
Effects of Temperature and Density on House Cricket Survival and Growth and on the Prevalence of Acheta Domesticus Densovirus
 , ,
, ,
Abstract
:Simple Summary
Abstract
1. Introduction
2. Materials and Methods
2.1. Cricket Rearing and Experimental Units
2.2. DNA Extraction and qPCR Analysis
2.3. Data Analyses and Visualization
3. Results
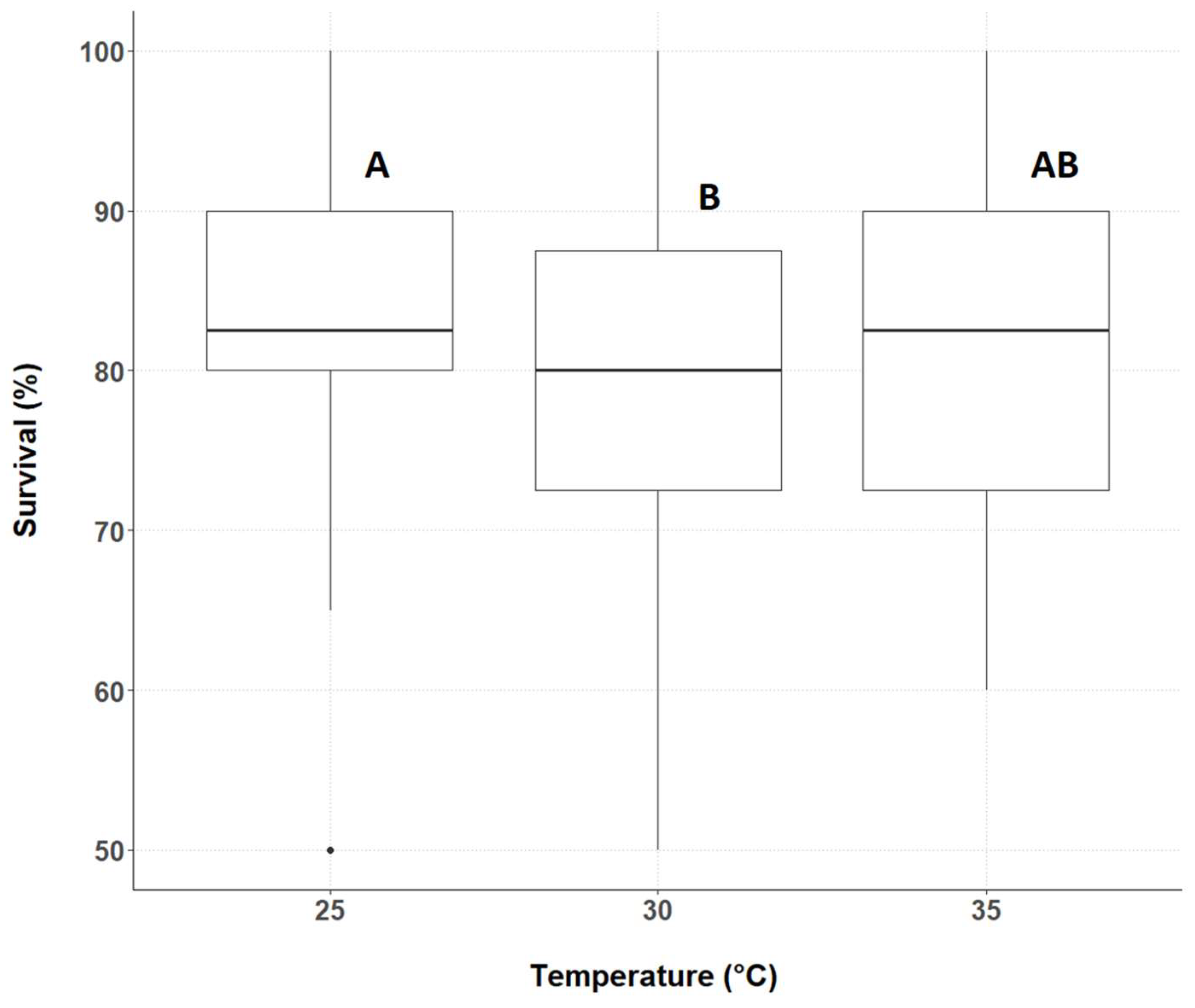
3.1. Cricket Survival
3.2. Cricket Biomass
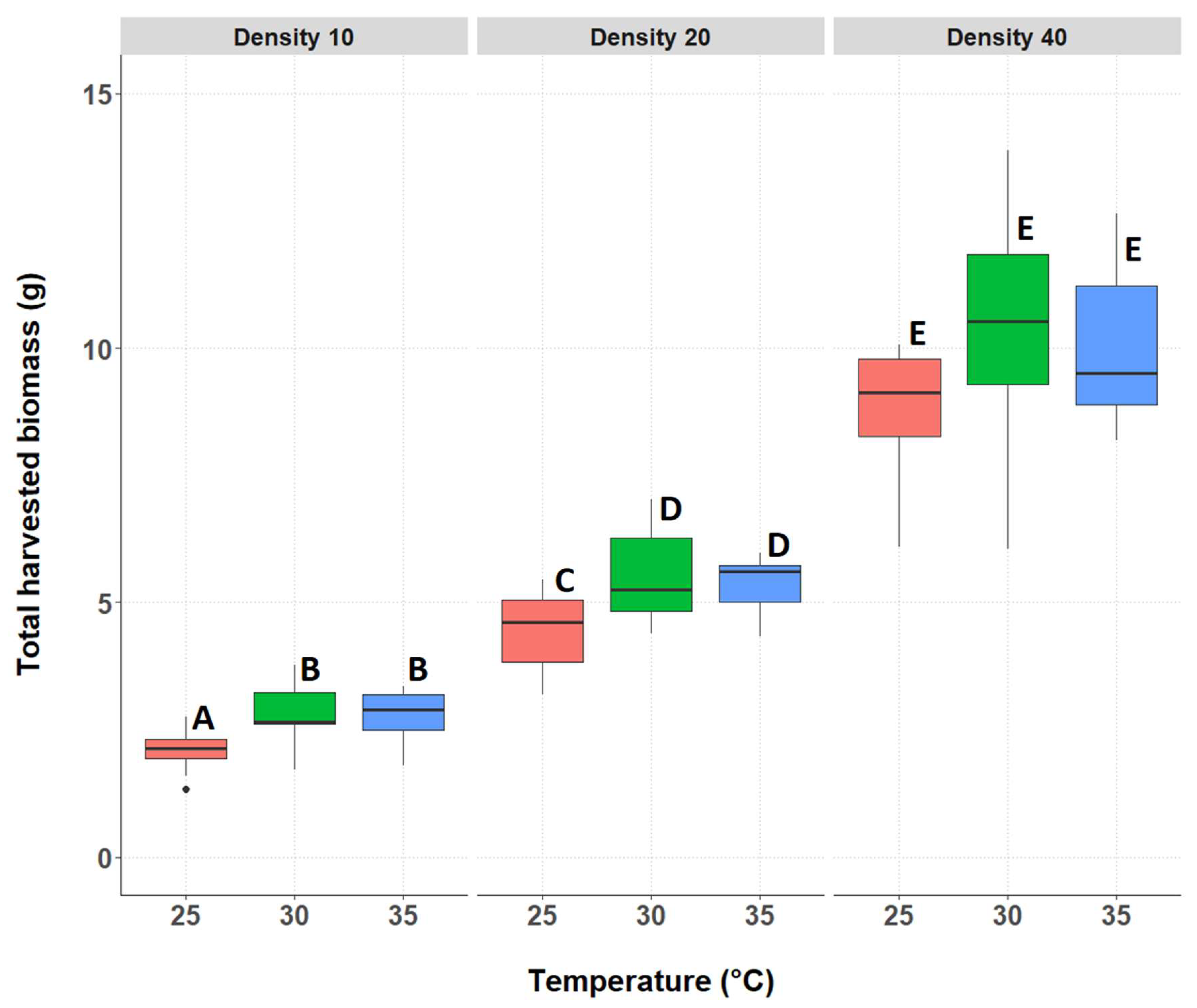
3.2.1. Total Biomass of the Surviving Individuals
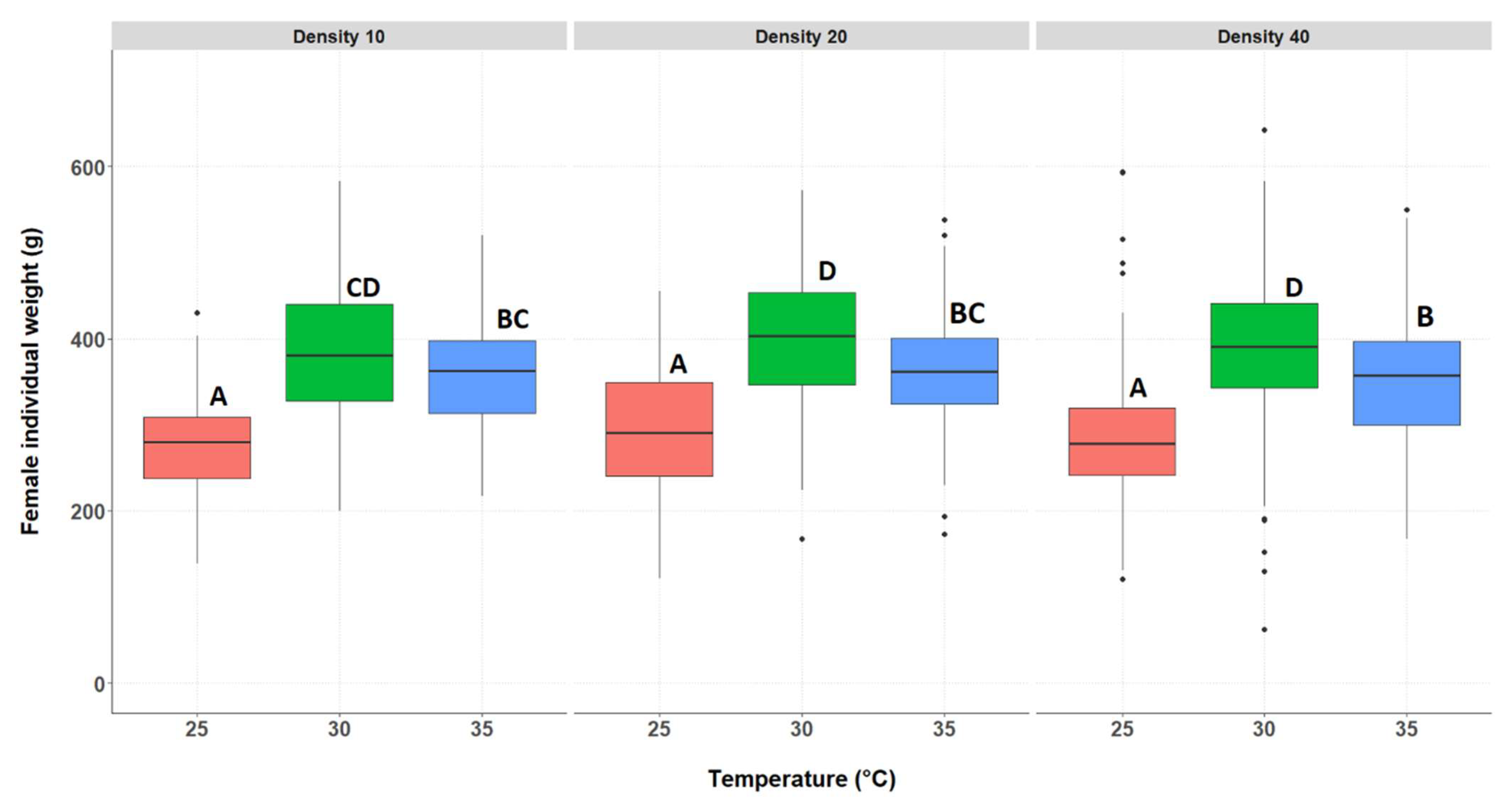
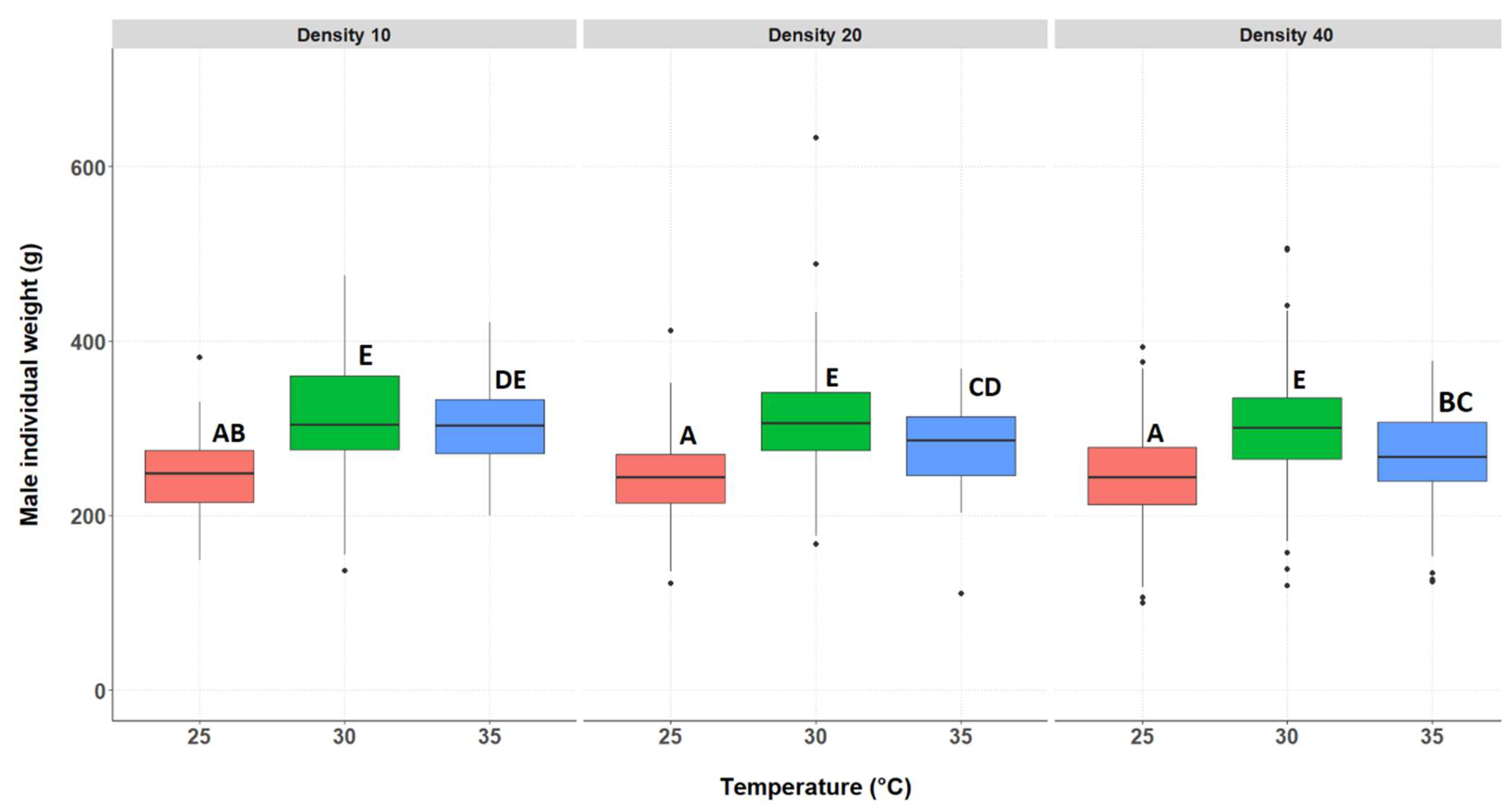
3.2.2. Individual Weight of the Surviving Crickets, Separated by Genders
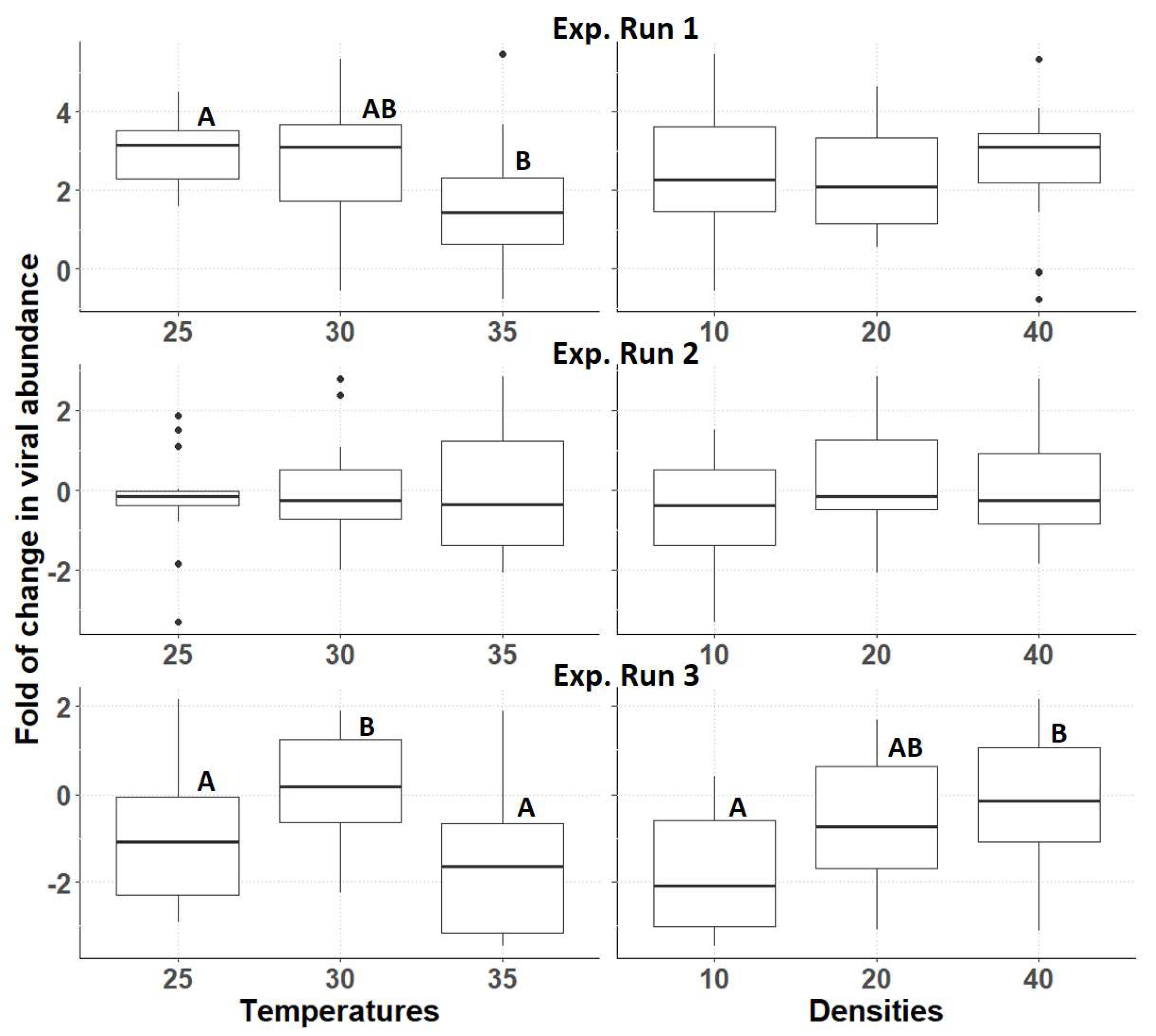
3.3. Relative Abundance of AdDV
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Mc Carthy, U.; Uysal, I.; Badia-Melis, R.; Mercier, S.; O’Donnell, C.; Ktenioudaki, A. Global food security—Issues, challenges and technological solutions. Trends Food Sci. Technol. 2018, 77, 11–20. [Google Scholar] [CrossRef]
- Van Huis, A.; Tomberlin, J.K. Insects as Food and Feed: From Production to Consumption; Wageningen Academic Press: Wageningen, The Netherlands, 2017. [Google Scholar]
- Van Huis, A.; Oonincx, D.G.A.B. The environmental sustainability of insects as food and feed. A review. Agron. Sustain. Dev. 2017, 37, 1–14. [Google Scholar] [CrossRef] [Green Version]
- Oonincx, D.G.; van Broekhoven, S.; van Huis, A.; van Loon, J.J. Feed conversion, survival and development, and composition of four insect species on diets composed of food by-products. PLoS ONE 2015, 10, e0144601. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Smetana, S.; Mathys, A.; Knoch, A.; Heinz, V. Meat alternatives: Life cycle assessment of most known meat substitutes. Int. J. Life Cycle Assess. 2015, 20, 1254–1267. [Google Scholar] [CrossRef]
- van Huis, A. Edible crickets, but which species? J. Insects Food Feed 2020, 6, 91–94. [Google Scholar] [CrossRef]
