
Signal and Nutritional Effects of Mixed Diets on Reproduction of a Predatory Ladybird, Cheilomenes propinqua

 , , and
, , and
Abstract
:Simple Summary
Abstract
1. Introduction
2. Materials and Methods
2.1. Insects
2.2. Experimental Design
- Not developed (oocytes are not visible; follicles are absent; no or hardly any constrictions of ovarioles);
- Poorly developed (follicles are few and small; ovarioles are somewhat constricted around the follicles);
- Moderately developed (follicles are of medium size and become vitellinized; ovarioles are markedly swollen; the largest oocyte is filled with whitish yolk);
- Fully developed (follicles are vitellinized; mature oocytes are present; the largest oocyte reached the maximum size).
- Not developed (practically inconspicuous);
- Poorly developed (composed of small scattered globules interspersed around the internal organs);
- Moderately developed (internal organs are partially hidden by the fat body composed of well-formed globules);
- Fully developed (internal organs are completely hidden by the fat body composed of well-formed globules and interconnected stringy lobes).
- Reproductively active (ovaries are fully developed).
- Intermediate (ovaries are moderately developed);
- Diapausing (ovaries are not or poorly developed; the fat body is moderately or fully developed);
- Underdeveloped (both ovaries and fat body are not or poorly developed).
- The duration of the pre-oviposition period (from adult emergence to the first oviposition, only for females that laid eggs);
- Total fecundity (the number of eggs laid during the observation period);
- Oviposition intensity (the mean number of eggs laid daily calculated for the period from the onset of egg-laying to the end of observation);
- The degree of ovaries development at dissection;
- The degree of fat body development at dissection;
- The reproductive state at dissection;
- The signs of resorption at dissection;
- The size of the largest follicle at dissection (for reproductively active females);
- The onset of egg-laying (yes or no).
2.3. Statistical Analysis
3. Results
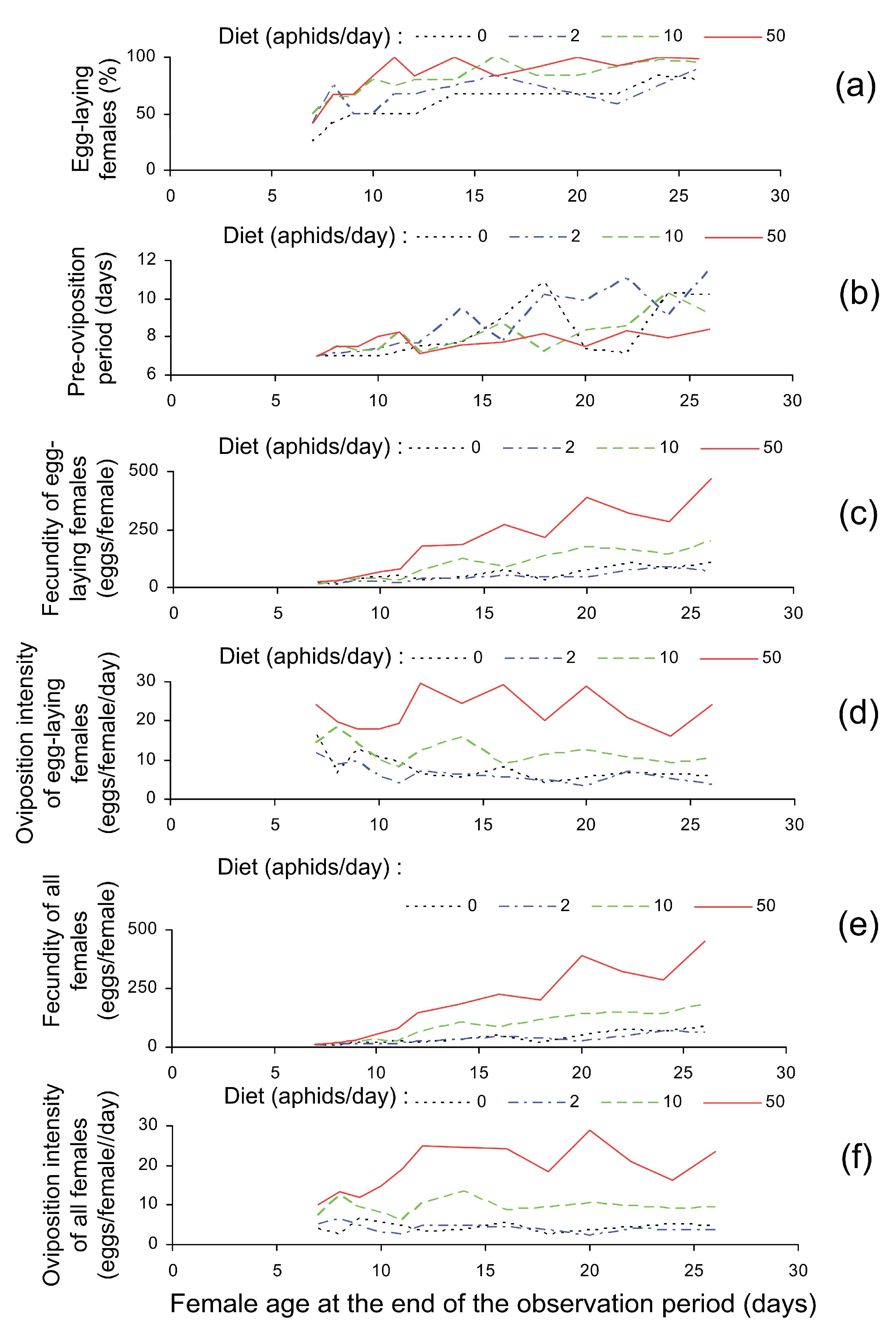
3.1. Reproduction Parameters
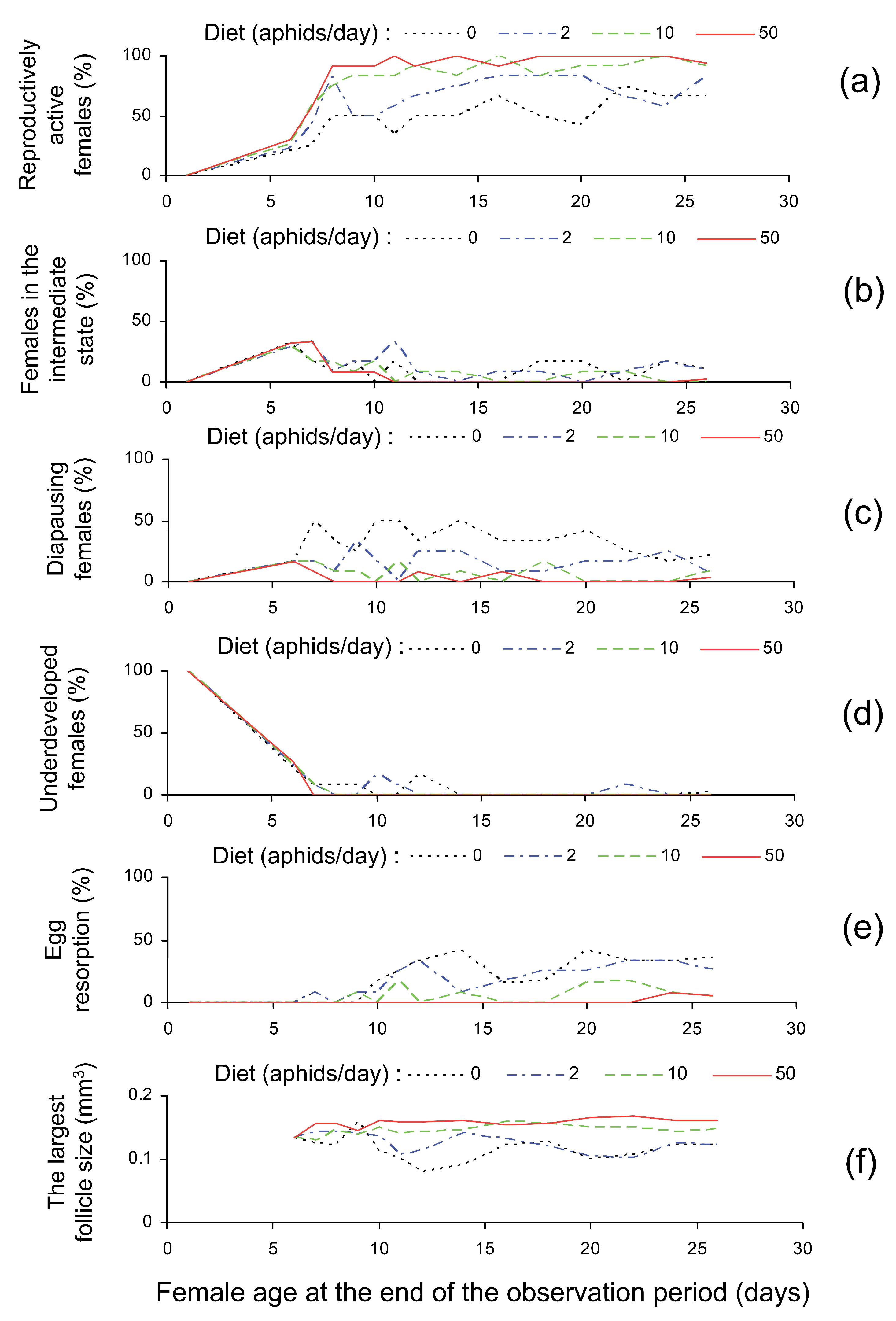
3.2. Dissection Results
4. Discussion
Author Contributions
Funding
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Van Lenteren, J.C. IOBC Internet Book of Biological Control, Version 6. 2012. Available online: https://www.iobc-global.org/publications_iobc_internet_book_of_biological_control.html (accessed on 25 May 2023).
