
How Rearing Systems for Various Species of Flies Benefit Humanity


Abstract
:Simple Summary
Abstract
1. Introduction
2. The Historical Importance of Fly Rearing as a Foundation for Insect Rearing
3. Uses and Rearing Diets of Selected Fly Species
4. Rearing Flies for Animal Feed and to Feed the Growing Human Population
Environmental Concerns of Industrial Insect Rearing for Food and Feed
5. Rearing for Pollination Services and Pest Control
5.1. Diets for Predaceous Syrphids
5.2. Tachinid Rearing
6. Rearing of Medicinal Maggots
6.1. Medicinal Maggots for Wound Therapy
6.2. Rearing Practices for Fly Colony Maintenance in Maggot Therapy
6.3. Benefits and Risks of Maggot Therapy
6.4. Mechanisms of Action by Maggots
6.5. Commercial Suppliers and Medical Procedures
- International Biotherapy Society (http://biotherapysociety.org/ accessed on 11 June 2023);
- BioMonde (https://biomonde.com accessed on 11 June 2023) in Germany (but they also operate in Great Britain);
- Monarch Labs (https://www.monarchlabs.com/ accessed on 11 June 2023).
7. Rearing of Forensic Flies
8. Laboratory-Reared Drosophila melanogaster as a Model for Great Scientific Discoveries
9. Key Topics of Research in Fly Rearing
10. Conclusions
Author Contributions
Funding
Data Availability Statement
Acknowledgments
Conflicts of Interest
Appendix A

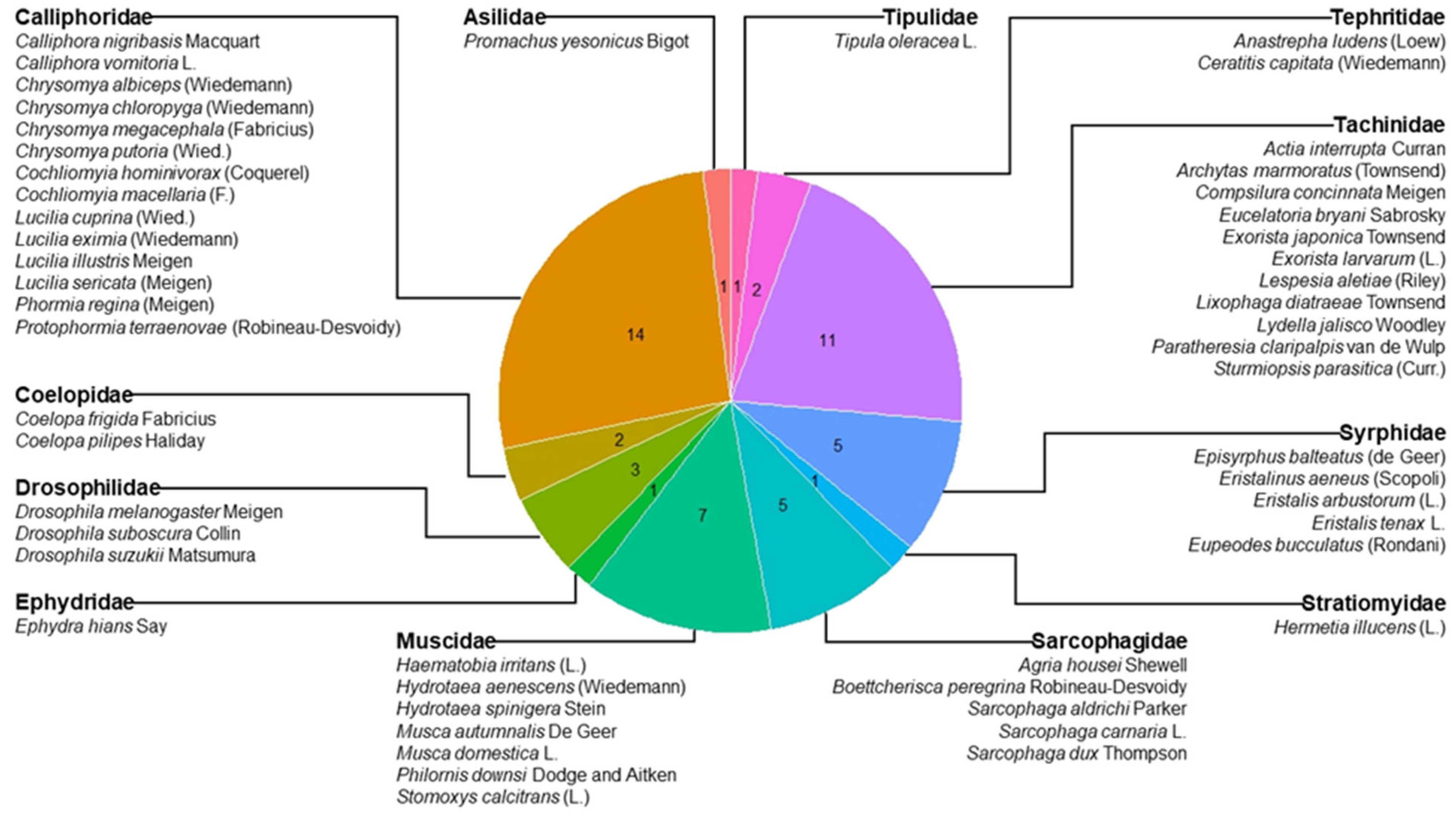{kind=link}
| Family and Species | Uses | Larval Diets | Adult Diets | References |
|---|---|---|---|---|
| Asilidae | ||||
| Promachus yesonicus Bigot | Biological control | White grubs as larval prey | Leafhoppers and moths | [99] |
| Calliphoridae | ||||
| Calliphora nigribasis Macquart | To estimate the postmortem interval in forensic research | Beef liver | Not indicated | [130] |
| Calliphora vomitoria L. | Laboratory experiments | Bovine meat treated with trypsin, salt solution, rice starch | Not indicated | [27] |
| Chrysomya albiceps (Wiedemann) | Forensic research | Beef liver | Adults were not reared | [130] |
| Chrysomya chloropyga (Wiedemann) | Waste bioconversion, nutrient recovery | Carrion | Not indicated | [22] |
| Chrysomya megacephala (Fabricius) | Forensic research, Pollination services in mango crops, Waste bioconversion, Source of biodiesel | Beef liver; Minced beef; Semi-synthetic diets with sardine, rumen, or chicken eggs, water, milk powder, yeast, casein, nipagin, and agar; A non-meat-based artificial diet with soya flour, milk powder, and egg as protein sources, sodium benzoate, and a gelling agent; Carrion, manure, kitchen waste (meat and vegetables), waste streams, and meat processing waste; Restaurant garbage | 1% sugar solution | [22,130,131,179,180] |
| Chrysomya putoria (Wied.) | Forensic research; Waste bioconversion | Minced beef; semi-synthetic diets with sardine, rumen, or chicken eggs, water, milk powder, yeast, casein, nipagin, and agar; Carrion, meat- and manure-based waste streams | Not indicated | [22,131] |
| Cochliomyia hominivorax (Coquerel) | Application of the Sterile Insect Technique | Various types of artificial diets: lean-meat-diet, liquid diet, gelled diet, cellulose-fiber-based diet | A mixture of ground horse meat and honey. Sucrose, dried egg powder or milk powder | [181] |
| Cochliomyia macellaria (F.) | Forensic research; Laboratory experiments | Beef liver; Dried bovine blood, spray-dried poultry eggs, dried milk substitute, polyacrylate gel | Sugar solution | [62,137] |
| Lucilia cuprina (Wied.) | Maggot therapy; Research on aldrin epoxidase activity | Artificial diet; Fish liver; artificial diet made of yeast and powdered milk in an agar base | Artificial diet | [62,182] |
| Lucilia eximia (Wiedemann) | Forensic research | Beef liver | Adults were not reared | [130] |
| Lucilia illustris Meigen | Maggot therapy | Beef liver, sterile raw chicken egg | Not indicated | [113] |
| Lucilia sericata (Meigen) | Maggot therapy; Organic waste treatment; Waste bioconversion; Laboratory experiments; Evidence in criminal investigations in forensic research | Black pudding, horse blood agar, meat processing waste; Artificial diet made of milk powder, dried yeast, wheat germ, agar, propionic acid, and water; Poultry waste, fish waste, carrion, meat, manure; Carrion, manure, meat processing waste, fish, and poultry; Artificial diet, including milk powder, dried yeast, wheat germ, cellulose powder, agar powder, propionic acid, and water; Chicken liver, ground beef with fat | Layers of red meat; Artificial diet with the same ingredients of larval diet but without propionic acid; Sugar and water ad libitum, blood | [22,61,63,116,132,183] |
| Phormia regina (Meigen) | Evidence in criminal investigations in forensic research; Research on the nutritional ecology of saprophagous calliphorids | Chicken liver, ground beef with fat; Lean pork muscle; Artificial meridic diet with casein, yeast, sucrose, and cellulose; Lamb’s liver | Sugar and water ad libitum, chicken blood and liver | [132,133,184] |
| Protophormia terraenovae (Robineau-Desvoidy) | Animal feed ingredient | Meat waste | Not indicated | [185] |
| Coelopidae | ||||
| Coelopa frigida Fabricius | Research as an alternative ingredient for animal feed | Seaweed and algae | Not indicated | [22] |
| Coelopa pilipes Haliday | Research as an alternative ingredient for animal feed | Seaweed and algae | Not indicated | [22] |
| Drosophilidae | ||||
| Drosophila melanogaster Meigen | Model organism in biological and medical research | A variety of synthetic media including chemically defined (i.e., holidic) diet consisting of a gelling agent, sugar, amino acids, metal ions, cholesterol, vitamins, preservatives, and other nutrients | Same as larvae | [186] |
| Drosophila suboscura Collin | Research on evolutionary biology | Artificial medium composed of agar, corn meal, dead brewer’s yeast, charcoal coloring, and nipagin | Same as larvae | [187] |
| Drosophila suzukii Matsumura | Research for pest control applying the Sterile Insect Technique | Various artificial diet formulations containing yeast and sucrose | Same as larvae | [188] |
| Ephydridae | ||||
| Ephydra hians Say | Environmental research | Algae, fish food | Not indicated | [189] |
| Muscidae | ||||
| Haematobia irritans (L.) | Research on pest control | On-animal: feces from the host. Off-animal: fresh bovine feces from animals known to not have been treated with chemicals that could inhibit development; fresh feces mixed with peanut hull pellets; a mixture of vermiculite, wheat bran and a commercial animal feed combined with fresh feces | On-animal: blood directly from the host. Off-animal: citrated bovine blood | [190] |
| Hydrotaea aenescens (Wiedemann) | Forensic research | A mixture of horse and dog manure. Artificial diet (wheat bran, alfalfa meal, yeast suspension, water, and malt sugar) | 50:50 mixture of table sugar and powdered milk, water | [191] |