- Halloran, A.; Hanboonsong, Y.; Roos, N.; Bruun, S. Life cycle assessment of cricket farming in north-eastern Thailand. J. Clean. Prod. 2017, 156, 83–94. [Google Scholar] [CrossRef]
- Mariod, A.A.; Saeed Mirghani, M.E.; Hussein, I. (Eds.) Acheta domesticus House Cricket. In Unconventional Oilseeds and Oil Sources; Elsevier Inc.: Amsterdam, The Netherlands, 2017; pp. 323–325. [Google Scholar]
- Montowska, M.; Kowalczewski, P.L.; Rybicka, I.; Fornal, E. Nutritional value, protein and peptide composition of edible cricket powders. Food Chem. 2019, 289, 130–138. [Google Scholar] [CrossRef]
- Udomsil, N.; Imsoonthornruksa, S.; Gosalawit, C.; Ketudat-Cairns, M. Nutritional values and functional properties of house cricket (Acheta domesticus) and field cricket (Gryllus bimaculatus). Food Sci. Technol. Res. 2019, 25, 597–605. [Google Scholar] [CrossRef]
- Osimani, A.; Milanovic, V.; Cardinali, F.; Roncolini, A.; Garofalo, C.; Clementi, F.; Pasquini, M.; Mozzon, M.; Foligni, R.; Raffaelli, N.; et al. Bread enriched with cricket powder (Acheta domesticus): A technological, microbiological and nutritional evaluation. Innov. Food Sci. Emerg. Technol. 2018, 48, 150–163. [Google Scholar] [CrossRef]
- Szelei, J.; Woodring, J.; Goettel, M.S.; Duke, G.; Jousset, F.X.; Liu, K.Y.; Zadori, Z.; Li, Y.; Styer, E.; Boucias, D.G.; et al. Susceptibility of North-American and European crickets to Acheta domesticus densovirus (AdDNV) and associated epizootics. J. Invertebr. Pathol. 2011, 106, 394–399. [Google Scholar] [CrossRef]
- Maciel-Vergara, G.; Ros, V.I.D. Viruses of insects reared for food and feed. J. Invertebr. Pathol. 2017, 147, 60–75. [Google Scholar] [CrossRef]
- Liu, K.; Li, Y.; Jousset, F.X.; Zadori, Z.; Szelei, J.; Yu, Q.; Pham, H.T.; Lepine, F.; Bergoin, M.; Tijssen, P. The Acheta domesticus densovirus, isolated from the European house cricket, has evolved an expression strategy unique among parvoviruses. J. Virol. 2011, 85, 10069–10078. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Weissman, D.B.; Gray, D.A.; Thi Pham, H.; Tijssen, P. Billions and billions sold: Pet-feeder crickets (Orthoptera: Gryllidae), commercial cricket farms, an epizootic densovirus, and government regulations make for a potential disaster. Zootaxa 2012, 3504, 67–88. [Google Scholar] [CrossRef]
