate efficacy during isolation and testing [
5,
6]. Being heterotrophic organisms with indeterminate growth, fungal growth often responds directly to the quality and availability of nutrients in the immediate environment [
7,
8]. Fungal growth over time can be evaluated using various methods, generally classified as direct or indirect measures [
9]. The most widely applied direct methods include measurement of hyphal extension [
10], dry weight from liquid cultures, and radial expansion on solid agar [
11,
12]. Accepted indirect methods make use of spectrophotometry in liquid macro- or micro-cultures [
10,
13,
14], fluorescence of labeled fungi and light sheet fluorescence [
15,
16], multispectral imaging [
17], quantification of chitin production [
18,
19], and advances in the spectrophotometric analysis of microcultures on agar media [
20].
Measurement of the radial expansion of fungal colonies on solid media is a widely-used direct method to quantify growth. The method is straightforward, and allows for measurement of other phenotypic traits such as spore production and colony morphology (e.g., color, branching pattern) [
6,
8,
21]. However, this approach fails to account for some important aspects of growth, such as the density of the mycelium [
20]. An expanding colony can exhibit varying degrees of mycelial density while covering the same area, which is not captured when using radial expansion measurements. Using a dry weight method to measure growth in liquid macrocultures accounts for this discrepancy in mycelial density, but requires interference with cultures through direct sampling for quantification, making real-time monitoring of in situ growth impractical [
13,
22].
In addition to this, the solid media environment may not be biologically appropriate for some fungi based on their specific ecologies. In entomopathogenic fungi, in vivo growth progresses primarily through the insect hemocoel, which is a submerged liquid environment [
23]. The physiological and developmental biology of fungi can vary considerably, depending on whether they grow on a solid or liquid medium. For instance, solid media may support a higher production of secondary metabolites or enzymes compared to liquid media in some fungal isolates [
24,
25]. In terms of practicality, fungal growth on agar plates is also time-consuming to conduct. The timescale for growth of many entomopathogenic fungi when analyzing radial expansion spans multiple days, and in the case of measuring dry weight, the processing of samples involves a lengthy process of collection, drying, and weighing [
3,
9,
21].
In situ spectrophotometry provides a viable alternative to these methods by directly correlating optical density (OD) values with an increase in fungal biomass. It is generally understood that growing fungus changes the OD of liquid cultures, because turbidity directly correlates with unit population size, which serves as the basis for the traditional spectrophotometric analysis of fungal growth [
26]. In this study, we apply spectrophotometric analysis to measure the growth of entomopathogenic fungi, which are typically performed on solid agar media, as previously described. Previous studies have demonstrated the use of spectrophotometric measurements for assessing filamentous fungal growth in microcultures. However, in these studies, the relationship between dry mycelial mass and OD is extrapolated using correlation coefficients [
13], or hyphal extension is employed as a growth metric [
10]. The aim of this study was to establish a direct correlation between the dry weight of mycelial cultures and their corresponding OD values for six isolates of
Metarhizium spp. This correlation allows for the construction of isolate-specific standard curves, enabling the quantification of biomass based on OD measurements.
2. Materials and Methods
2.1. Fungal Isolates and Preparation of Inoculum
The growth of two isolates within each of three different species of
Metarhizium were compared to investigate variations among species and isolates. This was assessed in addition to the effect on the relationship between dry weight and OD. Six fungal isolates of the genus
Metarhizium were used to produce standard curves of OD by dry weight:
M. brunneum KVL 16_36 (Isolated from the commercial product Met52, Novozymes A/S, Krogshøjvej 36, Bagsværd, Denmark),
M. brunneum KVL 12_30 [
27],
M. acridum KVL 18_06 (ARSEF 6421),
M. acridum KVL 04_55 (ARSEF 7486),
M. guizhouense KVL 19_24 (ARSEF 977), and
M guizhouense KVL 19_28 (ARSEF 3611). The acronym ARSEF refers to the United States Department of Agriculture (USDA) Agricultural Research Service (ARS) collection of Entomopathogenic Fungal cultures (
https://data.nal.usda.gov/dataset/ars-collection-entomopathogenic-fungal-cultures-arsef. URL accessed on 11 March 2022). The acronym KVL refers to the entomopathogenic fungus culture collection maintained at the Section for Organismal Biology, Department of Plant and Environmental Sciences, University of Copenhagen.
Metarhizium (Metschnikoff) Sorokin (Order Hypocreales: Family Clavicipitaceae) was selected as the focus of our study, due to its multifaceted importance in both evolutionary ecology and practical applications in pest management [
2,
3,
4].