| Hydrotaea spinigera Stein | Forensic research | Slightly decomposed lean pork on top of humus soil | A mixture of milk powder and sugar, and freshwater | [192] |
| Musca autumnalis De Geer | Research for use in organic waste treatment/bioconversion | Cattle manure | Not indicated | [22] |
| Musca domestica L. | As a host to produce parasitoids for biocontrol; Organic waste treatment/bioconversion. Recovery of nutrients; Animal feed | Various diet formulations containing rice bran, powdered milk, dry yeast, fish meal, soybean meal, chicken feed, and water; Poultry, swine and cattle manure. Bran and blood; Millet bran, wheat bran, and cow dung (70% humidity adjusted by adding tap water to dry substrates); Poultry, pig, and cow manure, garbage, food scrapes, concentrated pig slurry; Maze bran | Water and a mixture of sugar and powdered milk; A mixture of powdered milk and granulated sugar, water; A liquid formula, including water, sugar, milk, and other ingredients not reported; Honey, a mixture of 50% dry milk and 50% sugar. Fruit: banana, pineapple and papaya | [22,63,67,193,194] |
| Philornis downsi Dodge and Aitken | Research for pest control | Chicken blood, or chicken blood added with hydrolyzed protein, milk powder, or brewer’s yeast | A diet consisting of water, ripe papaya, sugar, hydrolyzed protein, and milk powder | [195] |
| Stomoxys calcitrans (L.) | To produce healthy individuals for experimentation. Research on pest control; Conduct research to establish alternatives of parasite control methods; | A mixture of wheat bran, wood shavings, fish meal, ammonium bicarbonate and water; Alfalfa hay and water; A mixture of crushed dry dog food, beer yeast and blood meal, mixed with vermiculite | Bovine blood with sodium citrate tribasic dihydrate and water; Citrated equine blood, plus a mixture of brown sugar mixed with tap water; Bovine blood | [196,197,198] |
| Sarcophagidae | ||||
| Agria housei Shewell | Parasitoid–Biocontrol against lepidopteran pests | Media based on salmon and fresh liver | Not indicated | [64] |
| Boettcherisca peregrina Robineau-Desvoidy | Waste treatment. Biodiesel production | Swine manure, fermentation residues, degreased food waste, fish rendering waste and poultry processing waste | Not indicated | [22] |
| Sarcophaga aldrichi Parker | Parasitoid–Biocontrol against lepidopteran pests | Artificial diet—Pork liver and fish | Not indicated | [64] |
| Sarcophaga carnaria L. | Organic waste treatment/bioconversion; Food additives in the fish industry; | Fish and poultry rendering waste; Poultry and fish wastes; Decaying meat, fish waste | Granular sugar and water | [22,63,199] |
| Sarcophaga dux Thomson | Waste treatment | Carrion, animal manure | Not indicated | [22] |
| Stratiomyidae | ||||
| Hermetia illucens (L.) | Bioactive compounds for cosmetic applications; Bioconversion of sewage sludge; Animal feed; Source of provitamin A carotenoids; Organic waste treatment/bioconversion; Biodisel production | Various kinds of decaying organic matter including vegetable waste, restaurant waste, poultry manure, fruit waste, seaweed, among others; Gainesville diet (wheat bran, alfalfa meal, corn meal, tap water) mixed with municipal sewage sludge; A variety of substrates including cattle, chicken and swine manure; restaurant waste, beet molasses, potato steam peelings, spent grains and beer yeast, bread and cookie remains, fruit and vegetables, artificial diets; White mushrooms, orange carrots, Cinderella pumpkins, sweet potatoes and clementines; Manure, plant waste, sugar beet pulp, meat and bone meal, distillers grains, swine manure, cattle manure, poultry manure, cereal processing waste, citric waste, brewery waste, olive waste; Poultry, pig and dairy manure, rotting plant material, food scrapes, municipal organic waste, market waste, cattle feces, urine slurry; Livestock manure, fruit and vegetables, fermented coconut waste, brewer’s spent grain | Adults do not require food for mating and oviposition, but providing water and sugar or honey can increase longevity | [22,63,69,84,200,201,202] |
| Syrphidae | ||||
| Episyrphus balteatus (de Geer) | Potential biocontrol agents of aphid pests | Artificial diet with drone honeybee brood powder combined with autolyzed brewer’s yeast and sucrose | Bee-collected pollen and honey solution | [103] |
| Eristalinus aeneus (Scopoli) | Candidate for pollination services | Brewery spent grain and soaked oat grains | Fresh pollen, sugar, honey, and water | [203] |
| Eristalis arbustorum (L.) | Research for use as biomarkers of pollutants | Rabbit droppings mixed with water alone or together with vitamin C and yeast | Flowers and honey | [204] |
| Eristalis tenax L. | Crop pollination, biodegradation of organic animal waste, a model for studying motion vision and flight dynamics, a model for studying investment in microbial immunity | Artificial diet based on cereal grains mixed with water; Fresh rabbit feces mixed with water, cow dung | Water, fresh pollen, and sugar; Bee pollen and honey, water | [97] |
| Eupeodes bucculatus (Rondani) | Potential biocontrol agents of aphid pests | Artificial diet with drone honeybee, brood powder | Bee-collected pollen and honey solution | [103] |
| Tachinidae | ||||
| Actia interrupta Curran | Laboratory experimentation as a potential biocontrol of insect pests | Host larvae of Choristoneura fumiferana (Clemens) | Not indicated | [205] |
| Archytas marmoratus (Townsend) | Biological control of lepidopteran pests | Galleria mellonella larval hosts reared on different artificial diets | Not indicated | [105] |
| Compsilura concinnata Meigen | Biological control of lepidopteran pests | Lymantria dispar L. larval host | Not indicated | [64] |
| Eucelatoria bryani Sabrosky | Parasitoid for biological control of Heliotis spp. and Helicoverpa spp. | Artificial medium containing soy flour, free amino acids, chicken egg yolk | Not indicated | [206] |
| Exorista japonica Townsend | Parasitoid for biological control of lepidopteran pests | Larvae of the common armyworm Mythimna separata Walker reared on artificial diet | Not indicated | [178] |
| Exorista larvarum (L.) | Parasitoid for biological control of lepidopteran pests | Artificial medium (skimmed milk, chicken egg yolk, yeast extract, sucrose, and gentamicin) alone or enriched with H. illucens hemolymph; Last instar larvae of the greater wax moth (Galleria mellonella (L.)) | Sugar cubes and cotton balls soaked in a honey and water solution; Pollen | [66,207] |
| Lespesia aletiae (Riley) | Biological control of lepidopteran pests | Sixth instar fall armyworm (Spodoptera frugiperda (J.E. Smith)) larval hosts | Water, an aqueous sucrose solution and hydrolyzed brewer’s yeast | [208] |
| Lixophaga diatraeae Townsend | Biological control of lepidopteran pests | Larval host (Diatraea spp., G. mellonella). Artificial medium gelled with agarose containing organic acids, inorganic salts, amino acids, B and fat-soluble vitamins, gelatin, protein hydrolysates, glycogen, adenosine triphosphate, lecithin, corn oil, and cholesterol | Not indicated | [64] |