- Styer, E.L.; Hamm, J.J. Report of a Densovirus in a Commercial Cricket Operation in the Southeastern United-States. J. Invertebr. Pathol. 1991, 58, 283–285. [Google Scholar] [CrossRef]
- Fédière, G. Epidemiology and pathology of Densovirinae. Contrib. Microbiol. 2000, 4, 1–11. [Google Scholar]
- Duffield, K.R.; Hunt, J.; Sadd, B.M.; Sakaluk, S.K.; Oppert, B.; Rosario, K.; Behle, R.W.; Ramirez, J.L. Active and Covert Infections of Cricket Iridovirus and Acheta domesticus Densovirus in Reared Gryllodes sigillatus Crickets. Front. Microbiol. 2021, 12, 780796. [Google Scholar] [CrossRef] [PubMed]
- Semberg, E.; de Miranda, J.R.; Low, M.; Jansson, A.; Forsgren, E.; Berggren, A. Diagnostic protocols for the detection of Acheta domesticus densovirus (AdDV) in cricket frass. J. Virol. Methods 2019, 264, 61–64. [Google Scholar] [CrossRef] [PubMed]
- Eilenberg, J.; Vlak, J.M.; Nielsen-LeRoux, C.; Cappellozza, S.; Jensen, A.B. Diseases in insects produced for food and feed. J. Insects Food Feed 2015, 1, 87–102. [Google Scholar] [CrossRef] [Green Version]
- Hoffmann, K.H. Environmental Physiology and Biochemistry of Insects; Springer: Berlin/Heidelberg, Germany, 1985. [Google Scholar]
- Chown, S.L.; Chown, S.; Nicolson, S. Insect Physiological Ecology: Mechanisms and Patterns; Oxford University Press: Oxford, UK, 2004. [Google Scholar]
- Dixon, A.F.G.; Honek, A.; Keil, P.; Kotela, M.A.A.; Sizling, A.L.; Jarosik, V. Relationship between the minimum and maximum temperature thresholds for development in insects. Funct. Ecol. 2009, 23, 257–264. [Google Scholar] [CrossRef]
- Clifford, C.W.; Woodring, J.P. Methods for rearing the house cricket, Acheta domesticus (L.), along with baseline values for feeding rates, growth rates, development times, and blood composition. J. Appl. Entomol. 1990, 109, 1–14. [Google Scholar] [CrossRef]