The fungal cultures were grown on quarter-strength Sabouraud dextrose agar + yeast media (SDAY/4: 2.5 g/L 1:1 animal:bacterial peptone (bacteriological peptone and Acuferm Neoeptone, Neogen Corp., 620 Lesher Place, Lansing, MI, USA), 10 g/L dextrose (Bacteriological, Oxoid Ltd., Wade Road, Hampshire, UK), 2.5 g/L yeast extract (Neogen Corp.), 15 g/L agar (Bacteriological (European Type) No. 1, Neogen Corp.))) in Petri dishes (90 mm × 15 mm triple-vented, Sterilin Ltd., 1 Ashley Road, Altrincham, Cheshire, UK) at 23 °C, and conidia were harvested after 14 days. The conidia were collected in 0.1% (v/v) Tween® 80 (Merk KgaA, Frankfurter Straße 250, Darmstadt, Germany) via agitation with a Drigalski spatula from sporulating colonies, and the resulting suspension was centrifuged; the supernatant was removed, and the colonies were rinsed twice with 0.1% (v/v) Tween® 80 to remove all fragments of mycelia. Conidial suspensions were prepared at a concentration of 2 × 106 conidia per mL by dilution using 0.1% (v/v) Tween® 80. The concentration of the stock suspension was determined by counting conidia from 1000× serially diluted stock suspension in a Fuchs Rosenthall hemocytometer (×400 magnification). To verify germination, 100 µL of 100× diluted stock suspension was spread with a Drigalski spatula on an agar plate of SDAY/4 and incubated for 24 h at 23 °C. Four microscope coverslips (22 mm × 22 mm) were then placed over the culture surface, and 100 conidia were counted under each coverslip. Conidia were considered to have germinated with the germ tube that was at least as long as the width of the conidium, and conidial germination after 24 h was verified as >98% in all cases before being used in further assays.
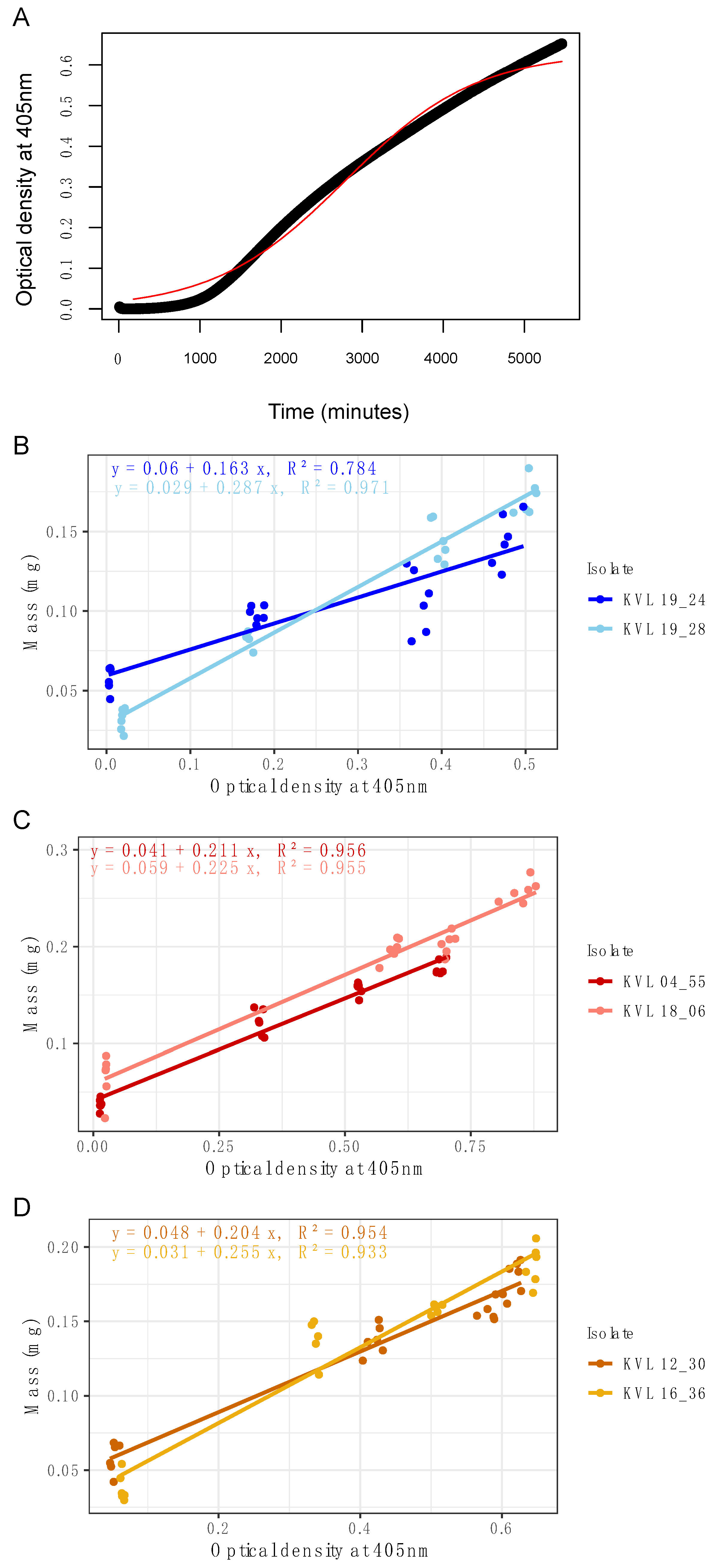
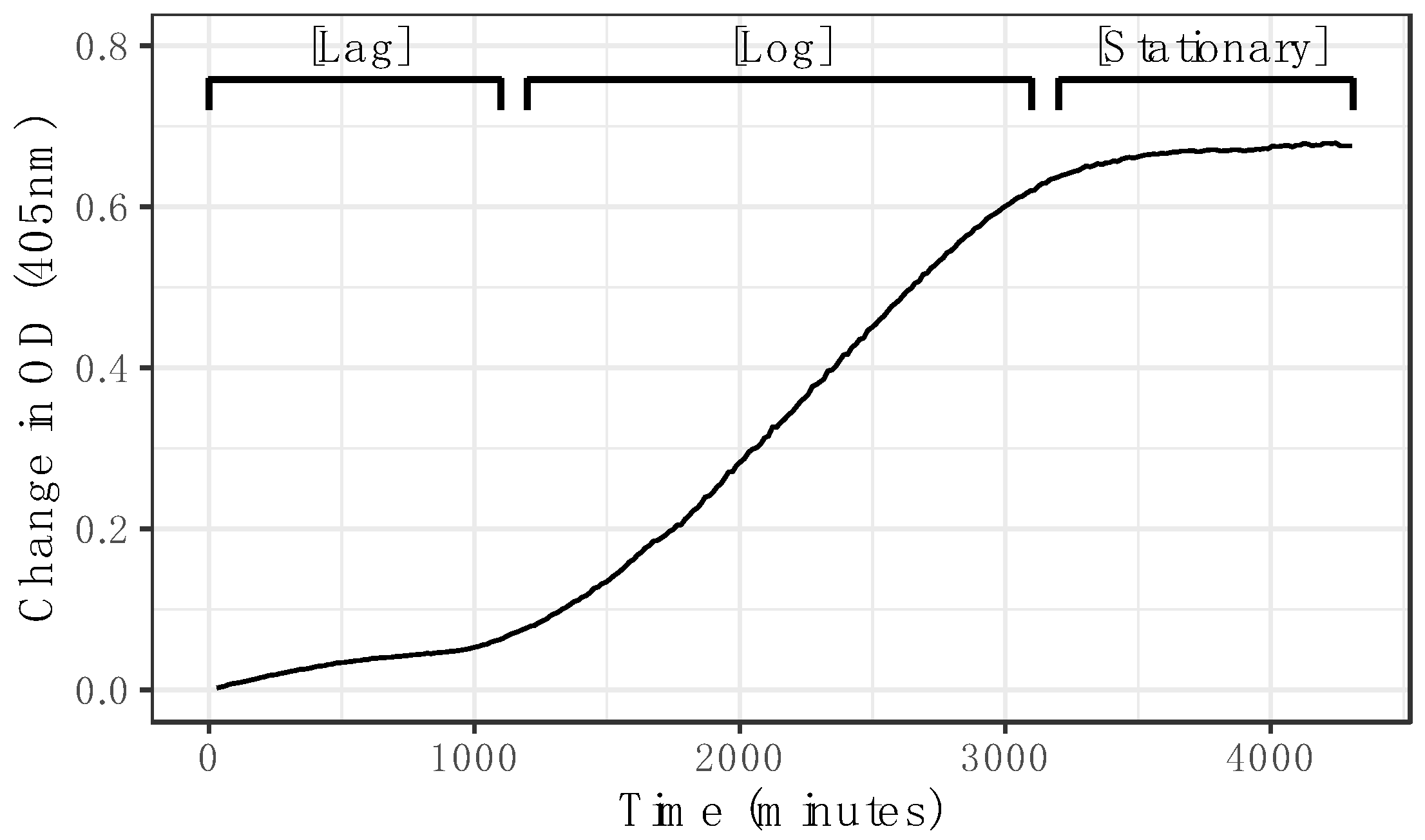
2.2. Continuous Growth Curve
To determine whether measurement by spectrophotometry in liquid microcultures can accurately capture the growth curve of an entomopathogenic fungus, a growth curve of