| Lydella jalisco Woodley | Biological control of Eoreuma loftini (Dyar) | Larvae of the rice borer E. loftini | Water in cotton balls, and with a diet consisting of fructose syrup, agar, spray dried whole egg, and distilled water | [209] |
| Paratheresia claripalpis van de Wulp | Biological control of sugarcane borers (Diatraea spp.) | Artificial diet | Not indicated | [64] |
| Sturmiopsis parasitica (Curr.) | Biological control of lepidopteran stem borer pests | Larval hosts of Chilo auricilius Dudgn., C. partellus, C. infuscatellus Sn., C. sacchariphagus indicus (Kapur), Sesamia inferens (Wlk.) | Cotton swabs soaked in a honey solution and sugar cubes | [210] |
| Tephritidae | ||||
| Anastrepha ludens (Loew) | Application of the Sterile Insect Technique; As a host to produce parasitoids for biocontrol; To assess the biosafety of potential probiotic bacteria | Artificial diet with maize cob fractions, maize flour, dried torula yeast, sugar, citric acid, sodium benzoate, methyl-paraben, guar gum, and water; Artificial diet with dried yeast, natural wheat germ, refined sugar, corn cob fractions, sodium benzoate, hydrochloric acid, viterra plus capsules, water | A 3:1 sugar: protein mixture and water | [211,212,213] |
| Ceratitis capitata (Wiedemann) | Application of the Sterile Insect Technique; Larva is used to produce various feed and food ingredients | Artificial diet including dry ground maize plant fractions, wheat bran, sugar, inactive dry yeast, citric acid, methyl-paraben, sodium benzoate, and water; Meridic diet formulation (i.e., most of the ingredients are chemically defined) | A 3:1 sugar: protein hydrolysate mixture and water; Chemically defined (i.e., holidic) diet formulation | [86,214,215] |
| Tipulidae | ||||
| Tipula oleracea L. | Source of larvae for bioassays on pest control | Dried powdered grass, lettuce leaves | Not indicated | [216] |
References
- Vockeroth, J. Introducing the ubiquitous Diptera. Biodiversity 2002, 3, 3–5. [Google Scholar]
- Labandeira, C.C.; Yeates, D.K.; Wiegmann, B.M. Fossil history and evolutionary ecology of Diptera and their associations with plants. In The Evolutionary Biology of Flies; Yeates, D.K., Wiegmann, B.M., Eds.; Columbia University Press: New York, NY, USA, 2005; pp. 217–273. [Google Scholar]
- Yeates, D.K.; Wiegmann, B.M.; Courtney, G.W.; Meier, R.; Lambkin, C.; Pape, T. Phylogeny and systematics of Diptera: Two decades of progress and prospects. Zootaxa 2007, 1668, 565–590. [Google Scholar] [CrossRef]
- Stork, N.E. How many species of insects and other terrestrial arthropods are there on Earth. Annu. Rev. Entomol. 2018, 63, 31–45. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Mlynarek, J.J. Diptera pollinators. Curr. Biol. 2022, 32, R305–R306. [Google Scholar] [CrossRef] [PubMed]
- McGavin, G.C.; Sorkin, L.N.; Gorton, S. Smithsonian Handbooks: Insects Spiders and Other Terrestrial Arthropods; Covent Garden Books: New York, NY, USA, 2002. [Google Scholar]
- Menees, J.H. The skeletal elements of the gnathocephalon and its appendages in the larvae of higher Diptera. Ann. Entomol. Soc. Am. 1962, 55, 607–616. [Google Scholar] [CrossRef]
- Foley, D.H. Flies as vectors of disease. Biodiversity 2002, 3, 18–20. [Google Scholar]
- Lockwood, J.A. Insects as weapons of war, terror, and torture. Annu. Rev. Entomol. 2012, 57, 205–227. [Google Scholar] [CrossRef]
- Gerhardt, R.R.; Hribar, L.J. Flies (Diptera). In Medical and Veterinary Entomology, 3rd ed.; Mullen, G.R., Durden, L.A., Eds.; Academic Press: London, UK, 2019; pp. 171–191. [Google Scholar]
- Alsan, M. The effect of the tsetse fly on African development. Am. Econ. Rev. 2015, 105, 382–410. [Google Scholar] [CrossRef] [Green Version]
- Aluja, M.; Mangan, R.L. Fruit fly (Diptera: Tephritidae) host status determination: Critical conceptual, methodological, and regulatory considerations. Annu. Rev. Entomol. 2008, 53, 473–502. [Google Scholar] [CrossRef]
- Savage, J. Cleaning up the world: Dipteran decomposers. Biodiversity 2002, 3, 12–15. [Google Scholar]
- Ssymank, A.; Kearns, C.A.; Pape, T.; Thompson, F.C. Pollinating flies (Diptera): A major contribution to plant diversity and agricultural production. Biodiversity 2008, 9, 86–89. [Google Scholar] [CrossRef]
- Doyle, T.; Hawkes, W.L.S.; Massy, R.; Powney, G.D.; Menz, M.H.M.; Wotton, K.R. Pollination by hoverflies in the Anthropocene. Proc. R. Soc. B 2020, 287, 20200508. [Google Scholar] [CrossRef]
- Sherman, R.A. Maggot therapy takes us back to the future of wound care: New and improved maggot therapy for the 21st century. J. Diabetes Sci. Technol. 2009, 3, 336–344. [Google Scholar] [CrossRef] [Green Version]
- Kruglikova, A.A.; Chernysh, S.I. Surgical maggots and the history of their medical use. Entomol. Rev. 2013, 93, 667–674. [Google Scholar] [CrossRef]
- Hernigou, P.; Hernigou, J.; Scarlat, M. The dark age of medieval surgery in France in the first part of Middle Age (500–1000): Royal touch, wound suckers, bizarre medieval surgery, monk surgeons, Saint Healers, but foundation of the oldest worldwide still-operating hospital. Int. Orthop. 2021, 45, 1633–1644. [Google Scholar] [CrossRef]
- Orkusz, A. Edible insects versus meat—Nutritional comparison: Knowledge of their composition is the key to good health. Nutrients 2021, 13, 1207. [Google Scholar] [CrossRef]
- Bosch, G.; Oonincx, D.G.A.B.; Jordan, H.R.; Zhang, J.; van Loon, J.J.A.; van Huis, A.; Tomberlin, J.K. Standardisation of quantitative resource conversion studies with black soldier fly larvae. J. Insects Food Feed 2020, 6, 95–109. [Google Scholar] [CrossRef]
- Cadinu, L.A.; Barra, P.; Torre, F.; Delogu, F.; Madau, F.A. Insect rearing: Potential, challenges, and circularity. Sustainability 2020, 12, 4567. [Google Scholar] [CrossRef]
- Parry, N.J.; Pieterse, E.; Weldon, C.W. The case for a wider range of flies for use in waste bioconversion. J. Insects Food Feed 2021, 7, 1161–1175. [Google Scholar] [CrossRef]
- Singh, P. Artificial Diets for Insects, Mites, and Spiders; Springer: New York, NY, USA, 1977. [Google Scholar]
- Cohen, A.C. Insect Diets: Science and Technology, 2nd ed.; CRC Press: Boca Raton, FL, USA, 2015. [Google Scholar]
- Cohen, A.C. Design, Operation, and Control of Insect Rearing Systems; CRC Press: Boca Raton, FL, USA, 2021. [Google Scholar]
- Bogdanow, E.A. About the breeding of larvae in usual flesh flies (Calliphora vomitoria) in sterilized nutrients. Arch. Eur. J. Physiol. 1906, 113, 97–105. [Google Scholar] [CrossRef] [Green Version]
- Bogdanow, E.A. About the dependence of the growth of larvae of bacteria and enzymes and about variability and inheritance in the flesh flies. Arch. Anat. Physiol. 1908, 173–200. [Google Scholar]
- Delcourt, A.; Guyenot, E. The possibility of studying certain Diptera in a defined environment. CR Hebd. Acad. Sci. 1910, 151, 255–257. [Google Scholar]