- Neven, L.G. Physiological responses of insects to heat. Postharvest Biol. Technol. 2000, 21, 103–111. [Google Scholar] [CrossRef]
- Kok, R. Preliminary project design for insect production: Part 4—facility considerations. J. Insects Food Feed 2021, 7, 541–551. [Google Scholar] [CrossRef]
- Kieruzel, M. The selection of shelter place by the house cricket. Acta Neurobiol. Exp. (Wars) 1976, 36, 561–580. [Google Scholar]
- Tennis, P.S.; Koonce, J.F.; Teraguchi, M. The effects of population density and food surface area on body weight of Acheta domesticus (L.) (Orthoptera: Gryllidae). Can. J. Zool. 1977, 55, 2004–2010. [Google Scholar] [CrossRef]
- Barragan-Fonseca, K.B.; Dicke, M.; van Loon, J.J.A. Influence of larval density and dietary nutrient concentration on performance, body protein, and fat contents of black soldier fly larvae (Hermetia illucens). Entomol. Exp. Appl. 2018, 166, 761–770. [Google Scholar] [CrossRef] [Green Version]
- Deruytter, D.; Coudron, C.L.; Claeys, J. The Effects of Density on the Growth and Temperature Production of Tenebrio molitor Larvae. Sustainability 2022, 14, 6234. [Google Scholar] [CrossRef]
- Kuriwada, T.; Kumano, N.; Shiromoto, K.; Haraguchi, D. High Population Density and Egg Cannibalism Reduces the Efficiency of Mass-Rearing in Euscepes postfasciatus (Coleoptera: Curculionidae). Fla. Entomol. 2009, 92, 221–228. [Google Scholar] [CrossRef]
- Mahavidanage, S.; Fuciarelli, T.M.; Li, X.; Rollo, C.D. The effects of rearing density on growth, survival, and starvation resistance of the house cricket Acheta domesticus. J. Orthoptera Res. 2023, 32, 25–31. [Google Scholar] [CrossRef]
- Morales-Ramos, J.A.; Rojas, M.G.; Dossey, A.T.; Berhow, M. Self-selection of food ingredients and agricultural by-products by the house cricket, Acheta domesticus (Orthoptera: Gryllidae): A holistic approach to develop optimized diets. PLoS ONE 2020, 15, e0227400. [Google Scholar] [CrossRef]
