
Molecular Characterization of Serratia marcescens Strain Isolated from Yellow Mealworms, Tenebrio molitor, in The Netherlands
{kind=link}
{kind=link}
Abstract
:Simple Summary
Abstract
1. Introduction
2. Materials and Methods
3. Results
3.1. Characteristics of the T. molitor Specimen
3.1.1. Bacterial Identification
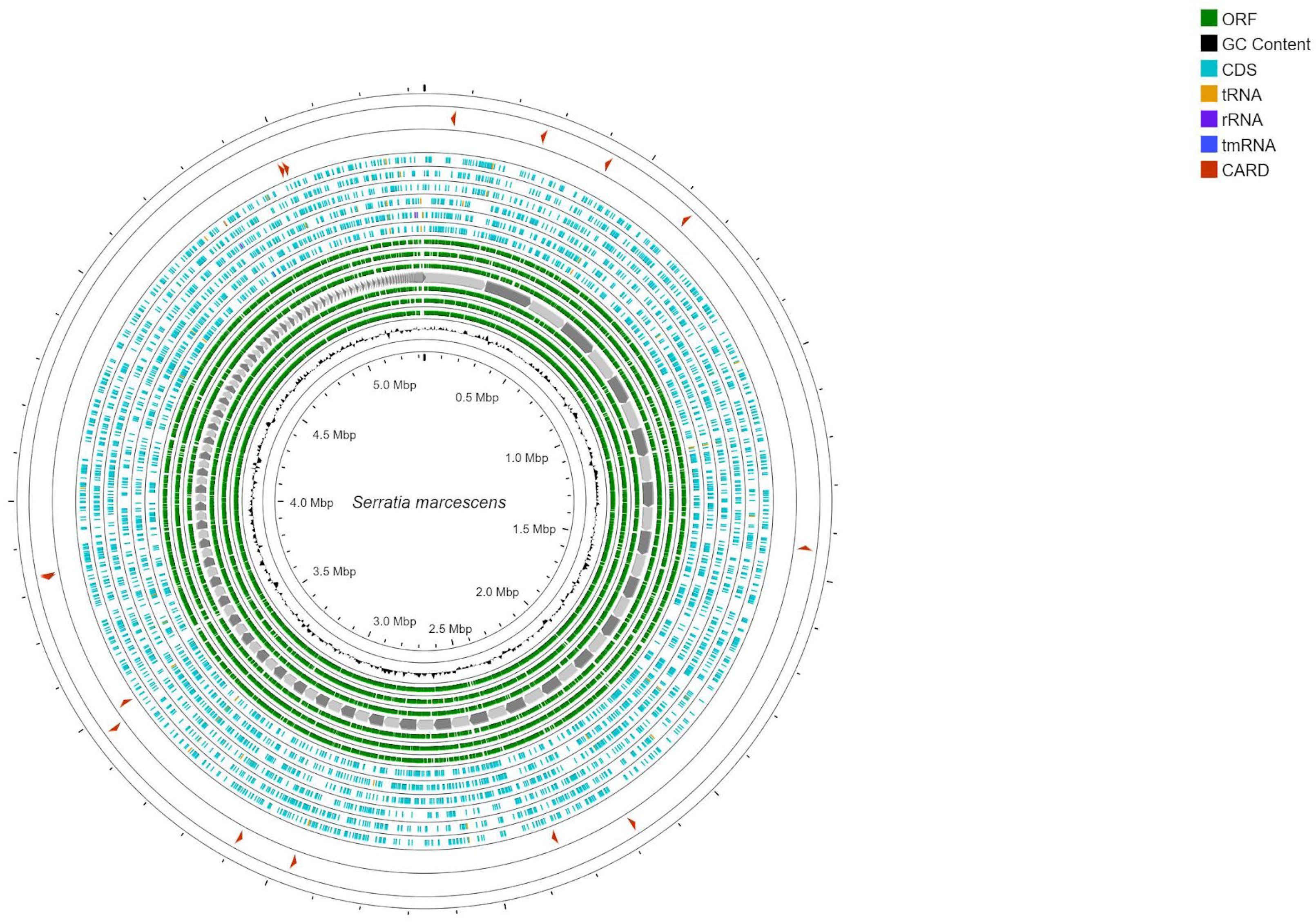
3.1.2. Molecular Characterization of S. marcescens
3.1.3. Phylogenetic Tree
4. Discussion
5. Conclusions
Author Contributions
Funding
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Van Huis, A. Insects as food and feed, a new emerging agricultural sector: A Review. J. Insects Food Feed. 2020, 6, 27–44. [Google Scholar] [CrossRef]
- Morales-Ramos, J.A.; Kay, S.; Rojas, M.G.; Shapiro-Ilan, D.I.; Tedders, W.L. Morphometric analysis of instar variation in Tenebrio molitor (Coleoptera: Tenebrionidae). Ann. Entomol. Soc. Am. 2015, 108, 146–159. [Google Scholar] [CrossRef]
- Ludwig, D.; Fiore, C. Further studies on the relationship between parental age and the life cycle of the mealworm, Tenebrio molitor. Ann. Entomol. Soc. Am. 1960, 53, 595–600. [Google Scholar] [CrossRef]
- Veldkamp, T.; Claeys, J.; Haenen, O.L.M.; van Loon, J.J.A.; Spranghers, T. The Basics of Edible Insect Rearing; Wageningen Academic Publishers: Wageningen, The Netherlands, 2021; ISBN 978-90-8686-365-5. [Google Scholar]
- Bjørge, J.D.; Overgaard, J.; Malte, H.; Gianotten, N.; Heckmann, L.-H. Role of temperature on growth and metabolic rate in the tenebrionid beetles Alphitobius diaperinus and Tenebrio molitor. J. Insect Physiol. 2018, 107, 89–96. [Google Scholar] [CrossRef] [PubMed]
- Punzo, F.; Mutchmor, J.A. Effect of temperature, relative humidity and period of exposure on the survival capacity of Tenebrio molitor (Coleoptera: Tenebrionidae). J. Kans. Entomol. Soc. 1980, 53, 260–270. [Google Scholar]
- EFSA Scientific Committee. Risk profile related to production and consumption of insects as food and feed. EFSA J. 2015, 13, 4257. [Google Scholar] [CrossRef]
- Osimani, A.; Milanović, V.; Cardinali, F.; Garofalo, C.; Clementi, F.; Pasquini, M.; Riolo, P.; Ruschioni, S.; Isidoro, N.; Loreto, N.; et al. The bacterial biota of laboratory-reared edible mealworms (Tenebrio molitor L.): From feed to frass. Int. J. Food Microbiol. 2018, 272, 49–60. [Google Scholar] [CrossRef]