- Guyenot, E. A biological study of a Drosophila ampelophila Low fly I—The possibility of an aseptic life for an individual and the line. CR Seances Soc. Biol. Fil. 1913, 74, 97–99. [Google Scholar]
- Guyenot, E. Biological study in a fly, Drosophila ampeltophila Low II—The role of yeast in food. CR Seances Soc. Biol. Fil. 1913, 74, 178–180. [Google Scholar]
- Guyenot, E. Biological studies on a fly, Drosophila ampelophila Low V—Nutrition of adults and fertility. CR Seances Soc. Biol. Fil. 1913, 74, 332–334. [Google Scholar]
- Guyenot, E. Biological studies on a fly Drosophila ampelophila Low. Need to create a definite feeding environment. CR Seances Soc. Biol. Fil. 1914, 76, 483–485. [Google Scholar]
- Guyenot, E. Primary trials of determination of an artificial nutritive environment for the raising of a fly, Drosophila ampelophila Low. CR Seances Soc. Biol. Fil. 1914, 76, 548–550. [Google Scholar]
- Baumberger, J.P. The food of Drosophila melanogaster Meigen. Proc. Natl. Acad. Sci. USA 1917, 3, 122–126. [Google Scholar] [CrossRef] [Green Version]
- Baumberger, J.P. A nutritional study of insects, with special reference to microorganisms and their substrata. J. Exp. Zool. 1919, 28, 1–81. [Google Scholar] [CrossRef] [Green Version]
- Morgan, T.H.; Sturtevant, A.N.H.; Muller, H.J.; Bridges, C.B. The Mechanism of Mendelian Heredity; Henry Holt and Company: New York, NY, USA, 1915. [Google Scholar]
- Bridges, C.B. Apparatus and methods for Drosophila culture. Am. Nat. 1932, 66, 250–273. [Google Scholar] [CrossRef]
- Bridges, C.B.; Darby, H.H. Culture media for Drosophila and the pH of media. Am. Nat. 1933, 67, 437–472. [Google Scholar] [CrossRef]
- Loeb, J. The simplest constituents required for growth and the completion of the life cycle in an insect (Drosophila). Science 1915, 41, 169–170. [Google Scholar] [CrossRef] [Green Version]
- Baumberger, J.P. Solid media for rearing Drosophila. Am. Nat. 1917, 51, 447–448. [Google Scholar] [CrossRef] [Green Version]
- Baumberger, J.P.; Glasser, R.W. The rearing of Drosophila ampelophila Loew on solid media. Science 1917, 45, 21–22. [Google Scholar] [CrossRef]
- Loeb, J.; Northrop, J.H. On the influence of food and temperature on the duration of life. J. Biol. Chem. 1917, 32, 103–121. [Google Scholar] [CrossRef]
- Michelbacher, A.E.; Hoskins, W.M.; Herms, W.B. The nutrition of flesh fly larvae, Lucilia sericata (Meig.). J. Exp. Zool. 1932, 64, 109–131. [Google Scholar] [CrossRef]
- Marucci, P.E.; Clancy, D.W. The artificial culture of fruit flies and their parasites. Proc. Hawaii Entomol. Soc. 1950, 14, 163–166. [Google Scholar]
- Moore, I. A method for artificially culturing the olive fly (Dacus oleae Gmel.) under aseptic conditions. J. Agric. Res. Stat. Reh. Ktav. 1959, 9, 295–296. [Google Scholar]
- Hagen, K.S.; Santas, L.; Tsecouras, A. A Technique of Culturing the Olive Fly, Dacus oleae Gmel, on Synthetic Media Under Xenic Conditions. In Proceedings of the Symposium on the Use and Application of Radioisotopes and Radiation in the Control of Plant and Animal Insect Pests, Athens, Greece, 22–26 April 1963. [Google Scholar]
- Santas, L.A. On a new synthetic nutritive substrate for rearing larvae of Dacus oleae Gmel. (Diptera-Trypitidae). Bull. Agr. Bank. Greece 1965, 144, 8. [Google Scholar]
- Economopoulos, A.P.; Tzanakakis, M.E. Egg yolk and olive juice as supplements to the yeast hydrolysate-sucrose diet for adults of Dacus oleae. Life Sci. 1967, 6, 2409–2416. [Google Scholar] [CrossRef]
- Orphanidis, P.S.; Petsikon, N.; Petsekos, P.G. Eléments sommaires concernant un élevage expérimental du Dacus de l’olive sur substrat artificial. In Proceedings of the 8th FAO Ad Hoc Conference on the Control of Olive Pests and Diseases, Athens, Greece, 8–12 May 1969. [Google Scholar]
- Baumhover, A.H.; Graham, A.J.; Bitter, B.A.; Hopkins, D.E.; New, W.D.; Dudley, F.H.; Bushland, R.C. Screw-worm control through releases of sterilized flies. J. Econ. Entomol. 1955, 48, 462–466. [Google Scholar] [CrossRef]
- Bushland, R.C.; Lindquist, A.W.; Knipling, E.F. Eradication of screw-worms through release of sterilized males. Science 1955, 122, 287–288. [Google Scholar] [CrossRef] [PubMed]
- Melvin, R.; Bushland, R.C. A method of rearing Chochliomyia Americana C. and P. on artificial media. U.S. Bur. Entomol. Plant Quar. 1936, 88, 2. [Google Scholar]
- Melvin, R.; Bushland, R.C. The nutritional requirements of screwworm larvae. J. Econ. Entomol. 1940, 33, 850–852. [Google Scholar] [CrossRef]
- Boller, E.F.; Chambers, D.L. Quality control: An idea book for fruit fly workers. IOBC/WPRS Bull. 1977, 5, 162. [Google Scholar]
- Leppla, N.C.; Huettel, M.D.; Chambers, D.L.; Ashley, T.R.; Miyashita, D.H.; Wong, T.T.Y.; Harris, E.J. Strategies for colonization and maintenance of the Mediterranean fruit fly. Entomol. Exp. Appl. 1983, 33, 89–96. [Google Scholar] [CrossRef]
- Cáceres, C.; Hendrichs, J.; Vreysen, M.J. Development and improvement of rearing techniques for fruit flies (Diptera: Tephritidae) of economic importance. Int. J. Trop. Insect Sci. 2014, 34, S1–S12. [Google Scholar] [CrossRef]
- Bakri, A.; Metha, K.; Lance, D.R. Sterilizing insects with ionizing radiation. In Sterile Insect Technique. Principles and Practice in Area-Wide Integrated Pest Management, 2nd ed.; Dyck, V.A., Hendrichs, J., Robinson, A.S., Eds.; CRC Press: Boca Raton, FL, USA, 2021; pp. 355–398. [Google Scholar]
- Parker, A.; Mamai, W.; Maiga, H. Mass-rearing for the sterile insect technique. In Sterile Insect Technique. Principles and Practice in Area-Wide Integrated Pest Management, 2nd ed.; Dyck, V.A., Hendrichs, J., Robinson, A.S., Eds.; CRC Press: Boca Raton, FL, USA, 2021; pp. 283–316. [Google Scholar]
- Pérez-Staples, D.; Díaz-Fleischer, F.; Montoya, P. The sterile insect technique: Success and perspectives in the Neotropics. Neotrop. Entomol. 2021, 50, 172–185. [Google Scholar] [CrossRef]
- Vreysen, M.J.B.; Abd-Alla, A.M.M.; Bourtzis, K.; Bouyer, J.; Caceres, C.; de Beer, C.; Oliveira Carvalho, D.; Maiga, H.; Mamai, W.; Nikolouli, K.; et al. The insect pest control laboratory of the joint FAO/IAEA programme: Ten years (2010–2020) of research and development, achievements and challenges in support of the Sterile Insect Technique. Insects 2021, 12, 346. [Google Scholar] [CrossRef]
- Zhang, B.; Numata, H.; Mitsui, H.; Goto, S.A. Simple, heat-sterilizable artificial diet excluding animal-derived ingredients for adult blowfly, Lucilia sericata. Med. Vet. Entomol. 2009, 23, 443–447. [Google Scholar] [CrossRef]
- Stadler, F. The maggot therapy supply chain: A review of the literature and practice. Med. Vet. Entomol. 2020, 34, 1–9. [Google Scholar] [CrossRef] [Green Version]
- Čičková, H.; Newton, G.L.; Lacy, R.C.; Kozánek, M. The use of fly larvae for organic waste treatment. Waste Manag. 2015, 35, 68–80. [Google Scholar] [CrossRef]