- Untergasser, A.; Cutcutache, I.; Koressaar, T.; Ye, J.; Faircloth, B.C.; Remm, M.; Rozen, S.G. Primer3—New capabilities and interfaces. Nucleic Acids Res. 2012, 40, e115. [Google Scholar] [CrossRef] [Green Version]
- Lee, C.; Kim, J.; Shin, S.G.; Hwang, S. Absolute and relative QPCR quantification of plasmid copy number in Escherichia coli . J. Biotechnol. 2006, 123, 273–280. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hellemans, J.; Mortier, G.; De Paepe, A.; Speleman, F.; Vandesompele, J. qBase relative quantification framework and software for management and automated analysis of real-time quantitative PCR data. Genome Biol. 2007, 8, R19. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- R Core Team. R: A Language and Environment for Statistical Computing; R Foundation for Statistical Computing: Vienna, Austria, 2020. [Google Scholar]
- Shapiro, S.S.; Wilk, M.B. An Analysis of Variance Test for Normality (Complete Samples). Biometrika 1965, 52, 591–611. [Google Scholar] [CrossRef]
- Bates, D.; Mächler, M.; Bolker, B.; Walker, S. Fitting Linear Mixed-Effects Models Usinglme4. J. Stat. Softw. 2015, 67, 6–12. [Google Scholar] [CrossRef]
- Burnham, K.P.; Anderson, D.R. Model Selection and Multimodel Inference: A Practical Information-Theoretic Approach, 2nd ed.; Springer: New York, NY, USA, 2002. [Google Scholar]
- Searle, S.R.; Speed, F.M.; Milliken, G.A. Population Marginal Means in the Linear-Model—An Alternative to Least-Squares Means. Am. Stat. 1980, 34, 216–221. [Google Scholar]
- Wickham, H. ggplot2: Elegant Graphics for Data Analysis; Springer International Publishing: Berlin/Heidelberg, Germany, 2016. [Google Scholar]