- Van Lenteren, J.C.; Alomar, O.; Ravensberg, W.J.; Urbaneja, A. Biological Control Agents for Control of Pests in Greenhouses. In Integrated Pest and Disease Management in Greenhouse Crops; Gullino, M.L., Albajes, R., Nicot, P.C., Eds.; Springer: Cham, Switzerland, 2020; pp. 409–439. [Google Scholar]
- Van Lenteren, J.C.; Bueno, V.H.; Klapwijk, J.N. Augmentative Biological Control. In Biological Control: Global Impacts, Challenges and Future Directions of Pest Management; Mason, P.G., Ed.; Csiro Publishing: Clayton, VIC, Australia, 2021; pp. 90–109. [Google Scholar]
- Parrella, M.P.; Lewis, E. Biological control in greenhouse and nursery production: Present status and future directions. Am. Entomol. 2017, 63, 237–250. [Google Scholar] [CrossRef]
- Michaud, J.P. Coccinellids in Biological Control. In Ecology and Behaviour of the Ladybird Beetles (Coccinellidae); Hodek, I., van Emden, H.F., Honěk, A., Eds.; John Wiley & Sons: Chichester, UK, 2012; pp. 488–519. [Google Scholar]
- Hodek, I. Food Relationships. In Ecology of Coccinellidae; Hodek, I., Honěk, A., Eds.; Kluwer: Dordrecht, The Netherlands, 1996; pp. 143–238. [Google Scholar]
- Hodek, I.; Evans, E.W. Food Relationships. In Ecology and Behaviour of the Ladybird Beetles (Coccinellidae); Hodek, I., van Emden, H.F., Honěk, A., Eds.; Wiley-Blackwell: Chichester, UK, 2012; pp. 141–274. [Google Scholar]
- Nedvěd, O.; Honěk, A. Life History and Development. In Ecology and Behaviour of the Ladybird Beetles (Coccinellidae); Hodek, I., van Emden, H.F., Honěk, A., Eds.; Wiley-Blackwell: Chichester, UK, 2012; pp. 54–109. [Google Scholar]
- Chen, J.; Qin, Q.J.; Liu, S.; He, Y.Z. Effect of six diets on development and fecundity of Harmonia axyridis (Pallas) (Coleoptera: Coccinellidae). Afr. Entomol. 2012, 20, 76–84. [Google Scholar] [CrossRef]
- Mirhosseini, M.A.; Hosseini, M.R.; Jalali, M.A. Effects of diet on development and reproductive fitness of two predatory coccinellids (Coleoptera: Coccinellidae). Eur. J. Entomol. 2015, 112, 446–452. [Google Scholar] [CrossRef] [Green Version]
- De Castro-Guedes, C.F.; de Almeida, L.M.; do Rocio Chiarello Penteado, S.; Moura, M.O. Effect of different diets on biology, reproductive variables and life and fertility tables of Harmonia axyridis (Pallas) (Coleoptera, Coccinellidae). Rev. Bras. Entomol. 2016, 60, 260–266. [Google Scholar] [CrossRef] [Green Version]
- Rojas, M.G.; Morales-Ramos, J.A.; Riddick, E.W. Use of Tenebrio molitor (Coleoptera: Tenebrionidae) powder to enhance artificial diet formulations for Coleomegilla maculata (Coleoptera: Coccinellidae). Biol. Control 2016, 100, 70–78. [Google Scholar] [CrossRef] [Green Version]
- Hodek, I. Diapause/Dormancy. In Ecology and Behaviour of the Ladybird Beetles (Coccinellidae); Hodek, I., van Emden, H.F., Honěk, A., Eds.; Wiley-Blackwell: Chichester, UK, 2012; pp. 275–342. [Google Scholar]
- Horton, D.R.; Hinojosa, T.; Olson, S.R. Effects of photoperiod and prey type on diapause tendency and preoviposition period in Perillus bioculatus (Hemiptera: Pentatomidae). Can. Entomol. 1998, 130, 315–320. [Google Scholar] [CrossRef]
- Zaslavskiy, V.A.; Semyanov, V.P.; Vaghina, N.P. Food as a cue factor controlling adult diapause in the lady beetle, Harmonia sedecimnotata (Coleoptera, Coccinellidae). Entomol. Rev. 1998, 78, 774–779. [Google Scholar]
- Michaud, J.P.; Qureshi, J.A. Induction of reproductive diapause in Hippodamia convergens (Coleoptera: Coccinellidae) hinges on prey quality and availability. Eur. J. Entomol. 2005, 102, 483–487. [Google Scholar] [CrossRef] [Green Version]
- Reznik, S.Y.; Vaghina, N.P. Dynamics of fat content during induction and termination of “trophic diapause” in Harmonia sedecimnotata Fabr. females (Coleoptera, Coccinellidae). Entomol. Rev. 2006, 86, 125–132. [Google Scholar] [CrossRef]
- Reznik, S.Y.; Vaghina, N.P. Effects of photoperiod and diet on diapause tendency, maturation and fecundity in Harmonia axyridis (Coleoptera: Coccinellidae). J. Appl. Entomol. 2013, 137, 452–461. [Google Scholar] [CrossRef]
- Berkvens, N.; Bonte, J.; Berkvens, D.; Tirry, L.; De Clercq, P. Influence of diet and photoperiod on development and reproduction of European populations of Harmonia axyridis (Pallas) (Coleoptera: Coccinellidae). BioControl 2008, 53, 211–221. [Google Scholar] [CrossRef]
- Reznik, S.; Ovchinnikov, A.; Ovchinnikova, A.; Bezman-Moseyko, O.; Belyakova, N. Photoperiodic, thermal and trophic responses of a predatory ladybird Cheilomenes propinqua. J. Appl. Entomol. 2021, 145, 134–144. [Google Scholar] [CrossRef]
- Ohgushi, T.; Sawada, H. Population equilibrium with respect to a variable food resource and its behavioural basis in a herbivorous lady beetle, Henosepilachna niponica. J. Animal Ecol. 1985, 54, 781–796. [Google Scholar] [CrossRef]