- Dindo, M.L.; Grenier, S. Production of dipteran parasitoids. In Mass Production of Beneficial Organisms: Invertebrates and Entomopathogens; Morales-Ramos, J.A., Guadalupe-Rojas, M., Shapiro-Ilan, D.I., Eds.; Academic Press: Cambridge, MA, USA, 2014; pp. 101–143. [Google Scholar]
- Masiero, F.S.; Silva, D.G.; Luchese, M.; Estercio, T.; Persio, N.V.; Thyseen, P.J. In vitro evaluation of the association of medicinal larvae (Insecta, Diptera, Calliphoridae) and topical agents conventionally used for the treatment of wounds. Acta Trop. 2019, 190, 68–72. [Google Scholar] [CrossRef]
- Dindo, M.L.; Rezaei, M.; De Clercq, P. Improvements in the rearing of the tachinid parasitoid Exorista larvarum (Diptera: Tachinidae): Influence of adult food on female longevity and reproduction capacity. J. Insect Sci. 2019, 19, 6. [Google Scholar] [CrossRef] [Green Version]
- Ganda, H.; Abihona, H.A.; Zannou-Boukari, E.T.; Kenis, M.; Chrysostome, C.A.; Mensah, G.A. Influence of adult diet on biological parameters of the housefly, Musca domestica L. (Diptera: Muscidae). J. Basic Appl. Zool. 2020, 81, 46. [Google Scholar] [CrossRef]
- Sheppard, D.C.; Tomberlin, J.K.; Joyce, J.A.; Kiser, B.C.; Sumner, S.M. Rearing methods for the black soldier fly (Diptera: Stratiomyidae). J. Med. Entomol. 2002, 39, 695–698. [Google Scholar] [CrossRef] [Green Version]
- Barragan-Fonseca, K.B.; Dicke, M.; van Loon, J.J. Nutritional value of the black soldier fly (Hermetia illucens L.) and its suitability as animal feed–a review. J. Insects Food Feed 2017, 3, 105–120. [Google Scholar] [CrossRef]
- Belluco, S.; Losasso, C.; Maggioletti, M.; Alonzi, C.C.; Paoletti, M.G.; Ricci, A. Edible insects in a food safety and nutritional perspective: A critical review. Compr. Rev. Food Sci. Food. Saf. 2013, 12, 296–313. [Google Scholar] [CrossRef]
- Van Huis, A. Potential of insects as food and feed in assuring food security. Annu. Rev. Entomol. 2013, 58, 563–583. [Google Scholar] [CrossRef]
- Tessari, P.; Lante, A.; Mosca, G. Essential amino acids: Master regulators of nutrition and environmental footprint? Sci. Rep. 2016, 6, 26074. [Google Scholar] [CrossRef] [Green Version]
- Van Huis, A.; Van Itterbeeck, J.; Klunder, H.; Mertens, E.; Halloran, A.; Muir, G.; Vantomme, P. Edible Insects: Future Prospects for Food and Feed Security; Food and Agriculture Organization of the United Nations: Rome, Italy, 2013. [Google Scholar]
- Oonincx, D.G.; van Broekhoven, S.; van Huis, A.; van Loon, J.J. Feed conversion, survival and development, and composition of four insect species on diets composed of food by-products. PLoS ONE 2015, 10, e0144601. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lamsal, B.; Wang, H.; Pinsirodom, P.; Dossey, A.T. Applications of insect-derived protein ingredients in food and feed industry. J. Am. Oil Chem. Soc. 2019, 96, 105–123. [Google Scholar] [CrossRef]
- Lu, S.; Taethaisong, N.; Meethip, W.; Surakhunthod, J.; Sinpru, B.; Sroichak, T.; Archa, P.; Thongpea, S.; Paengkoum, S.; Patindra-Purba, R.A.; et al. Nutritional composition of black soldier fly larvae (Hermetia illucens L.) and its potential uses as alternative protein sources in animal diets: A review. Insects 2022, 13, 831. [Google Scholar] [CrossRef] [PubMed]
- Makkar, H.P.; Tran, G.; Heuzé, V.; Ankers, P. State-of-the-art on use of insects as animal feed. Anim. Feed Sci. Technol. 2014, 197, 1–33. [Google Scholar] [CrossRef]
- Abd El-Wahab, A.; Meyer, L.; Kölln, M.; Chuppava, B.; Wilke, V.; Visscher, C.; Kamphues, J. Insect larvae meal (Hermetia illucens) as a sustainable protein source of canine food and its impacts on nutrient digestibility and fecal quality. Animals 2021, 11, 2525. [Google Scholar] [CrossRef]
- Penazzi, L.; Schiavone, A.; Russo, N.; Nery, J.; Valle, E.; Madrid, J.; Martínez, S.; Hernández, F.; Pagani, E.; Ala, U.; et al. In vivo and in vitro digestibility of an extruded complete dog food containing black soldier fly (Hermetia illucens) larvae meal as protein source. Front. Vet. Sci. 2021, 8, 542. [Google Scholar] [CrossRef]
- Cappellozza, S.; Leonardi, M.G.; Savoldelli, S.; Carminati, D.; Rizzolo, A.; Cortellino, G.; Terova, G.; Moretto, E.; Badaile, A.; Concheri, G.; et al. A first attempt to produce proteins from insects by means of a circular economy. Animals 2019, 9, 278. [Google Scholar] [CrossRef] [Green Version]
- Pinotti, L.; Ottoboni, M. Substrate as insect feed for biomass production. J. Insects Food Feed 2021, 7, 585–596. [Google Scholar] [CrossRef]
- Kenis, M.; Bouwassi, B.; Boafo, H.; Devic, E.; Han, R.; Koko, G.; Koné, N.; Maciel-Vergara, G.; Nacambo, S.; Charles, S.; et al. Small-scale fly larvae production for animal feed. In Edible Insects in Sustainable Food Systems; Halloran, A., Flore, R., Vantomme, P., Roos, N., Eds.; Springer: Cham, Switzerland, 2018; pp. 239–261. [Google Scholar]
- Romano, N.; Sinha, A.; Powell, A.; Fischer, H. Mineral composition in black soldier fly (Hermetia illucens) larvae and resulting frass from fruit and their peels. J. Insects Food Feed 2022, 9, 43–53. [Google Scholar] [CrossRef]
- Borel, P.; Hammaz, F.; Morand-Laffargue, L.; Creton, B.; Halimi, C.; Sabatier, D.; Desmarchelier, C. Using black soldier fly larvae reared on fruits and vegetables waste as a sustainable dietary source of provitamin a carotenoids. Food Chem. 2021, 359, 129911. [Google Scholar] [CrossRef]
- Melgar-Lalanne, G.; Hernández-Álvarez, A.J.; Salinas-Castro, A. Edible insects processing: Traditional and innovative technologies. Compr. Rev. Food Sci. Food Saf. 2019, 18, 1166–1191. [Google Scholar] [CrossRef] [Green Version]
- Van Huis, A. Insect pests as food and feed. J. Insects Food Feed 2020, 6, 327–331. [Google Scholar] [CrossRef]
- Melo-Ruíz, V.; Vargas-Martínez, N.; Sánchez-Herrera, K.; Rodríguez-Diego, J.; Díaz-Gaarcía, R. The use of fruit fly infested mamey fruit as human food. J. Insects Food Feed 2017, 3, 57–60. [Google Scholar] [CrossRef]
- González, C.M.; Garzón, R.; Rosell, C.M. Insects as ingredients for bakery goods. A comparison study of H. illucens, A. domestica and T. molitor flours. Innov. Food Sci. Emerg. Technol. 2019, 51, 205–210. [Google Scholar] [CrossRef]
- Wang, Y.S.; Shelomi, M. Review of black soldier fly (Hermetia illucens) as animal feed and human food. Foods 2017, 6, 91. [Google Scholar] [CrossRef] [Green Version]
- Parodi, A.; Van Dijk, K.; Van Loon, J.J.; De Boer, I.J.; Van Schelt, J.; Van Zanten, H.H. Black soldier fly larvae show a stronger preference for manure than for a mass-rearing diet. J. Appl. Entomol. 2020, 144, 560–565. [Google Scholar] [CrossRef]
- Van Huis, A.; Rumpold, B.; Van der Fels-Klerx, H.; Tomberlin, J. Advancing edible insects as food and feed in a circular economy. J. Insects Food Feed 2021, 7, 935–948. [Google Scholar] [CrossRef]
- List of Edible Insects of the World. 1 April 2017. Available online: https://www.wur.nl/en/Research-Results/Chair-groups/Plant-Sciences/Laboratory-of-Entomology/Edible-insects/Worldwide-species-list.htm (accessed on 20 November 2022).