- Jensen, A.N.; Hansen, S.H.; Baggesen, D.L. Salmonella typhimurium level in mealworms (Tenebrio molitor) after exposure to contaminated substrate. Front. Microbiol. 2020, 11, 1613. [Google Scholar] [CrossRef]
- Council on Animal Affairs (RDA). The Emerging Insect Industry; Invertebrates as Production Animals; RDA: The Hague, The Netherlands, 2018. [Google Scholar]
- Pinotti, L.; Ottoboni, M. Substrate as insect feed for bio-mass production. J. Insects Food Feed. 2021, 7, 585–596. [Google Scholar] [CrossRef]
- Eilenberg, J.; Vlak, J.M.; Nielsen-LeRoux, C.; Cappellozza, S.; Jensen, A.B. Diseases in insects produced for food and feed. J. Insects Food Feed. 2015, 1, 87–102. [Google Scholar] [CrossRef]
- Mahlen, S.D. Serratia infections: From military experiments to current practice. Clin. Microbiol. Rev. 2011, 24, 755–791. [Google Scholar] [CrossRef] [PubMed]
- Prashant, D. Serratia marcescens—An Overview. Microbes Notes. Available online: https://microbenotes.com/serratia-marcescens-an-overview/ (accessed on 13 May 2023).
- Ochieng, J.B.; Boisen, N.; Lindsay, B.; Santiago, A.; Ouma, C.; Ombok, M.; Fields, B.; Stine, O.C.; Nataro, J.P. Serratia marcescens is injurious to intestinal epithelial cells. Gut Microbes 2014, 5, 729–736. [Google Scholar] [CrossRef] [PubMed]
- Slain, D.; Howard, C.; Cooper, C.G. An antimicrobial treatment assessment of Serratia marcescens bacteremia and endocarditis. Front. Antibiot. 2022, 1, 942721. [Google Scholar] [CrossRef]
- Hejazi, A.; Falkiner, F.R. Serratia marcescens. J. Med. Microbiol. 1997, 46, 903–912. [Google Scholar] [CrossRef] [PubMed]
- Fusco, V.; Abriouel, H.; Benomar, N.; Kabisch, J.; Chieffi, D.; Cho, G.-S.; Franz, C.M.A.P. Opportunistic food-borne pathogens. In Food Safety and Preservation; Elsevier: Amsterdam, The Netherlands, 2018; pp. 269–306. [Google Scholar]
- Public Health Agengy of Canada. Pathogen Safety Data Sheets: Infectious Substances—Serratia spp.; Public Health Agency of Canada: Ottawa, ON, Canada, 2011. Available online: https://www.canada.ca/en/public-health/services/laboratory-biosafety-biosecurity/pathogen-safety-data-sheets-risk-assessment/serratia.html (accessed on 13 May 2023).