- Saska, P.; van der Werf, W.; Hemerik, L.; Luff, M.L.; Hatten, T.D.; Honek, A.; Pocock, M. Temperature effects on pitfall catches of epigeal arthropods: A model and method for bias correction. J. Appl. Ecol 2013, 50, 181–189. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Chia, S.Y.; Tanga, C.M.; Khamis, F.M.; Mohamed, S.A.; Salifu, D.; Sevgan, S.; Fiaboe, K.K.M.; Niassy, S.; van Loon, J.J.A.; Dicke, M.; et al. Threshold temperatures and thermal requirements of black soldier fly Hermetia illucens: Implications for mass production. PLoS ONE 2018, 13, e0206097. [Google Scholar] [CrossRef] [Green Version]
- Lazzaro, B.P.; Flores, H.A.; Lorigan, J.G.; Yourth, C.P. Genotype-by-environment interactions and adaptation to local temperature affect immunity and fecundity in Drosophila melanogaster. PLoS Pathog. 2008, 4, e1000025. [Google Scholar] [CrossRef] [Green Version]
- Richardson, J.M.L.; Baker, R.L. Effect of Body Size and Feeding on Fecundity in the Damselfly Ischnura verticalis (Odonata: Coenagrionidae). Oikos 1997, 79, 477–483. [Google Scholar] [CrossRef]
- Stahlschmidt, Z.R.; Rollinson, N.; Acker, M.; Adamo, S.A.; Martin, L. Are all eggs created equal? Food availability and the fitness trade-off between reproduction and immunity. Funct. Ecol. 2013, 27, 800–806. [Google Scholar] [CrossRef]
- de Miranda, J.R.; Granberg, F.; Low, M.; Onorati, P.; Semberg, E.; Jansson, A.; Berggren, A. Virus Diversity and Loads in Crickets Reared for Feed: Implications for Husbandry. Front. Vet. Sci. 2021, 8, 642085. [Google Scholar] [CrossRef] [PubMed]
- Steinhaus, E.A. Crowding as a Possible Stress Factor in Insect Disease. Ecology 1958, 39, 503–514. [Google Scholar] [CrossRef]
- Guo-Ping, K.; Xi-Jie, G. Overview of silkworm pathology in China. Afr. J. Biotechnol. 2011, 10, 18046–18056. [Google Scholar] [CrossRef]
- Genersch, E. Honey bee pathology: Current threats to honey bees and beekeeping. Appl. Microbiol. Biotechnol. 2010, 87, 87–97. [Google Scholar] [CrossRef] [PubMed]
- Perrin, A.; Gosselin-Grenet, A.S.; Rossignol, M.; Ginibre, C.; Scheid, B.; Lagneau, C.; Chandre, F.; Baldet, T.; Ogliastro, M.; Bouyer, J. Variation in the susceptibility of urban Aedes mosquitoes infected with a densovirus. Sci. Rep. 2020, 10, 18654. [Google Scholar] [CrossRef] [PubMed]