- Ohgushi, T. A reproductive tradeoff in an herbivorous lady beetle: Egg resorption and female survival. Oecologia 1996, 106, 345–351. Available online: https://link.springer.com/article/10.1007/BF00334562 (accessed on 21 June 2023). [CrossRef] [PubMed]
- Pervez, A. Ecology and Behaviour of Predaceous Ladybird Beetles (Coleoptera: Coccinellidae). In Biodiversity, Environment and Ecosystem Services; Arya, M.K., Ed.; Discovery Publishing House: New Delhi, India, 2023; pp. 12–29. [Google Scholar]
- Racioppi, J.V.; Burton, R.L.; Eikenbary, R. The effects of various oligidic synthetic diets on the growth of Hippodamia convergens. Entomol. Exp. Appl. 1981, 30, 68–72. [Google Scholar] [CrossRef]
- Evans, E.W. Egg production in response to combined alternative foods by the predator Coccinella transversalis. Entomol. Exp. Appl. 2000, 94, 141–147. [Google Scholar] [CrossRef] [Green Version]
- Evans, E.W. Searching and reproductive behaviour of female aphidophagous ladybirds (Coleoptera: Coccinellidae): A review. Eur. J. Entomol. 2003, 100, 1–10. Available online: https://www.eje.cz/pdfs/eje/2003/01/01.pdf (accessed on 21 June 2023). [CrossRef]
- Evans, E.W.; Gunther, D.I. The link between food and reproduction in aphidophagous predators: A case study with Harmonia axyridis (Coleoptera: Coccinellidae). Eur. J. Entomol. 2005, 102, 423–430. Available online: https://www.eje.cz/pdfs/eje/2005/03/13.pdf (accessed on 21 June 2023). [CrossRef] [Green Version]
- Evans, E.W.; Stevenson, A.T.; Richards, D.R. Essential versus alternative foods of insect predators: Benefits of a mixed diet. Oecologia 1999, 121, 107–112. [Google Scholar] [CrossRef]
- Nielsen, F.H.; Hauge, M.S.; Toft, S. The influence of mixed aphid diets on larval performance of Coccinella septempunctata (Col., Coccinellidae). J. Appl. Entomol. 2002, 126, 194–197. [Google Scholar] [CrossRef] [Green Version]
- Snyder, W.E.; Clevenger, G.M. Negative dietary effects of Colorado potato beetle eggs for the larvae of native and introduced ladybird beetles. Biol. Control 2004, 31, 353–361. [Google Scholar] [CrossRef]
- Sighinolfi, L.; Febvay, G.; Dindo, M.L.; Rey, M.; Pageaux, J.; Baronio, P.; Grenier, S. Biological and biochemical characteristics for quality control of Harmonia axyridis (Pallas) (Coleoptera, Coccinellidae) reared on a liver-based diet. Arch. Insect Biochem. Physiol. 2008, 68, 26–39. [Google Scholar] [CrossRef] [PubMed]
- Soares, A.O.; Coderre, D.; Schanderl, H. Dietary self-selection behaviour by the adults of the aphidophagous ladybeetle Harmonia axyridis (Coleoptera: Coccinellidae). J. Anim. Ecol. 2004, 73, 478–486. [Google Scholar] [CrossRef]
- Phoofolo, M.W.; Giles, K.L.; Elliott, N.C. Quantitative evaluation of suitability of the greenbug, Schizaphis graminum, and the bird cherry-oat aphid, Rhopalosiphum padi, as prey for Hippodamia convergens (Coleoptera: Coccinellidae). Biol. Control 2007, 41, 25–32. [Google Scholar] [CrossRef]
- Berkvens, N.; Bonte, J.; Berkvens, D.; Deforce, K.; Tirry, L.; De Clercq, P. Pollen as an alternative food for Harmonia axyridis. BioControl 2008, 53, 201–210. [Google Scholar] [CrossRef]
- Bonte, M.; Samih, M.A.; De Clercq, P. Development and reproduction of Adalia bipunctata on factitious and artificial foods. BioControl 2010, 55, 485–491. [Google Scholar] [CrossRef]
- Sayed, S.M.; El Arnaouty, S.A. Effect of corn pollens, as supplemental food, on development and reproduction of the predatory species, Hippodamia variegata (Goeze) (Coleoptera: Coccinellidae). Egypt. J. Biol. Pest Control 2016, 26, 457–461. [Google Scholar]
- Hassani, F.; Shirvani, A.; Rashki, M. Fitness evaluation of Oenopia conglobata contaminata (Menetries) (Col.: Coccinellidae) fed on different diets. Acta Biol. Slov. 2019, 62, 35–43. [Google Scholar]
- de Lima, M.S.; Pontes, W.J.T.; de Lucena Nóbrega, R. Pollen did not provide suitable nutrients for ovary development in a ladybird Brumoides foudrasii (Coleoptera: Coccinellidae). Divers. J. 2020, 5, 1486–1494. [Google Scholar] [CrossRef]
- Li, D.; Chen, P.; Liu, J.; Chi, B.; Zhang, X.; Liu, Y. Artificial rearing of the ladybird Propylea japonica on a diet containing oriental armyworm, Mythimna separata. Entomol. Exp. Appl. 2021, 169, 472–479. [Google Scholar] [CrossRef]
- Stowe, H.E.; Michaud, J.P.; Kim, T. The benefits of omnivory for reproduction and life history of a specialized aphid predator, Hippodamia convergens (Coleoptera: Coccinellidae). Environ. Entomol. 2021, 50, 69–75. [Google Scholar] [CrossRef] [PubMed]
- Vahmani, A.; Shirvani, A.; Rashki, M. Biological parameters of Oenopia conglobata (Linnaeus) (Coleoptera: Coccinellidae) feeding on different diets. Int. J. Trop. Insect Sci. 2022, 42, 2241–2247. [Google Scholar] [CrossRef]