- Wang, J.; Ishfaq, M.; Fan, Q.; Chen, C.; Li, J. A Respiratory commensal bacterium acts as a risk factor for Mycoplasma gallisepticum infection in chickens. Vet. Immunol. Immunopathol. 2020, 230, 110127. [Google Scholar] [CrossRef]
- Adebowale, O.; Adeyemo, O. Characterization of bacterium types isolated from commercial laying hen farms in Ogun state Nigeria. Rev. Elev. Med. Vet. Pays Trop. 2018, 71, 137–141. [Google Scholar] [CrossRef]
- El-Barbary, M.I.; Hal, A.M. Molecular identification and pathogenicity of citrobacter and serratia species isolated from cultured Oreochromis niloticus. Egypt. J. Aquat. Res. 2017, 43, 255–263. [Google Scholar] [CrossRef]
- Camus, A.; Berliner, A.; Clauss, T.; Hatcher, N.; Marancik, D. Serratia marcescens associated ampullary system infection and septicaemia in a bonnethead shark, Sphyrna tiburo (L.). J. Fish. Dis. 2013, 36, 891–895. [Google Scholar] [CrossRef]
- Patterson, K.L.; Porter, J.W.; Ritchie, K.B.; Polson, S.W.; Mueller, E.; Peters, E.C.; Santavy, D.L.; Smith, G.W. The etiology of white pox, a lethal disease of the Caribbean elkhorn coral, Acropora palmata. Proc. Natl. Acad. Sci. USA 2002, 99, 8725–8730. [Google Scholar] [CrossRef]
- Chen, S.; Blom, J.; Walker, E.D. Genomic, physiologic, and symbiotic characterization of Serratia marcescens strains isolated from the mosquito Anopheles stephensi. Front. Microbiol. 2017, 8, 1483. [Google Scholar] [CrossRef]
- Maciel-Vergara, G.; Jensen, A.; Eilenberg, J. Cannibalism as a possible entry route for opportunistic pathogenic bacteria to insect hosts, exemplified by Pseudomonas aeruginosa, a pathogen of the giant mealworm Zophobas morio. Insects 2018, 9, 88. [Google Scholar] [CrossRef] [PubMed]
- Maciel-Vergara, G.; Jensen, A.B.; Lecocq, A.; Eilenberg, J. Diseases in edible insect rearing systems. J. Insects Food Feed. 2021, 7, 621–638. [Google Scholar] [CrossRef]
- Prinsloo, H.E. The phospholipase and gelatinase activity of gram-negative bacteria isolated from diseased insects, and typing of strains of Serratia marcescens. J. Invertebr. Pathol. 1967, 9, 420–427. [Google Scholar] [CrossRef]
- Pineda-Castellanos, M.; Rodríguez-Segura, Z.; Villalobos, F.; Hernández, L.; Lina, L.; Nuñez-Valdez, M. Pathogenicity of isolates of Serratia marcescens towards larvae of the scarab Phyllophaga blanchardi (Coleoptera). Pathogens 2015, 4, 210–228. [Google Scholar] [CrossRef]
- Steinhaus, E.A. Serratia marcescens Bizio as an insect pathogen. Hilgardia 1959, 28, 351–380. [Google Scholar] [CrossRef]
- Slatten, B.H.; Larson, A.D. Mechanism of pathogenicity of Serratia marcescens. J. Invertebr. Pathol. 1967, 9, 78–81. [Google Scholar] [CrossRef]