- Van Huis, A.; Oonincx, D.G. The environmental sustainability of insects as food and feed. A review. Agron. Sustain. Dev. 2017, 37, 1–14. [Google Scholar] [CrossRef] [Green Version]
- Oonincx, D. Environmental impact of insect rearing. In Insects as Animal Feed: Novel Ingredients for Use in Pet, Aquaculture and Livestock Diets; Hall, H., Fitches, E., Smith, R., Eds.; CABI: Oxfordshire, UK, 2021; pp. 53–59. [Google Scholar]
- Mertenat, A.; Diener, S.; Zurbrügg, C. Black soldier fly biowaste treatment–Assessment of global warming potential. Waste Manag. 2019, 84, 173–181. [Google Scholar] [CrossRef]
- Bang, A.; Courchamp, F. Industrial rearing of edible insects could be a major source of new biological invasions. Ecol. Lett. 2021, 24, 393–397. [Google Scholar] [CrossRef]
- Francuski, L.; Djurakic, M.; Ludoski, J.; Hurtado, P.; Pérez-Bañón, C.; Ståhls, G.; Rojo, S.; Milankov, V. Shift in phenotypic variation coupled with rapid loss of genetic diversity in captive populations of Eristalis tenax (Diptera: Syrphidae): Consequences for rearing and potential commercial use. J. Econ. Entomol. 2014, 107, 821–832. [Google Scholar] [CrossRef] [PubMed]
- Nicholas, S.; Thyselius, M.; Holden, M.; Nordström, K. Rearing and long-term maintenance of Eristalis tenax hoverflies for research studies. J. Vis. Exp. 2018, 135, e57711. [Google Scholar] [CrossRef] [Green Version]
- Wei, X.; Xu, X.; Deloach, C.J. Biological control of white grubs (Coleoptera: Scarabaeidae) by larvae of Promachus yesonicus (Diptera: Asilidae) in China. Biol. Control 1995, 5, 290–296. [Google Scholar] [CrossRef]
- Cohen, C.M.; Cole, T.J.; Brewer, M.S. Pick your poison: Molecular evolution of venom proteins in Asilidae (Insecta: Diptera). Toxins 2020, 12, 738. [Google Scholar] [CrossRef]
- Pollock, D.A.; Davidson, R.L. New records of ground and tiger beetles (Coleoptera: Carabidae) as prey for Asilidae (Diptera) in the high plains of New Mexico and Texas. Coleopt. Bull. 2020, 74, 15–23. [Google Scholar] [CrossRef]
- Dunn, L.; Lequerica, M.; Reid, C.R.; Latty, T. Dual ecosystem services of syrphid flies (Diptera: Syrphidae): Pollinators and biological control agents. Pest Manag. Sci. 2020, 76, 1973–1979. [Google Scholar] [CrossRef]
- Iwai, H.; Niijima, K.; Matsuka, M. An artificial diet for aphidophagous syrphids, Episyrphus balteatus (de Geer) and Eupeodes bucculatus (Rondani) (Diptera: Syrphidae) using drone honeybee brood powder. Appl. Entomol. Zool. 2007, 42, 167–172. [Google Scholar] [CrossRef] [Green Version]
- Grenier, S.; Greany, P.; Cohen, A.C. Potential for mass release of insect parasitoids and predators through development of artificial culture techniques. In Pest Management in the Subtropics: Biological Control—A Florida Perspective; Rosen, D., Bennett, F.D., Capinera, J., Eds.; Intercept Press: Andover, UK, 1994; pp. 181–295. [Google Scholar]
- Gross, H.R.; Rogers, C.E.; Carpenter, J.E. Development of Archytas marmoratus (Diptera: Tachinidae) reared on Galleria mellonella larvae (Lepidoptera: Pyralidae) feeding on selected diets. Biol. Control 1996, 6, 158–163. [Google Scholar] [CrossRef]
- Gross, H.R. Mass propagation of Archytas marmoratus (Diptera: Tachinidae). Environ. Entomol. 1994, 23, 183–189. [Google Scholar] [CrossRef]
- Bratti, A.; Costantini, W. Effects of new artificial host diets on the host–parasitoid system Galleria mellonella L. (Lepidoptera Galleriidae)—Archytas marmoratus Town. (Diptera Tachinidae). Redia 1991, 74, 445–448. [Google Scholar]
- Coulibaly, A.K.; Fanti, P. Eucelatoria bryani Sabr. (Diptera: Tachinidae) rearing on the factitious host Galleria mellonella L. (Lepidoptera: Galleriidae): Effect of host age at exposure to the parasitoid females. Boll. Ist. Entomol. Univ. Bologna 1992, 46, 229–238. [Google Scholar]
- King, E.G.; Hartley, G.G.; Martin, D.F.; Smith, J.W.; Summers, T.E.; Jackson, R.D. Production of the Tachinid Lixophaga diatraeae on Its Natural Host, the Sugar Cane Borer, and on Its Unnatural Host, the Greater Wax Moth; Advances in Agricultural Technology, Southern Series No. 3; U.S. Department of Agriculture, Science and Education Administration: New Orleans, LA, USA, 1979; 16p. [Google Scholar]
- Nettles, W.C.; Wilson, M.; Ziser, S.W. A diet and methods for the in vitro rearing of the tachinid, Eucelatoria sp. Ann. Entomol. Soc. Am. 1980, 73, 180–184. [Google Scholar] [CrossRef]
- Robinson, W. The use of blowfly larva in the treatment of infected wounds. Ann. Entomol. Soc. Am. 1933, 26, 270–276. [Google Scholar] [CrossRef]
- Sherman, R.A.; Pechter, E.A. Maggot therapy—A review of the therapeutic applications of fly larvae in human medicine, especially for treating osteomyelitis. Med. Vet. Entomol. 1988, 2, 225–230. [Google Scholar] [CrossRef]
- Sherman, R.A.; Wyle, F.A. Low-cost, low-maintenance rearing of maggots in hospitals, clinics, and schools. Am. J. Trop. Med. Hyg. 1996, 54, 38–41. [Google Scholar] [CrossRef]
- Baer, W.S. The classic: The treatment of chronic osteomyelitis with the maggot (larva of the blow fly). Clin. Orthop. Relat. Res. 2011, 469, 920–944. [Google Scholar] [CrossRef] [Green Version]
- McKeever, D.C. The classic: Maggots in treatment of osteomyelitis: A simple inexpensive method. Clin. Orthop. Relat. Res. 2008, 466, 1329–1335. [Google Scholar] [CrossRef] [Green Version]
- Tachibana, S.I.; Numata, H. An artificial diet for blow fly larvae, Lucilia sericata (Meigen) (Diptera: Calliphoridae). Appl. Entomol. Zool. 2001, 36, 521–523. [Google Scholar] [CrossRef] [Green Version]
- Connelly, K.L.; Freeman, E.; Smibert, O.C.; Lin, B. Wohlfahrtiimonas chitiniclastica bloodstream infection due to a maggot-infested wound in a 54-year-old male. J. Glob. Infect. Dis. 2019, 11, 125–126. [Google Scholar] [CrossRef]
- Bueide, P.J.; Hunt, J.; Bande, D.; Guerrero, D.M. Maggot wound therapy associated with Wohlfahrtiimonas chitiniclastica blood infection. Cureus 2021, 13, e12471. [Google Scholar] [CrossRef]
- Naik, G.; Harding, K.G. Maggot debridement therapy: The current perspectives. Chronic Wound Care Manag. Res. 2017, 4, 121–128. [Google Scholar] [CrossRef] [Green Version]
- Chambers, L.; Woodrow, S.; Brown, A.P.; Harris, P.D.; Phillips, D.; Hall, M.; Church, J.C.T.; Pritchard, D.I. Degradation of extracellular matrix components by defined proteinases from the greenbottle larva Lucilia sericata used for the clinical debridement of non-healing wounds. Br. J. Dermatol. 2003, 148, 14–23. [Google Scholar] [CrossRef] [PubMed]
- Masiero, F.S.; Aquino, M.F.K.; Nassu, M.P.; Pereira, D.I.B.; Leite, D.S.; Thyssen, P.J. First record of larval secretions of Cochliomyia macellaria (Fabricius 1775) (Diptera: Calliphoridae) inhbiting growth of Staphylococcus aureus and Pseudomonas aeruginosa. Neotrop. Entomol. 2017, 46, 125–129. [Google Scholar] [CrossRef] [PubMed]
- Bexfield, A.; Nigam, Y.; Thomas, S.; Ratcliffe, N.A. Detection and partial characterization of two antibacterial factors from the excretions/secretions of the medicinal maggot Lucilia sericata and their activity against methicillin-resistant Staphylococcus aureus (MRSA). Microbes Infect. 2004, 6, 1297–1304. [Google Scholar] [CrossRef]