M. brunneum KVL 12_30 was produced. The microcultures were prepared in 96 wells of a clear flat-bottom vented microplate (Starlab International GmbH, Neuer Höltigbaum 38, Hamburg, Germany) by inoculating 100 µL of 2 × 10
6 conidial suspension into 100 µL Sabouraud dextrose + yeast media (SDY/4: 2.5 g/L 1:1 animal:bacterial peptone, 10 g/L dextrose, 2.5 g/L yeast extract). The OD of each well was measured at 405 nm in a Synergy
TM HT MultiDetection Microplate Reader (BioTek Instruments Ltd., Cheadle, UK) with Gen5 software Version 2.00.18 every 10 min for 96 h at 24 °C, without removal of the microplate from the plate reader. The wavelength of 405 nm was previously described as being fit for this purpose [
10]. Pilot assays were performed comparing a range of wavelengths for measuring growth in microcultures that confirmed this. The microcultures were checked visually for bacteria using a compound microscope upon completion to ensure they were uncontaminated.
2.3. Standard Curve for Fungal Dry Weight and OD
To establish a correlation between OD and fungal biomass, OD values were measured, and the mycelial mats were subsequently extracted and weighed for the six isolates at four time points (20, 40, 60, and 80 h post-inoculation) during the linear growth phase. The period of logistic growth was determined from the continuous growth curve generated, as detailed in
Section 2.2, utilizing the described parameters for media and conidial suspension. The fungal microcultures were prepared as described in
Section 2.2. For each measurement at the four time points, a single microplate containing 60 wells was utilized to assess the determination of dry weight, resulting in a total of 240 wells analyzed across the four measurements. To mitigate edge effects arising from temperature and evaporation, the 36 wells around the edge of the plate were excluded from analysis and loaded with 200 µL of blank media [
28]. The microplates were incubated at 23 ± 0.5 °C, and the OD measured at 405 nm at 20, 40, 60, and 80 h post-inoculation using a Synergy