- Williams, T.; Hernandez, O. Costs of cannibalism in the presence of an iridovirus pathogen of Spodoptera frugiperda. Ecol. Entomol. 2006, 31, 106–113. [Google Scholar] [CrossRef] [Green Version]
- Grispoldi, L.; Karama, M.; El-Ashram, S.; Saraiva, C.M.; Garcia-Diez, J.; Chalias, A.; Barbera, S.; Cenci-Goga, B.T. Hygienic Characteristics and Detection of Antibiotic Resistance Genes in Crickets (Acheta domesticus) Breed for Flour Production. Microbiol. Res. 2021, 12, 503–512. [Google Scholar] [CrossRef]
- Maciel-Vergara, G.; Jensen, A.B.; Eilenberg, J. Cannibalism as a Possible Entry Route for Opportunistic Pathogenic Bacteria to Insect Hosts, Exemplified by Pseudomonas aeruginosa, a Pathogen of the Giant Mealworm Zophobas morio. Insects 2018, 9, 88. [Google Scholar] [CrossRef] [Green Version]
- Wang, Y.; Gosselin Grenet, A.S.; Castelli, I.; Cermenati, G.; Ravallec, M.; Fiandra, L.; Debaisieux, S.; Multeau, C.; Lautredou, N.; Dupressoir, T.; et al. Densovirus crosses the insect midgut by transcytosis and disturbs the epithelial barrier function. J. Virol. 2013, 87, 12380–12391. [Google Scholar] [CrossRef] [Green Version]
- Kortsmit, Y.; van der Bruggen, M.; Wertheim, B.; Dicke, M.; Beukeboom, L.W.; van Loon, J.J.A. Behaviour of two fly species reared for livestock feed: Optimising production and insect welfare. J. Insects Food Feed 2023, 9, 149–169. [Google Scholar] [CrossRef]
- Ye, J.W.; Li, J. Factors affecting cannibalism by Mallada basalis. Biocontrol Sci. Technol. 2020, 30, 442–450. [Google Scholar] [CrossRef]
- Kakimoto, T.; Fujisaki, K.; Miyatake, T. Egg laying preference, larval dispersion, and cannibalism in Helicoverpa armigera (Lepidoptera: Noctuidae). Ann. Entomol. Soc. Am. 2003, 96, 793–798. [Google Scholar] [CrossRef]
- Lee, K.P.; Simpson, S.J.; Wilson, K. Dietary protein-quality influences melanization and immune function in an insect. Funct. Ecol. 2008, 22, 1052–1061. [Google Scholar] [CrossRef]