- Toft, S. The quality of aphids as food for generalist predators: Implications for natural control of aphids. Eur. J. Entomol. 2005, 102, 371–383. Available online: https://www.eje.cz/pdfs/eje/2005/03/07.pdf (accessed on 21 June 2023). [CrossRef] [Green Version]
- Kovář, I. Family Coccinellidae Latreille, 1807. In Catalogue of Palaearctic Coleoptera; Löbl, I., Smetana, A., Eds.; Apollo Books: Stenstrup, Denmark, 2007; Volume 4, pp. 568–631. [Google Scholar]
- Mahmoud, A.K.; Yousif, N.E.; Satti, A.A. Effect of three aphid species on the life tables data of the coccinellid predator, Cheilomenes propinqua vicina (Mulsant). Environ. Nat. Resour. Int. J. 2016, 1, 49–55. [Google Scholar]
- Abdel-Salam, A.H.; El-Serafi, H.A.; Bayoumy, M.H.; Abdel-Hady, A.A. Effect of temperature and aphid-host plant variety on performance and thermal requirements of Coccinella undecimpunctata L. and Cheilomenes propinqua isis (Mulsant). J. Plant Prot. Pathol. 2018, 9, 375–380. [Google Scholar]
- Borkakati, R.N.; Venkatesh, M.R.; Saikia, D.K. Insect pests of Brinjal and their natural enemies. J. Entomol. Zool. Stud. 2019, 7, 932–937. [Google Scholar]
- Badary, H. Ecological aspects of Sassetia spp. (Coccidae: Coccoidae: Hemiptera) and thier natural enemies in Egypt. Egypt. Acad. J. Biol. Sci. A Entomol. 2011, 4, 163–174. [Google Scholar] [CrossRef]
- Bayoumy, M.H.; Michaud, J.P. Cannibalism in two subtropical lady beetles (Coleoptera: Coccinellidae) as a function of density, life stage, and food supply. J. Insect Behav. 2015, 28, 387–402. [Google Scholar] [CrossRef]
- Ghanim, A.A.; Abou-El-naga, A.A.; El-Serafi, H.A.; Jabbar, A.S. Studies on the population densities of certain insect pests attacking grapevine trees and their associated predators in Mansoura district. J. Plant Prot. Pathol. 2015, 6, 1089–1104. [Google Scholar] [CrossRef] [Green Version]
- Adly, D.; Fadl, H.A.; Mousa, S.F.M. Survey and seasonal abundance of mealybug species, their parasitoids and associated predators on guava trees in Egypt. Egypt. J. Biol. Pest Control 2016, 26, 657–664. [Google Scholar]
- Youssif, M.A.I. Coccinellid species and their insect parasitoids in pear orchards at El-Khattara district, Sharkia Governorate, Egypt. J. Entomol. Zool. Stud. 2019, 7, 780–790. [Google Scholar]
- Ramadan, M.M.; Hassan, M.A.; Afifi, M. Ecological studies on the Peach green aphid, Myzus persicae and its natural enemies. J. Plant Prot. Pathol. 2022, 13, 29–35. [Google Scholar] [CrossRef]
- Spodek, M.; Ben-Dov, Y.; Mondaca, L.; Protasov, A.; Erel, E.; Mendel, Z. The cotton mealybug, Phenacoccus solenopsis Tinsley (Hemiptera: Pseudococcidae) in Israel: Pest status, host plants and natural enemies. Phytoparasitica 2018, 46, 45–55. [Google Scholar] [CrossRef]
- Atuncha, H.; Ateka, E.; Amata, R.; Mwirichia, R.; Kasina, M.; Mbevi, B.; Wakoli, E. Evaluation of predation potential of coccinellids on cassava whiteflies. J. Entomol. Nematol. 2013, 5, 84–87. [Google Scholar] [CrossRef]
- Yarpuzlu, F.; Uygun, N. Effects of different temperatures on development and fecundity of a predatory beetle, Cheilomenes propinqua (Mulsant) (Coleoptera: Coccinellidae). Türkiye Biyol. Mücadele Derg. 2010, 1, 97–107. [Google Scholar]
- Nordey, T.; Boni, S.B.; Agbodzavu, M.K.; Mwashimaha, R.; Mlowe, N.; Ramasamy, S.; Deletre, E. Comparison of biological methods to control Aphis fabae Scopoli (Hemiptera: Aphididae) on kalanchoe crops in East Africa. Crop Prot. 2021, 142, 105520. [Google Scholar] [CrossRef]
- El-Heneidy, A.H.; El-Serafy, H.A.; Mohamed, N.E.; Elbanna, E.H.E. Biological characteristics of two coccinellid predators when reared on Aphis gossypii Glover and an artificial diet under laboratory condition. J. Plant Prot. Pathol. 2021, 12, 575–578. [Google Scholar] [CrossRef]
- Reznik, S.Y.; Ovchinnikov, A.N.; Bezman-Moseyko, O.S.; Samartsev, K.G.; Belyakova, N.A. Storage potential of the predatory ladybird Cheilomenes propinqua in relation to temperature, humidity, and factitious food. Insects 2022, 13, 613. [Google Scholar] [CrossRef]
- Ovchinnikov, A.; Reznik, S.; Bezman-Moseyko, O.; Belyakova, N. Walking activity of a predatory ladybird, Cheilomenes propinqua: Impacts of photoperiod, temperature, and starvation. BioControl 2022, 67, 513–522. [Google Scholar] [CrossRef]
- Messelink, G.J.; Bennison, J.; Alomar, O.; Ingegno, B.L.; Tavella, L.; Shipp, L.; Palevsky, E.; Wäckers, F.L. Approaches to conserving natural enemy populations in greenhouse crops: Current methods and future prospects. BioControl 2014, 59, 377–393. [Google Scholar] [CrossRef]
- Brenard, N.; Sluydts, V.; De Bruyn, L.; Leirs, H.; Moerkens, R. Food supplementation to optimize inoculative release of the predatory bug Macrolophus pygmaeus in sweet pepper. Entomol. Exp. Appl. 2018, 166, 574–582. [Google Scholar] [CrossRef] [Green Version]