TM HT MultiDetection microplate reader with Gen5 software. At each time point, one microplate per isolate was collected after OD measurement and stored at −20 °C. The OD values of all of the microplates were also measured after conidia settled (15 min post-inoculation) to establish the baseline reading OD of each culture. This baseline reading was subtracted from subsequent measurements to determine changes in OD for the construction of standard curves.
To quantify the changes in biomass of the microcultures, mycelial mats were extracted for the determination of dry weight. Thawed microplates were centrifuged in an Eppendorf Centrifuge 5810R (1968× g) at 4000 rpm for three minutes to force fungal material to the bottom of the wells, and the remaining media supernatant was removed using a pipette. The wells were then filled with 200 µL of 99% ethanol, and mycelia were scraped from the bottom of the well with a pipette tip to re-suspend the fungal material. The entire content of the well was then transferred to a pre-weighed aluminum weigh boat using a cut pipette tip. The process was repeated three times, refilling each well with 200 µL of ethanol and scraping to ensure complete removal of residual mycelial matter and rinsing of the pipette tip. Thus, a total of 600 µL of ethanol was utilized to thoroughly wash each well. Complete extraction of mycelial material from the wells was confirmed through microscopic examination of the microplates. If any residual mycelial material was observed, the extraction process was repeated until the wells were free of any remaining matter. For each isolate, ten replicate wells containing resultant mycelial suspensions were pooled into pre-weighed aluminum boats, resulting in a total of six pooled dry weight measurements per microplate with an approximate volume of 6 mL. The pooled samples were subsequently dried in an oven for 72 h at 60 °C within a heat-resistant, lidded box. Finally, the dried samples were weighed on a precision scale (Sartorius ME36S Ultra Micro Balance, 31 g × 0.001 mg (Sartorius UK Ltd., Epsom, UK).