- Bexfield, A.; Bond, A.E.; Morgan, C.; Wagstaff, J.; Newton, R.P.; Ratcliffe, N.A.; Dudley, E.; Nigam, Y. Amino acid derivatives from Lucilia sericata excretions/secretions may contribute to the beneficial effects of maggot therapy via increased angiogenesis. Br. J. Dermatol. 2010, 162, 554–562. [Google Scholar] [CrossRef] [PubMed]
- Sherman, R.A.; Hall, M.J.R.; Thomas, S. Medicinal maggots: An ancient remedy for some contemporary afflictions. Annu. Rev. Entomol. 2000, 45, 55–81. [Google Scholar] [CrossRef]
- Sherman, R.A.; Sherman, J.; Gilead, L.; Lipo, M.; Mumcuoglu, K.Y. Maggot debridement therapy in outpatients. Arch. Phys. Med. Rehabil. 2001, 82, 1226–1229. [Google Scholar] [CrossRef]
- Wilson, M.R.; Nigam, Y.; Knight, J.; Pritchard, D.I. What is the optimal treatment time for larval therapy? A study on incubation time and tissue debridement by bagged maggots of the greenbottle fly, Lucilia sericata. Int. Wound J. 2019, 16, 219–225. [Google Scholar] [CrossRef] [Green Version]
- Benecke, M. A brief survey of the history of forensic entomology. Acta Biol. Benrodis 2008, 14, 15–38. [Google Scholar]
- Tomberlin, J.K.; Mohr, R.; Benbow, M.E.; Tarone, A.M.; VanLaerhoven, S. A roadmap for bridging basic and applied research in forensic entomology. Annu. Rev. Entomol. 2011, 56, 401–421. [Google Scholar] [CrossRef] [Green Version]
- Cammack, J.A.; Cohen, A.C.; Kreitlow, K.L.; Roe, R.M.; Watson, D.W. Decomposition of concealed and exposed porcine remains in the North Carolina Piedmont. J. Med. Entomol. 2016, 53, 67–75. [Google Scholar] [CrossRef]
- Vélez, M.C.; Wolff, M. Rearing five species of Diptera (Calliphoridae) of forensic importance in Colombia in semicontrolled field conditions. Pap. Avulsos Zool. 2008, 48, 41–47. [Google Scholar] [CrossRef] [Green Version]
- Rabêlo, K.C.; Thyssen, P.J.; Salgado, R.L.; Araújo, M.S.; Vasconcelos, S.D. Bionomics of two forensically important blowfly species Chrysomya megacephala and Chrysomya putoria (Diptera: Calliphoridae) reared on four types of diet. Forensic Sci. Int. 2011, 210, 257–262. [Google Scholar] [CrossRef]
- Noblesse, A.P.; Meeds, A.W.; Weidner, L.M. Blow flies (Diptera: Calliphoridae) and the American diet: Effects of fat content on blow fly development. J. Med. Entomol. 2022, 59, 1191–1197. [Google Scholar] [CrossRef]
- Owings, C.; McQueen, M.; Smith, M.; Wal, R.; McKee-Zech, H. Rearing container size impacts immature development time of Phormia regina (Meigen) (Diptera: Calliphoridae) and time of colonization estimations. J. Forensic Entomol. 2022. Available online: https://jfe-ojs-tamu.tdl.org/jfe/article/view/6 (accessed on 11 June 2023).
- Gruszka, J.; Matuszewski, S. Insect rearing protocols in forensic entomology: Benefits from collective rearing of larvae in a carrion beetle Necrodes littoralis L. (Silphidae). PLoS ONE 2021, 16, e0260680. [Google Scholar] [CrossRef]
- Sherman, R.A.; Tran, J.M.T. A simple, sterile food source for rearing the larvae of Lucilia sericata (Diptera: Calliphoridae). Med. Vet. Entomol. 1995, 9, 393–398. [Google Scholar] [CrossRef]
- Weidner, L.M.; Nigoghosian, G.; Hanau, C.G.; Jennings, D.E. Analysis of alternative food sources for rearing entomological evidence. J. Med. Entomol. 2020, 57, 1407–1410. [Google Scholar] [CrossRef]
- Chaudhury, M.F.; Skoda, R. An artificial diet for rearing Cochliomyia macellaria (Diptera: Calliphoridae). J. Econ. Entomol. 2013, 106, 1927–1931. [Google Scholar] [CrossRef]
- Zuha, R.; Jenarthanan, L.; Disney, R.; Omar, B. Multiple species of scuttle flies (Diptera: Phoridae) as contaminants in forensic entomology laboratory insect colony. Trop. Biomed. 2015, 32, 568–572. [Google Scholar]
- Estrada, D.A.; Grella, M.D.; Thyssen, P.; Linhares, A.X. Chrysoma albiceps (Diptera: Calliphoridae) developmental rate on artificial diet with animal tissues for forensic purpose. Neotrop. Entomol. 2009, 38, 203–207. [Google Scholar] [CrossRef] [PubMed]
- Kökdener, M.; Gündüz, N.E.A.; Zeybekoğlu, Ü.; Aykut, U.; Yılmaz, A.F. The effect of different heavy metals on the development of Lucilia sericata (Diptera: Calliphoridae). J. Med. Entomol. 2022, 59, 1928–1935. [Google Scholar] [CrossRef] [PubMed]
- Tabor, K.L.; Fell, R.D.; Brewster, C.C.; Pelzer, K.; Behonick, G.S. Effects of antemortem ingestion of ethanol on insect successional patterns and development of Phormia regina (Diptera: Calliphoridae). J. Med. Entomol. 2005, 42, 481–489. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Grassberger, M.; Reiter, C. Effect of temperature on Lucilia sericata (Diptera: Calliphoridae) development with special reference to the isomegalen-and isomorphen-diagram. Forensic Sci. Int. 2001, 120, 32–36. [Google Scholar] [CrossRef]
- Grassberger, M.; Reiter, C. Effect of temperature on development of Liopygia (= Sarcophaga) argyrostoma (Robineau-Desvoidy) (Diptera: Sarcophagidae) and its forensic implications. J. Forensic Sci. 2002, 47, 1332–1336. [Google Scholar] [CrossRef]
- Marchenko, M.I. Medicolegal relevance of cadaver entomofauna for the determination of the time of death. Forensic Sci. Int. 2001, 120, 89–109. [Google Scholar] [CrossRef]
- Arias, A.M. Drosophila melanogaster and the development of biology in the 20th century. In Methods in Molecular Biology: Drosophila: Methods and Protocols; Dahmann, C., Ed.; Humana Press: Totowa, NJ, USA, 2008; pp. 1–25. [Google Scholar]
- Jennings, B.H. Drosophila—A versatile model in biology & medicine. Mater. Today 2011, 14, 190–195. [Google Scholar] [CrossRef]
- Tower, J. Drosophila flies in the face of aging. J. Gerontol. A Biol. Sci. Med. Sci. 2019, 74, 1539–1541. [Google Scholar] [CrossRef] [Green Version]
- Reaume, C.J.; Sokolowski, M.B. The nature of Drosophila melanogaster. Curr. Biol. 2006, 16, R623–R628. [Google Scholar] [CrossRef] [Green Version]
- Roberts, D.B. Drosophila melanogaster: The model organism. Entomol. Exp. Appl. 2006, 121, 93–103. [Google Scholar] [CrossRef]
- Popis, M.; Borowiec, B.; Jankowski, M. Drosophila melanogaster research: History, breakthrough and perspectives. Med. J. Cell. Biol. 2018, 6, 182–185. [Google Scholar] [CrossRef] [Green Version]
- Reiter, L.T.; Potocki, L.; Chien, S.; Gribskov, M.; Bier, E. A systematic analysis of human disease-associated gene sequences in Drosophila melanogaster. Genome Res. 2001, 11, 1114–1125. [Google Scholar] [CrossRef] [Green Version]
- Brumby, A.M.; Richardson, H.E. Using Drosophila melanogaster to map human cancer pathways. Nat. Rev. Cancer 2005, 5, 626–639. [Google Scholar] [CrossRef]
- Mirzoyan, Z.; Sollazzo, M.; Allocca, M.; Valenza, A.M.; Grifoni, D.; Bellosta, P. Drosophila melanogaster: A model organism to study cancer. Front. Genet. 2019, 10, 51. [Google Scholar] [CrossRef] [Green Version]