- Wilson, J.K.; Ruiz, L.; Davidowitz, G. Dietary Protein and Carbohydrates Affect Immune Function and Performance in a Specialist Herbivore Insect (Manduca sexta). Physiol. Biochem. Zool. 2019, 92, 58–70. [Google Scholar] [CrossRef] [Green Version]
- Harsányi, E.; Juhász, C.; Kovács, E.; Huzsvai, L.; Pintér, R.; Fekete, G.; Varga, Z.I.; Aleksza, L.; Gyuricza, C. Evaluation of Organic Wastes as Substrates for Rearing Zophobas morio, Tenebrio molitor, and Acheta domesticus Larvae as Alternative Feed Supplements. Insects 2020, 11, 604. [Google Scholar] [CrossRef]
- Bawa, M.; Songsermpong, S.; Kaewtapee, C.; Chanput, W. Effect of Diet on the Growth Performance, Feed Conversion, and Nutrient Content of the House Cricket. J. Insect Sci. 2020, 20, 1–10. [Google Scholar] [CrossRef]






{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Name | Reference | Primer Sequence 5′ to 3′ (Forward/Reverse) | Amplicon Size (bp) | Gene Target |
|---|---|---|---|---|
| EF1 | This study | GGAAATCAAGAAGGAAGTCAGC/GGCATCCAAAGCCTCAATAAG | 199 | A. domesticus elongation factor 1 alpha gene |
| AdDV 96 | Duffield et al. [18] | GCGAGCAATCCCGACTACTA/CGCGTTGTTGATGTCCTTCC | 96 | Non-structural protein |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Takacs, J.; Bryon, A.; Jensen, A.B.; van Loon, J.J.A.; Ros, V.I.D. Effects of Temperature and Density on House Cricket Survival and Growth and on the Prevalence of Acheta Domesticus Densovirus. Insects 2023, 14, 588. https://doi.org/10.3390/insects14070588
Takacs J, Bryon A, Jensen AB, van Loon JJA, Ros VID. Effects of Temperature and Density on House Cricket Survival and Growth and on the Prevalence of Acheta Domesticus Densovirus. Insects. 2023; 14(7):588. https://doi.org/10.3390/insects14070588
Chicago/Turabian StyleTakacs, Jozsef, Astrid Bryon, Annette B. Jensen, Joop J. A. van Loon, and Vera I. D. Ros. 2023. "Effects of Temperature and Density on House Cricket Survival and Growth and on the Prevalence of Acheta Domesticus Densovirus" Insects 14, no. 7: 588. https://doi.org/10.3390/insects14070588