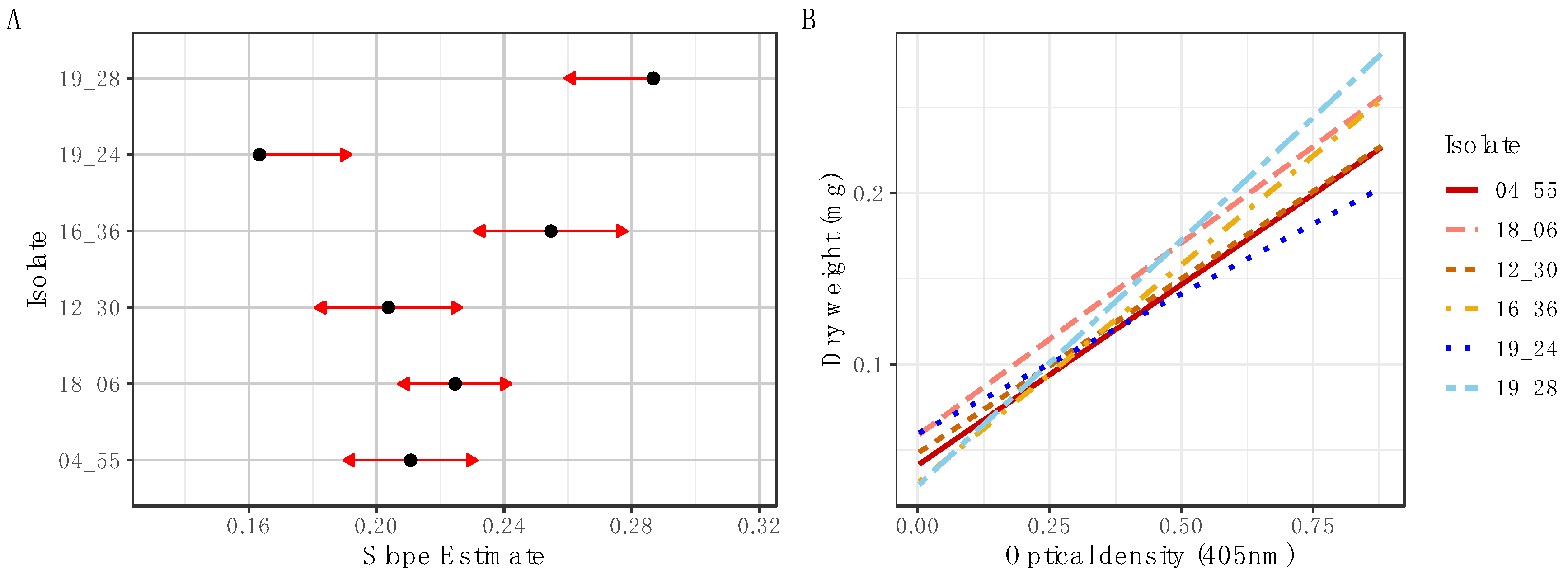
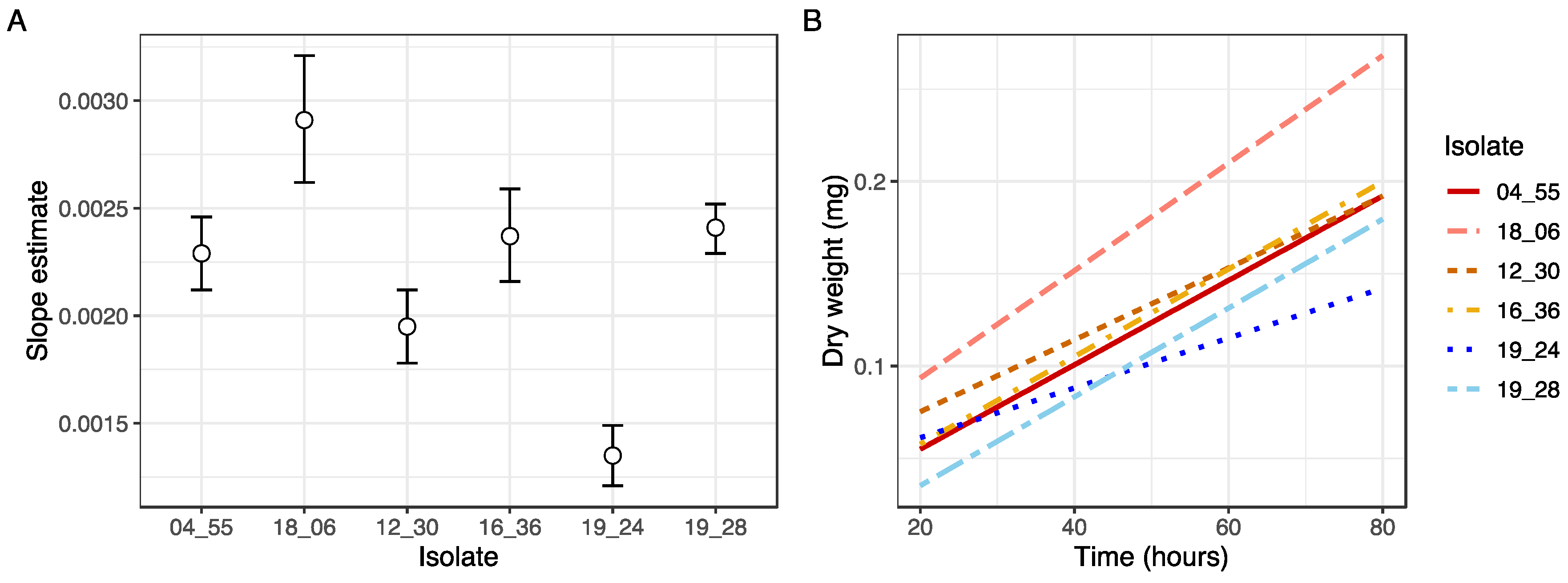
Standard curves were produced to establish the correlation between OD and change in biomass using the OD measurements and their corresponding pooled biomass samples. At each measurement time point, the base OD values were subtracted from the measured OD values, and the resulting OD values for the 10 pooled wells in the sample were averaged. Regression analysis was performed to construct standard curves of OD by dry weight using the R package stats [
29]. To test for differences in relationships between the OD and dry weight between isolates, a pairwise comparison of slope estimates was conducted using lsmeans [
30]. This analysis applies a
p value multiplicity adjustment to the least squares means of each isolate using Tukey’s HSD. Linear regression was also performed to determine slope estimates for biomass added over time for each isolate. All statistical calculations were conducted in R Version 1.4.1717 [
29].
2.4. Comparison of OD Growth Measure to Radial Growth Measure on Agar Plates
To evaluate how the microspectrophotometric method compared to traditional growth assays performed on solid media, the growth of two isolates (
M. guizhouense KVL 19_28 and
M. acridum KVL 04_55) was measured using both microspectrophotometry and radial expansion methods. These experiments were conducted under the same conditions at 23 °C in SDY/4 and SDAY/4 media, respectively. For the microspectrophotmetric analysis, media and conidial suspensions were prepared as described in
Section 2.2 in a microplate (n = 96). OD measurements were taken during the linear phase of the logistic growth curve (24 h post-inoculation) at 405 nm in 8-hour intervals. This provided a total of five measurements. To estimate the biomass, the linear equations deduced from the standard curves were applied to the spectrophotometric measurements. This allowed for the determination of the biomass estimation based on the spectrophotometric data.
To conduct the radial expansion analysis, for each isolate, four Petri dishes (90 mm × 15 mm, vented) of SDAY/4 were each inoculated with five cultures (n = 20). The inoculation was performed using 5 µL of a conidial suspension with a concentration of 2.4 × 10
4 spores per mL, prepared according to the methods described in
Section 2.1. Germination of the conidial suspension was confirmed as >99% after 24 h on a plate of SDAY/4, following the methods detailed in