- Brenard, N.; Sluydts, V.; Christianen, E.; Bosmans, L.; De Bruyn, L.; Moerkens, R.; Leirs, H. Biweekly supplementation with Artemia spp. cysts allows efficient population establishment by Macrolophus pygmaeus in sweet pepper. Entomol. Exp. Appl. 2019, 167, 406–414. [Google Scholar] [CrossRef]
- Kozlova, E.G.; Krasavina, L.P.; Orlova, G.S. Experience of biocontrol use in cucumber protection from the complex of greenhouse pests. Zashchita Karantin Rastenii 2019, 11, 39–42. (In Russian) [Google Scholar]
- Sade, A.; Steinberg, S.; Salinger-Bubnov, M.; Gilboa, E.; Klempert, G.; Roitman, N.; Grosman, A. Improved western flower thrips control through artemia-based early introduction of Orius laevigatus in commercial pepper greenhouses. IOBC/WPRS Bull 2019, 147, 39–46. [Google Scholar]
- Dader, B.; Colomer, I.; Adan, A.; Medina, P.; Vinuela, E. Compatibility of early natural enemy introductions in commercial pepper and tomato greenhouses with repeated pesticide applications. Insect Sci. 2020, 27, 1111–1124. [Google Scholar] [CrossRef] [Green Version]
- Kruidhof, H.M.; Elmer, W.H. Cultural Methods for Greenhouse Pest and Disease Management. In Integrated Pest and Disease Management in Greenhouse Crops; Gullino, M.L., Albajes, R., Nicot, P.C., Eds.; Springer: Cham, Switzerland, 2020; pp. 285–330. [Google Scholar]
- Benson, C.M.; Labbe, R.M. Exploring the role of supplemental foods for improved greenhouse biological control. Ann. Entomol. Soc. Am. 2021, 114, 302–321. [Google Scholar] [CrossRef]
- Woelke, J.B.; Bouw, M.; Cusumano, A.; Messelink, G.J. Lygus rugulipennis on chrysanthemum: Supplemental prey effects and an evaluation of trap plants. J. App. Entomol. 2023, 147, 157–166. [Google Scholar] [CrossRef]
- Agarwala, B.K.; Bhowmik, P.J. Effect of resource gradient on age and size at maturity and their influence on early-life fecundity in the predatory Asian lady beetle, Harmonia axyridis. Entomol. Exp. Appl. 2011, 141, 97–102. [Google Scholar] [CrossRef]
- Omkar; Sahu, J.; Kumar, G. Effect of prey quantity on reproductive and developmental attributes of a ladybird beetle, Anegleis cardoni. Int. J. Trop. Insect Sci. 2010, 30, 48–56. [Google Scholar] [CrossRef]
- Shah, M.A.; Khan, A.A. Qualitative and quantitative prey requirements of two aphidophagous coccinellids, Adalia tetraspilota and Hippodamia variegata. J. Insect Sci. 2014, 14, 72. [Google Scholar] [CrossRef] [Green Version]
- Ovchinnikova, A.A.; Ovchinnikov, A.N.; Dolgovskaya, M.Y.; Reznik, S.Y.; Belyakova, N.A. Trophic induction of diapause in native and invasive populations of Harmonia axyridis (Coleoptera: Coccinellidae). Eur. J. Entomol. 2016, 113, 469–475. [Google Scholar] [CrossRef] [Green Version]
- Sakurai, H.; Kawai, T.; Takeda, S. Physiological changes related to diapause of the lady beetle, Harmonia axyridis (Coleoptera: Coccinellidae). Appl. Entomol. Zool. 1992, 27, 479–487. [Google Scholar] [CrossRef] [Green Version]
- Osawa, N. The effect of prey availability on ovarian development and oosorption in the ladybird beetle Harmonia axyridis (Coleoptera: Coccinellidae). Eur. J. Entomol. 2005, 102, 503–511. Available online: https://www.eje.cz/pdfs/eje/2005/03/25.pdf (accessed on 21 June 2023). [CrossRef]
- Esquivel, J.F. Improved visualization of fat body cell conditions and abundance in the southern green stink bug (Hemiptera: Pentatomidae). J. Entomol. Sci. 2011, 46, 52–61. [Google Scholar] [CrossRef]
- Musolin, D.L. Surviving winter: Diapause syndrome in the southern green stink bug Nezara viridula in the laboratory, in the field, and under climate change conditions. Physiol. Entomol. 2012, 37, 309–322. [Google Scholar] [CrossRef]
- Raak-van den Berg, C.; Hemerik, L.; de Jong, P.W.; van Lenteren, J.C. Mode of overwintering of invasive Harmonia axyridis in the Netherlands. BioControl 2012, 57, 71–84. [Google Scholar] [CrossRef] [Green Version]
- Gao, Q.; Wei, B.-X.; Liu, W.; Wang, J.-L.; Zhou, X.-M.; Wang, X.-P. Differences in the development of internal reproductive organs, feeding amount and nutrient storage between pre-diapause and pre-reproductive Harmonia axyridis adults. Insects 2019, 10, 243. [Google Scholar] [CrossRef] [Green Version]
- Vargas, G.; Michaud, J.P.; Nechols, J.R. Larval food supply constrains female reproductive schedules in Hippodamia convergens (Coleoptera: Coccinellidae). Ann. Entomol. Soc. Am. 2012, 105, 832–839. [Google Scholar] [CrossRef] [Green Version]
- Barghi, N.; Ramirez-Lanzas, C. A high throughput method for egg size measurement in Drosophila. Sci. Rep. 2023, 13, 3791. [Google Scholar] [CrossRef]
- SYSTAT 10.2. Available online: https://systat.informer.com/10.2/ (accessed on 25 May 2023).