- Glover, J.P.; Medrano, E.G.; Isakeit, T.; Brewer, M.J. Transmission of cotton seed and boll rotting bacteria by the verde plant bug (Hemiptera: Miridae). J. Econ. Entomol. 2020, 113, 793–799. [Google Scholar] [CrossRef]
- Kwak, K.-W.; Han, M.-S.; Nam, S.-H.; Choi, J.-Y.; Lee, S.-H.; Choi, Y.-C.; Park, K.-H. Detection of insect pathogen Serratia marcescens in protaetia Brevitarsis seulensis (Kolbe) from Korea. Int. J. Indust Entomol. 2014, 28, 25–31. [Google Scholar] [CrossRef]
- Vasantharajan, V.; Munirathnamma, N. Studies on silworm diseases. III. Epizootiology of a septicemic disease of silkworms caused by Serratia marcescens. J. Indian Inst. Sci. 2013, 60, 33. [Google Scholar]
- Inglis, G.D.; Lawrence, A.M. Effects of Serratia marcescens on the F1 generation of laboratory-reared Heliothis virescens (Lepidoptera: Noctuidae). J. Econ. Entomol. 2001, 94, 362–366. [Google Scholar] [CrossRef]
- Inglis, G.D.; Lawrence, A.M.; Davis, F.M. Pathogens associated with southwestern corn borers and southern corn stalk borers (Lepidoptera: Crambidae). J. Econ. Entomol. 2000, 93, 1619–1626. [Google Scholar] [CrossRef] [PubMed]
- Sikorowski, P.P.; Lawrence, A.M.; Inglis, G.D. Effects of Serratia marcescens on rearing of the tobacco budworm (Lepidoptera: Noctuidae). Am. Entomol. 2001, 47, 51–60. [Google Scholar] [CrossRef]
- Lighthart, B.; Sewall, D.; Thomas, D.R. Effect of several stress factors on the susceptibility of the predatory mite, Metaseiulus occidentalis (Acari: Phytoseiidae), to the Weak Bacterial Pathogen Serratia Marcescens. J. Invertebr. Pathol. 1988, 52, 33–42. [Google Scholar] [CrossRef]
- Burritt, N.L.; Foss, N.J.; Neeno-Eckwall, E.C.; Church, J.O.; Hilger, A.M.; Hildebrand, J.A.; Warshauer, D.M.; Perna, N.T.; Burritt, J.B. Sepsis and hemocyte loss in honey bees (Apis mellifera) infected with Serratia Marcescens strain Sicaria. PLoS ONE 2016, 11, e0167752. [Google Scholar] [CrossRef] [PubMed]
- Konecka, E.; Mokracka, J.; Krzyminska, S.; Kaznowski, A. Evaluation of the pathogenic potential of insecticidal Serratia marcescens strains to humans. Pol. J. Microbiol. 2019, 68, 185–191. [Google Scholar] [CrossRef]
- Dupriez, F.; Rejasse, A.; Rios, A.; Lefebvre, T.; Nielsen-LeRoux, C. Impact and persistence of Serratia marcescens in Tenebrio molitor larvae and feed under optimal and stressed mass rearing conditions. Insects 2022, 13, 458. [Google Scholar] [CrossRef]
- University of Copenhagen; Insect Doctors. Diseases and Disorders in Mealworm Tenebrio molitor. Available online: https://www.insectdoctors.eu/en/insectdoctors/diseases-disorders.htm (accessed on 15 June 2023).