Section 2.1. The growth was recorded every 4 days, starting from the first day of detectable mycelium formation (day 4) until day 11. To calculate the radial expansion, the plates were digitally photographed at each measurement time point, and the colony area was calculated in ImageJ Version 1.53s [
31].
4. Discussion
In this study, we demonstrate that microspectrophotometry can be used to capture the growth curve of entomopathogenic fungi in situ, and that the change in absorbance can be directly correlated with an increase in fungal biomass. Optical density (OD) values correlated strongly with biomass, and it was demonstrated that it is possible to produce a standard curve for the quantification of dry weight from OD by extracting, drying, and weighing the microcultures of mycelia on a precision scale (
Figure 1).
In addition, we showed that change in the OD accurately represents biomass accumulation. This is not always the case in measurements of radial expansion, as the expansion of fungal colonies on solid media can often be equal in area, while having significantly different densities of mycelial growth [
20]. Fungal cultures grow in multiple planar dimensions on agar (i.e., upwards and downwards, as well as across the substrate surface), and the hyphal mass can thus be more or less dense depending on nutrient availability [
32]. The microspectrophotometric technique provides a quantitative measurement of hyphal density that is more uniformly distributed throughout the microculture, which produces a better representation of three-dimensional hyphal density than measuring two dimensions. This technique also makes it possible to capture specific effects from environmental changes on different fungal growth phase features that are only detectable at small time scales (
Figure 4B), which produce atypical growth curves, e.g., rapid depletion of a primary nutrient source followed by a secondary phase of logarithmic growth [
33]. Furthermore, the solid medium does not accurately represent the environment which entomopathogenic fungi encounter during infection (i.e., insect hemolymph). Given that most entomopathogenic fungi are filamentous with similar ecologies (i.e., penetration of host cuticle and subsequently spreading as individual cells through the insect hemocoel), this technique could be applied to investigate growth in other entomopathogenic species [
34]. However, differences in biomass accumulation during growth as represented in the relationship between fungal dry weight and OD necessitates the production of standard curves specific to the isolate under investigation, although this would only need to be undertaken once per isolate to be applied to subsequent high throughput measurements.