- Ives, P.M. Feeding and egg production of two species of coccinellids in the laboratory. Can. Entomol. 1981, 113, 999–1005. [Google Scholar] [CrossRef] [Green Version]
- Ferran, A.; Cruz De Boelpaepe, M.O.; Schanderl, H.; Larroque, M.M. Les aptitudes trophiques et reproductrices des femelles de Semiadalia undecimnotata [Col.: Coccinellidae]. Entomophaga 1984, 29, 157–170. [Google Scholar] [CrossRef]
- Xia, J.Y.; Van der Werf, W.; Rabbinge, R. Temperature and prey density on bionomics of Coccinella septempunctata (Coleoptera: Coccinellidae) feeding on Aphis gossypii (Homoptera: Aphididae) on cotton. Environ. Entomol. 1999, 28, 307–314. [Google Scholar] [CrossRef]
- Kajita, Y.; Evans, E.W. Ovarian dynamics and oosorption in two species of predatory lady beetles (Coleoptera: Coccinellidae). Physiol. Entomol. 2009, 34, 185–194. [Google Scholar] [CrossRef]
- Ponsonby, D.J.; Copland, M.J.W. Influence of host density and population structure on egg production in the coccidophagous ladybird, Chilocorus nigritus F. (Coleoptera: Coccinellidae). Agric. For. Entomol. 2007, 9, 287–296. [Google Scholar] [CrossRef]
- Atlıhan, R.; Güldal, H. Prey density-dependent feeding activity and life history of Scymnus subvillosus. Phytoparasitica 2009, 37, 35–41. [Google Scholar] [CrossRef]
- Agarwala, B.K.; Yasuda, H.; Sato, S. Life history response of a predatory ladybird, Harmonia axyridis (Pallas) (Coleoptera: Coccinellidae), to food stress. Appl. Entomol. Zool. 2008, 43, 183–189. [Google Scholar] [CrossRef] [Green Version]
- Bell, W.J.; Bohm, M.K. Oosorption in insects. Biol. Rev. 1975, 50, 373–396. [Google Scholar] [CrossRef]
- Kurihara, M. Oosorption in the Lady Beetle, Henosepilachna vigintioctomaculata (Coleoptera, Coccinellidae). In Recent Advances in Insect Embryology in Japan; Andō, H., Miya, K., Eds.; ISEBU Company: Tsukuba, Japan, 1985; pp. 25–44. [Google Scholar]
- Rondoni, G.; Ielo, F.; Ricci, C.; Conti, E. Behavioural and physiological responses to prey-related cues reflect higher competitiveness of invasive vs. native ladybirds. Sci. Rep. 2017, 7, 3716. [Google Scholar] [CrossRef]
- Ferrer, A.; Dixon, A.F.; Gibernau, M.; Hemptinne, J.L. Ovarian dynamics and specialisation in ladybirds. Ecol. Entomol. 2010, 35, 100–103. [Google Scholar] [CrossRef]
- Ferrer, A.; Corbani, A.C.; Dixon, A.F.; Hemptinne, J.L. Egg dumping by predatory insects. Physiol. Entomol. 2011, 36, 290–293. [Google Scholar] [CrossRef]
- Danks, H.V. Insect Dormancy: An Ecological Perspective; The Biological Survey of Canada: Ottawa, ON, Canada, 1987. [Google Scholar]
- Denlinger, D.L. Insect Diapause; Cambridge University Press: Cambridge, UK, 2022. [Google Scholar]
- Tauber, M.J.; Tauber, C.A.; Masaki, S. Seasonal Adaptations of Insects; Oxford University Press: New York, NY, USA, 1986. [Google Scholar]
- Inappen. Available online: https://inappen.com/products (accessed on 25 May 2023).
- Kasem, A.E.S. Improvement of the Methods of Rearing of a Predatory Bug Podisus maculiventris for Biological Protection of Plants. Ph.D. Thesis, All-Russia Institute of Plant Protection, St. Petersburg, Russia, 21 January 2021. Available online: http://vizrspb.ru/assets/docs/disser/KasemAE/KasemAS_disser.pdf (accessed on 25 May 2023). (In Russian).