- Da Silva, F.K.P.; Brück, D.W.; Brück, W.M. Isolation of proteolytic bacteria from mealworm (Tenebrio molitor) exoskeletons to produce chitinous material. FEMS Microbiol. Lett. 2017, 364, fnx177. [Google Scholar] [CrossRef]
- Mahata, M.E.; Lestari, I.; Dharma, A.; Ryanto, H.I.A.; Rizal, Y. Use of Serratia marcescens for feed processing: Broiler performance and pathogenicity assay. Int. J. Poult. Sci. 2012, 11, 424–426. [Google Scholar] [CrossRef]
- Baráti-Deák, B.; Da Costa Arruda, G.C.; Perjéssy, J.; Klupács, A.; Zalán, Z.; Mohácsi-Farkas, C.; Belák, Á. Inhibition of foodborne pathogenic bacteria by excreted metabolites of Serratia marcescens strains isolated from a dairy-producing environment. Microorganisms 2023, 11, 403. [Google Scholar] [CrossRef]
- Jansson, E.; Haenen, O.; Nonnemann, B.; Madsen, L.; Gelderen, E.V.; Aspan, A.; Säker, E.; Boonstra, M.; Gulla, S.; Colquhoun, D.J.; et al. MALDI-TOF MS: A diagnostic tool for identification of bacterial fish pathogens. Bull. Eur. Assoc. Fish. Pathol. 2020, 40, 240–248. [Google Scholar]
- Bushnell, B.; Rood, J.; Singer, E. BBMerge—Accurate paired shotgun read merging via overlap. PLoS ONE 2017, 12, e0185056. [Google Scholar] [CrossRef] [PubMed]
- Kumar, S.; Stecher, G.; Tamura, K. MEGA7: Molecular evolutionary genetics analysis version 7.0 for bigger datasets. Mol. Biol. Evol. 2016, 33, 1870–1874. [Google Scholar] [CrossRef] [PubMed]
- Saitou, N.; Nei, M. The neighbor-joining method: A new method for reconstructing phylogenetic trees. Mol. Biol. Evol. 1987, 33, 406–425. [Google Scholar] [CrossRef]
- Nei, M.; Kumar, S. Molecular Evolution and Phylogenetics; Oxford University Press: New York, NY, USA, 2000. [Google Scholar]
- Wang, H.; Li, Y.; Yang, M.; Zhang, X. Effects of temperature on weight gain, food conversion efficiency and digestive enzyme activities of Tenebrio molitor larvae. Chin. J. Appl. Entomol. 2017, 54, 434–439. [Google Scholar]
- Urs, K.C.D.; Hopkins, T.L. Effect of moisture on growth rate and development of two strains of Tenebrio molitor L. (Coleoptera, Tenebrionidae). J. Stored Prod. Res. 1973, 8, 291–297. [Google Scholar] [CrossRef]
- Inglis, G.D.; Lawrence, A.M.; Sikorowski, P.P. The use of meconia to nondestructively detect sublethal infections in heliothines (Lepidoptera: Noctuidae). J. Econ. Entomol. 2003, 96, 272–279. [Google Scholar] [CrossRef]
- Van der Fels-Klerx, H.J.; Camenzuli, L.; Belluco, S.; Meijer, N.; Ricci, A. Food safety issues related to uses of insects for feeds and foods. Compr. Rev. Food Sci. Food Saf. 2018, 17, 1172–1183. [Google Scholar] [CrossRef]
- Hoek van den Hil, E.F.; Antonis, A.F.G.; Brouwer, M.S.M.; Bruins, M.E.; Dame, M.A.; van Groenestijn, J.W.; Haenen, O.L.M.; Hoffmans, Y.; Meijer, N.P.; Veldkamp, T.; et al. Use of Insects for Food and Feed: Scientific Overview of the Present Knowledge on Insect Rearing, Use of Residual Streams as Substrates, and Safety Aspects; Report/Wageningen Food Safety Research; No. WFSR 2022.013; Wageningen University & Research: Wageningen, The Netherlands. [CrossRef]
- Piccirilli, A.; Cherubini, S.; Brisdelli, F.; Fazii, P.; Stanziale, A.; Di Valerio, S.; Chiavaroli, V.; Principe, L.; Perilli, M. Molecular Characterization by whole-genome sequencing of clinical and environmental Serratia marcescens strains isolated during an outbreak in a neonatal intensive care unit (NICU). Diagnostics 2022, 12, 2180. [Google Scholar] [CrossRef]
- Bolourchi, N.; Noori Goodarzi, N.; Giske, C.G.; Nematzadeh, S.; Haririzadeh Jouriani, F.; Solgi, H.; Badmasti, F. Comprehensive pan-genomic, resistome and virulome analysis of clinical OXA-48 producing carbapenem-resistant Serratia marcescens strains. Gene 2022, 822, 146355. [Google Scholar] [CrossRef]
- Thompson, S.A.; Maani, E.V.; Lindell, A.H.; King, C.J.; McArthur, J.V. Novel tetracycline resistance determinant isolated from an environmental strain of Serratia marcescens. Appl. Environ. Microbiol. 2007, 73, 2199–2206. [Google Scholar] [CrossRef]