In this study, we compared the use of microspectrophotometry with radial expansion analysis to assess the growth dynamics of two species of
Metarhizium, M. acridum and
M. guizhouense. Our findings demonstrated that the microspectrophotometric analysis yielded similar end point results and linear growth rates compared to the radial growth assay (
Figure 4A–C). However, the microspectrophotometric technique provided the additional advantage of capturing differences in early growth rates that were not evident in the radial growth assay (
Figure 4). The finer scale of measurement provided by the microspectrophotometric technique illustrates its value in accurately capturing different aspects of fungal growth curves that might be otherwise undetectable when measuring macroculture growth over many days. Compared to methods with fewer measurement points, this technique is better suited for detecting subtle differences in growth (
Figure 4).
Importantly, our study revealed a different relationship between
M. acridum and
M. guizhouense through finer measurement intervals during critical growth phases, particularly the early linear phase, which could not be discerned through radial expansion measurements performed over several days (
Figure 4). This development is significant for understanding the growth dynamics of fungi, as it has been previously shown that the classical growth curve does not always adequately describe the growth patterns of filamentous fungi [
33]. Different aspects of the growth curve can change due to variables such as nutrition and the host insect environment. Capturing the growth pattern can offer important insights into various aspects of how fungi grow during infection. Notably, atypically shaped growth curves have been suggested to be the rule rather than the exception [
33]. This arises from the depletion of distinct nutrients occurring at different rates, resulting in atypical-shaped growth curves, like bimodal growth peaks [
33]. These curves reveal nutritive preferences that might be otherwise overlooked without the necessary sensitivity in measurements. Our study revealed that the
M. acridum KVL 04_55 isolate generally exhibited more growth over time compared to other isolates (
Figure 4). However, a notable finding was that in the early stages, this isolate displayed a significantly slower growth rate compared to the other isolate examined,
M. guizhouense KVL 19_28. This observation provides important insights into the growth dynamics and pathogenic potential during early infection processes for this particular isolate [
35].
Additionally, our findings have practical implications regarding the pathogenic potential of the different isolates and species measured. By comparing the growth rates and total biomass produced between isolates, our findings shed light on the potential speed of host invasion across different species of
Metarhizium. Significantly differing slope estimates and total biomass production were observed not only between species, but also among isolates, indicating variations in pathogenicity potential within this in vitro setting (