- van Emden, H.F.; Wild, E.A. A fully defined artificial diet for Myzus persicae—The detailed technical manual. Entomol. Exp. Appl. 2020, 168, 582–586. [Google Scholar] [CrossRef]





{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Parameters and Sample Size | Factors | ||
|---|---|---|---|
| Diet (the Number of Aphids per Day) | The Age at Dissection (Days) | ||
| Survival (%, n = 782) 1 | c = 0.034, p = 0.076 | – 3 | |
| The onset of egg-laying (yes or no, n = 765) 1 | c = 0.016, p < 0.001 | c = 0.062, p < 0.001 | |
| For females that laid eggs | Pre-oviposition period (days, n = 584) 2 | c = −0.064, p = 0.321 | c = 3.531, p < 0.001 |
| Total fecundity (eggs, n = 584) 2 | c = 3.329, p < 0.001 | c = 9.543, p < 0.001 | |
| Egg-laying intensity (eggs/day, n = 584) 2 | c = 4.190, p < 0.001 | c = −2.722, p = 0.002 | |
| For all females | Total fecundity (eggs, n = 765) 2 | c = 9.0489, p < 0.001 | c = 13.247, p < 0.001 |
| Egg-laying intensity (eggs/day, n = 765) 2 | c = 4.859, p < 0.001 | c = 4.379, p < 0.001 | |
| Reproduction Parameter | Diet (the Number of Aphids Offered Daily) | |||
|---|---|---|---|---|
| 0 | 2 | 10 | 50 | |
| Egg-laying females (%) | 58.6 ± 4.4 a | 67.4 ± 4.0 b | 81.43 ± 3.9 c | 85.6 ± 4.9 c |
| The mean pre-oviposition period (days) | 8.7 ± 0.3 a | 9.4 ± 0.4 a | 8.4 ± 0.2 a | 7.9 ± 0.2 a |
| The mean total fecundity calculated for egg-laying females (eggs/female) | 71.1 ± 7.2 a | 47.5 ± 5.1 a | 123.0 ± 9.1 b | 271.1 ± 17.8 c |
| Oviposition intensity calculated for egg-laying females (eggs/female/day) | 7.0 ± 0.6 a | 5.5 ± 0.4 a | 11.4 ± 0.6 b | 22.9 ± 1.0 c |
| The mean total fecundity calculated for all females (eggs/female) | 44.5 ± 5.1 a | 33.9 ± 4.0 a | 102.6 ± 8.3 b | 237.8 ± 16.9 c |
| Oviposition intensity calculated for all females (eggs/female/day) | 4.4 ± 0.4 a | 3.9 ± 0.4 a | 9.5 ± 0.6 b | 20.1 ± 1.0 c |
| Reproductively active females (%) | 51.9 ± 3.9 a | 67.9 ± 4.2 b | 85.9 ± 3.0 c | 93.1 ± 3.1 d |
| Females in the intermediate state (%) | 9.1 ± 2.2 ab | 12.9. ± 2.9 a | 7.1 ± 1.8 ab | 4.6 ± 1.6 b |
| Diapausing females (%) | 35.6 ± 3.3 a | 15.9 ± 2.6 b | 6.4 ± 1.9 c | 2.2 ± 1.0 c |
| Underdeveloped females (%) | 3.3 ± 1.5 a | 3.2 ± 1.5 a | 0.6 ± 0.6 a | 0.0 ± 0.0 a |
| Females with egg resorption (%) | 23.2 ± 4.1 a | 19.3 ± 3.2 a | 6.1 ± 1.9 b | 1.1 ± 0.8 b |
| The size of the largest follicle (mm3) | 0.117 ± 0.004 a | 0.125 ± 0.003 a | 0.147 ± 0.002 ab | 0.160 ± 0.002 b |
| Females with moderately or fully developed fat body (%) | 46.8 ± 3.7 a | 25.5 ± 2.3 b | 9.0 ± 2.4 c | 3.0 ± 1.1 d |
| Parameters | Factors | ||
|---|---|---|---|
| Diet (the Number of Aphids per Day) | The Age at Dissection (Days) | ||
| Reproductive state at dissection (n = 765) | Reproductively active 1 | c = 0.026, p < 0.001 | c = 0.038, p < 0.001 |
| Intermediate 1 | c = −0.011, p = 0.007 | c = −0.032, p = 0.001 | |
| Diapausing 1 | c = −0.028, p < 0.001 | c = −0.020, p = 0.018 | |
| Underdeveloped 1 | c = −0.077, p = 0.059 | c = −0.056, p = 0.011 | |
| Resorption 1 | c = −0.032, p < 0.001 | c = 0.041, p < 0.001 | |
| Follicle size (mm3) 2 | c = 3.522, p < 0.001 | c = 0.597, p = 0.523 | |
| Fat body development (n = 765) 1 | c = −0.031, p < 0.001 | c = −0.014, p = 0.070 | |
| Correlation between Degrees of Development of Ovaries and Fat Body | Diet (the Number of Aphids Provided Daily) | |||
|---|---|---|---|---|
| 0 | 2 | 10 | 50 | |
| Spearman rank correlation coefficient, ρ | ρ = −0.763 | ρ = −0.688 | ρ = −0.731 | ρ = −0.634 |
| Chi-square χ2, sample size n, and significance p | χ2 = 148.4, n = 192, p < 0.001 | χ2 = 110.5, n = 185, p < 0.001 | χ2 = 175.7, n = 193, p < 0.001 | χ2 = 188.7, n = 195, p < 0.001 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Ovchinnikov, A.N.; Ovchinnikova, A.A.; Reznik, S.Y.; Belyakova, N.A. Signal and Nutritional Effects of Mixed Diets on Reproduction of a Predatory Ladybird, Cheilomenes propinqua. Insects 2023, 14, 587. https://doi.org/10.3390/insects14070587
Ovchinnikov AN, Ovchinnikova AA, Reznik SY, Belyakova NA. Signal and Nutritional Effects of Mixed Diets on Reproduction of a Predatory Ladybird, Cheilomenes propinqua. Insects. 2023; 14(7):587. https://doi.org/10.3390/insects14070587
Chicago/Turabian StyleOvchinnikov, Andrey N., Antonina A. Ovchinnikova, Sergey Y. Reznik, and Natalia A. Belyakova. 2023. "Signal and Nutritional Effects of Mixed Diets on Reproduction of a Predatory Ladybird, Cheilomenes propinqua" Insects 14, no. 7: 587. https://doi.org/10.3390/insects14070587