- Iguchi, A.; Nagaya, Y.; Pradel, E.; Ooka, T.; Ogura, Y.; Katsura, K.; Kurokawa, K.; Oshima, K.; Hattori, M.; Parkhill, J.; et al. Genome evolution and plasticity of Serratia marcescens, an important multidrug-resistant nosocomial pathogen. Genome Biol. Evol. 2014, 6, 2096–2110. [Google Scholar] [CrossRef] [PubMed]
- Josenhans, C.; Suerbaum, S. The role of motility as a virulence factor in bacteria. Int. J. Med. Microbiol. 2002, 291, 605–614. [Google Scholar] [CrossRef] [PubMed]
- Greco-Stewart, V.S.; Brown, E.E.; Parr, C.; Kalab, M.; Jacobs, M.R.; Yomtovian, R.A.; Ramírez-Arcos, S.M. Serratia marcescens strains implicated in adverse transfusion reactions form biofilms in platelet concentrates and demonstrate reduced detection by automated culture. Vox Sang. 2012, 102, 212–220. [Google Scholar] [CrossRef] [PubMed]
- Chan, X.; Chang, C.; Hong, K.; Tee, K.; Yin, W.; Chan, K. Insights of biosurfactant producing Serratia marcescens strain w2.3 isolated from diseased tilapia fish: A draft genome analysis. Gut Pathog. 2013, 5, 29. [Google Scholar] [CrossRef]
- Vandeweyer, D.; Lenaerts, S.; Callens, A.; Van Campenhout, L. Effect of blanching followed by refrigerated storage or industrial microwave drying on the microbial load of yellow mealworm larvae (Tenebrio molitor). Food Control 2017, 71, 311–314. [Google Scholar] [CrossRef]
- Lenaerts, S.; Van Der Borght, M.; Callens, A.; Van Campenhout, L. Suitability of microwave drying for mealworms (Tenebrio molitor) as alternative to freeze drying: Impact on nutritional quality and colour. Food Chem. 2018, 254, 129–136. [Google Scholar] [CrossRef]
- Kröncke, N.; Grebenteuch, S.; Keil, C.; Demtröder, S.; Kroh, L.; Thünemann, A.; Benning, R.; Haase, H. Effect of different drying methods on nutrient quality of the yellow mealworm (Tenebrio molitor L.). Insects 2019, 10, 84. [Google Scholar] [CrossRef]
- Kröncke, N.; Böschen, V.; Woyzichovski, J.; Demtröder, S.; Benning, R. Comparison of suitable drying processes for mealworms (Tenebrio molitor). Innov. Food Sci. Emerg. Technol. 2018, 50, 20–25. [Google Scholar] [CrossRef]
- Kaira, G.S.; Dhakar, K.; Pandey, A. A psychrotolerant strain of Serratia marcescens (mtcc 4822) produces laccase at wide temperature and pH range. AMB Express 2015, 5, 1. [Google Scholar] [CrossRef]
- Prescott, L.M.; Harley, J.P.; Klein, D.A. Microbial growth—The influence of environmental factors on growth. In Microbiology, 5th ed.; McGraw-Hill: New York City, NY, USA, 2002. [Google Scholar]
- IPIFF Guide on Good Hygiene Practices. Available online: https://ipiff.org/good-hygiene-practices/ (accessed on 13 June 2023).
- Haenen, O.L.M.; Evans, J.J.; Berthe, F. Bacterial infections from aquatic species: Potential for and prevention of contact zoonoses. Rev. Sci. Tech. L’oie 2013, 32, 497–507. [Google Scholar] [CrossRef]


Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Bello Gonzalez, T.d.J.; van Gelderen, B.; Harders, F.; Vloet, R.; Voorbergen-Laarman, M.; de Ruiter, B.; Haenen, O.L.M. Molecular Characterization of Serratia marcescens Strain Isolated from Yellow Mealworms, Tenebrio molitor, in The Netherlands. Insects 2023, 14, 770. https://doi.org/10.3390/insects14090770
Bello Gonzalez TdJ, van Gelderen B, Harders F, Vloet R, Voorbergen-Laarman M, de Ruiter B, Haenen OLM. Molecular Characterization of Serratia marcescens Strain Isolated from Yellow Mealworms, Tenebrio molitor, in The Netherlands. Insects. 2023; 14(9):770. https://doi.org/10.3390/insects14090770
Chicago/Turabian StyleBello Gonzalez, Teresita d. J., Betty van Gelderen, Frank Harders, Rianka Vloet, Michal Voorbergen-Laarman, Bart de Ruiter, and Olga L. M. Haenen. 2023. "Molecular Characterization of Serratia marcescens Strain Isolated from Yellow Mealworms, Tenebrio molitor, in The Netherlands" Insects 14, no. 9: 770. https://doi.org/10.3390/insects14090770