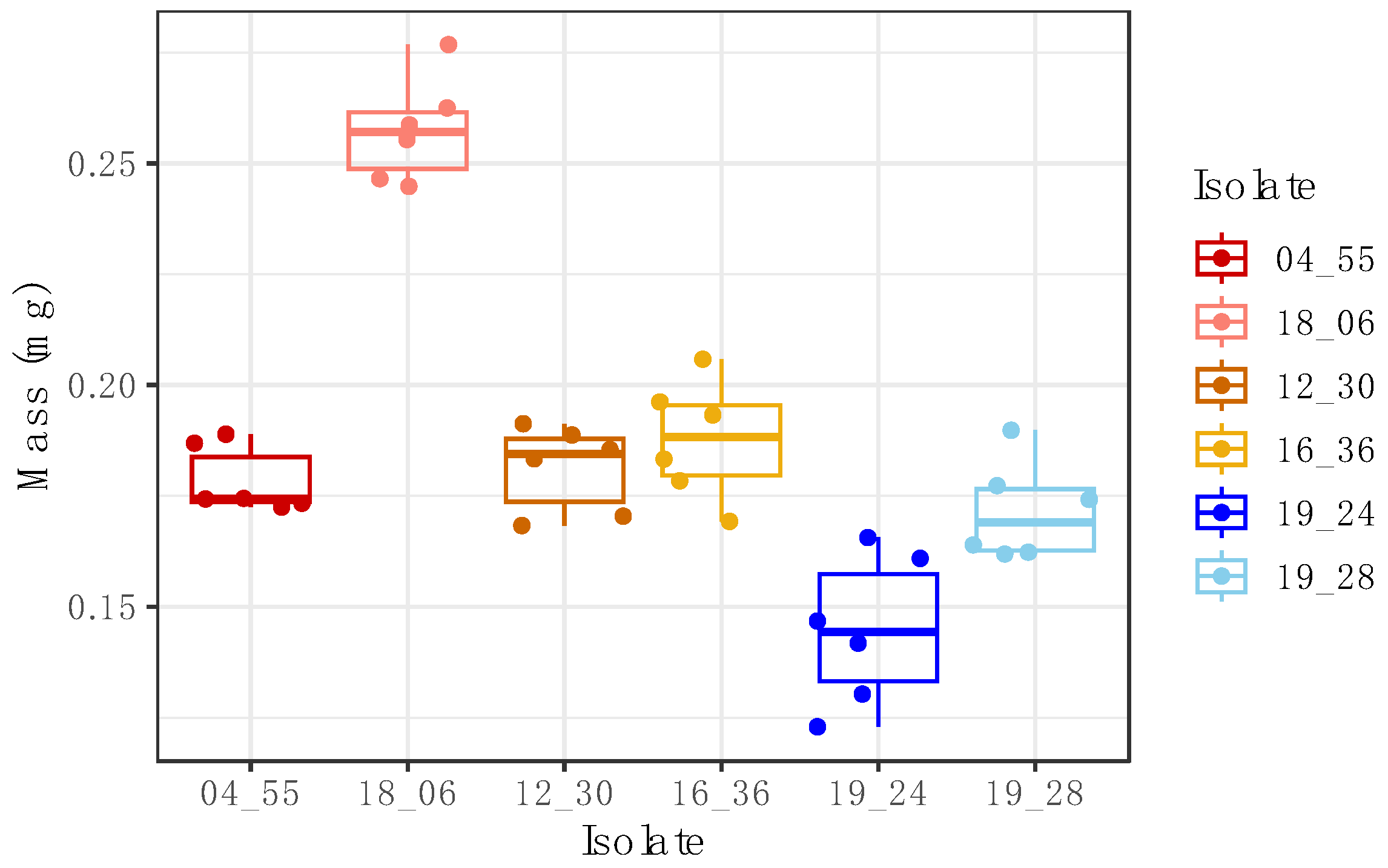
Figure 3 and
Figure A2). This highlights the practical application of the method in assessing the virulence and pathogenic potential of the examined isolates and species.
Previous studies demonstrated the use of microspectrophotometry in monitoring the growth of filamentous fungi, but were unable to determine the dry weight of individual microcultures, and therefore relied on comparisons of indirect metrics to extrapolate the relationship between OD and fungal dry weight [
10,
13], or used microscopic measurements of hyphal extension to infer growth [
10]. In this research, we inferred growth in microculture using mycelial dry weight, and generated standard curves for direct correlation. While the microspectrophotometric technique may provide limited phenotypic information compared to solid media bioassays, such as the measurement of spore production and colony color, it offers complementary advantages in capturing biomass build-up during the early phases of the growth curve, and obtains data rapidly. The speed at which growth data can be obtained using microspectrophotometry is a clear advantage, as linear growth of the tested
Metarhizium isolates could be measured within a few days, whereas radial growth on agar plates typically takes 10–14 days.
The need to produce entomopathogenic fungi for biocontrol at massive scales makes it important to be able to investigate the effects of different media and nutrients on entomopathogenic fungal growth, and identify optimal growth parameters [
36]. In addition, for experimental biologists, this method allows for large-scale experiments using growth as a primary measure of performance in areas of research such as fitness costs, adaptation [
37], and niche quantification [
38]. A more detailed picture of the different growth phases could provide an understanding of nutritional adaptation, for example, in revealing nutritive preferences [
33].
In conclusion, this technique allows for the rapid generation of growth curves of entomopathogenic fungi at a fine timescale with many replicates, and in a medium that is more ecologically relevant to entomopathogenic fungi than what typical solid media bioassays provide. Furthermore, this approach has the potential for application to other species of filamentous entomopathogenic fungi under investigation, such as
Beauveria spp.,
Hirsutella spp.,
Cordyceps spp., and
Lecanicillium spp. [
3]. The methodological developments described advance the applications of spectrophotometry to the monitoring of filamentous fungal growth in entomopathogenic fungi, and resolve the infeasibility of producing standard curves directly correlating change in OD with mycelial mass. This automated and high-throughput method for monitoring in situ fungal growth presented here will aid further studies on aspects affecting growth of these ecologically and commercially important organisms.









{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}