- Sandin, L.; Bergkvist, L.; Sangeeta, N.; Kielkopf, C.; Janefjord, C.; Helmfors, L.; Zetterberg, H.; Blennow, K.; Li, H.; Nilsberth, C.; et al. Beneficial effects of increased lysozyme levels in Alzheimer’s disease modelled in Drosophila melanogaster. FEBS J. 2016, 283, 3508–3522. [Google Scholar] [CrossRef] [Green Version]
- Tue, N.T.; Dat, T.Q.; Ly, L.L.; Anh, V.D.; Yoshida, H. Insights from Drosophila melanogaster model of Alzheimer’s disease. Front. Biosci.-Landmark 2020, 25, 134–146. [Google Scholar] [CrossRef]
- Aryal, B.; Lee, Y. Disease model organism for Parkinson disease: Drosophila melanogaster. BMB Rep. 2019, 52, 250–258. [Google Scholar] [CrossRef] [Green Version]
- Alfa, R.W.; Kim, S.K. Using Drosophila to discover mechanisms underlying type 2 diabetes. Dis. Model. Mech. 2016, 9, 365–376. [Google Scholar] [CrossRef] [Green Version]
- Meshrif, W.S.; El Husseiny, I.M.; Elbrense, H. Drosophila melanogaster as a low-cost and valuable model for studying type 2 diabetes. J. Exp. Zool. A Ecol. Integr. Physiol. 2022, 337, 457–466. [Google Scholar] [CrossRef]
- Baenas, N.; Wagner, A.E. Drosophila melanogaster as a model organism for obesity and type-2 diabetes mellitus by applying high-sugar and high-fat diets. Biomolecules 2022, 12, 307. [Google Scholar] [CrossRef]
- Zerofsky, M.; Harel, E.; Silverman, N.; Tatar, M. Aging of the innate immune response in Drosophila melanogaster. Aging Cell 2005, 4, 103–108. [Google Scholar] [CrossRef] [PubMed]
- Aceituno-Medina, M.; Rivera-Ciprian, J.P.; Hernández, E. Evaluation of a pelleted diet for larval mass-rearing of Anastrepha ludens and Anastrepha obliqua. Entomol. Exp. Appl. 2020, 168, 502–512. [Google Scholar] [CrossRef]
- Pascacio-Villafán, C.; Birke, A.; Williams, T.; Aluja, M. Modeling the cost-effectiveness of insect rearing on artificial diets: A test with a tephritid fly used in the sterile insect technique. PLoS ONE 2017, 12, e0173205. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Pascacio-Villafán, C.; Guillén, L.; Aluja, M. Agar and carrageenan as cost-effective gelling agents in yeast-reduced artificial diets for mass-rearing fruit flies and their parasitoids. Insects 2020, 11, 131. [Google Scholar] [CrossRef] [Green Version]
- Aluja, M.; Zamora-Briseño, J.A.; Pérez-Brocal, V.; Altúzar-Molina, A.; Guillén, L.; Desgarennes, D.; Vázquez-Rosas-Landa, M.; Ibarra-Laclette, E.; Alonso-Sánchez, A.G.; Moya, A. Metagenomic survey of the highly polyphagous Anastrepha ludens developing in ancestral and exotic hosts reveals the lack of a stable microbiota in larvae and the strong influence of metamorphosis on adult gut microbiota. Front. Microbiol. 2021, 12, 685937. [Google Scholar] [CrossRef]
- Ochoa-Sánchez, M.; Cerqueda-García, D.; Moya, A.; Ibarra-Laclette, E.; Altúzar-Molina, A.; Desgarennes, D.; Aluja, M. Bitter friends are not always toxic: The loss of acetic acid bacteria and the absence of Komagataeibacter in the gut microbiota of the polyphagous fly Anastrepha ludens could inhibit its development in Psidium guajava in contrast to A. striata and A. fraterculus that flourish in this host. Front. Microbiol. 2022, 13, 979817. [Google Scholar] [CrossRef]
- Heed, W.B.; Starmer, W.T.; Miranda, M.; Miller, M.W.; Phaff, H.J. An analysis of yeast flora associated with cactiphilic Drosophila and their host plants in the Sonoran Desert and its relation to temperate and tropical associations. Ecology 1976, 57, 51–160. [Google Scholar] [CrossRef]
- Vacek, D.C.; Starmer, W.T.; Heed, W.B. Relevance of the ecology of citrus yeasts to the diet of Drosophila. Microb. Ecol. 1979, 5, 43–49. [Google Scholar] [CrossRef]
- Starmer, W.T.; Fogleman, J.C. Coadaptation of Drosophila and yeasts in their natural habitat. J. Chem. Ecol. 1986, 12, 1037–1055. [Google Scholar] [CrossRef]
- Starmer, W.T.; Barker, J.; Phaff, H.J.; Fogleman, J.C. Adaptations of Drosophila and yeasts: Their interactions with the volatile 2-propanol in the cactus-microorganism-Drosophila model system. Aust. J. Biol. Sci. 1986, 39, 69–77. [Google Scholar] [CrossRef]
- Anagnostou, C.; Dorsch, M.; Rohlfs, M. Influence of dietary yeasts on Drosophila melanogaster life-history traits. Entomol. Exp. Appl. 2010, 136, 1–11. [Google Scholar] [CrossRef]
- Deutscher, A.T.; Reynolds, O.L.; Chapman, T.A. Yeast: An overlooked component of Bactrocera tryoni (Diptera: Tephritidae) larval gut microbiota. J. Econ. Entomol. 2017, 110, 298–300. [Google Scholar] [CrossRef]
- Piper, A.M.; Farnier, K.; Linder, T.; Speight, R.; Cunningham, J.P. Two gut-associated yeasts in a tephritid fruit fly have contrasting effects on adult attraction and larval survival. J. Chem. Ecol. 2017, 43, 891–901. [Google Scholar] [CrossRef]
- Nikolouli, K.; Sassù, F.; Ntougias, S.; Stauffer, C.; Cáceres, C.; Bourtzis, K. Enterobacter sp. AA26 as a protein source in the larval diet of Drosophila suzukii. Insects 2021, 12, 923. [Google Scholar] [CrossRef]
- Nguyen, B.; Ponton, F.; Than, A.; Taylor, P.W.; Chapman, T.; Morimoto, J. Interactions between ecological factors in the developmental environment modulate pupal and adult traits in a polyphagous fly. Ecol. Evol. 2019, 9, 6342–6352. [Google Scholar] [CrossRef]
- Morimoto, J.; Than, A.T.; Nguyen, B.; Lundbäck, I.; Dinh, H.; Ponton, F. Density-by-diet interactions during larval development shape adult life history trait expression and fitness in a polyphagous fly. Am. Nat. 2022, 199, E170–E185. [Google Scholar] [CrossRef]
- Pascacio-Villafán, C.; Righini, N.; Nestel, D.; Birke, A.; Guillén, L.; Aluja, M. Diet quality and conspecific larval density predict functional trait variation and performance in a polyphagous frugivorous fly. Funct. Ecol. 2022, 36, 1163–1176. [Google Scholar] [CrossRef]
- Henry, Y.; Renault, D.; Colinet, H. Hormesis-like effect of mild larval crowding on thermotolerance in Drosophila flies. J. Exp. Biol. 2018, 221, jeb169342. [Google Scholar] [CrossRef] [Green Version]
- Nakamura, S. Inbreeding and rotational breeding of the parasitoid fly, Exorista japonica (Diptera: Tachinidae), for successive rearing. Appl. Entomol. Zool. 1996, 31, 433–441. [Google Scholar] [CrossRef] [Green Version]
- Li, Z.; Yang, D.; Huang, M.; Hu, X.; Shen, J.; Zhao, Z.; Chen, J. Chrysomya megacephala (Fabricius) larvae: A new biodiesel resource. Appl. Energy 2012, 94, 349–354. [Google Scholar] [CrossRef]
- Reddy, P.R.; Rajan, V.V.; Verghese, A. A non-meat-based artificial diet and protocol for mass rearing of Chrysomya megacephala (Fab.) (Diptera: Calliphoridae), an important pollinator of mango. Curr. Sci. 2015, 108, 17–19. [Google Scholar]
- Chen, H.; Chaudhury, M.F.; Sagel, A.; Phillips, P.L.; Skoda, S.R. Artificial diets used in mass production of the New World screwworm, Cochliomyia hominivorax. J. Appl. Entomol. 2014, 138, 708–714. [Google Scholar] [CrossRef] [Green Version]
- Terras, M.A.; Rose, H.A. An artificial diet for Lucilia cuprina (Weidemann) and the effects of two diets on aldrin epoxidase activity. J. Aust. Entomol. Soc. 1985, 24, 121–123. [Google Scholar] [CrossRef]
- Firoozfar, F.; Moosa-Kazemi, H.; Baniardalani, M.; Abolhassani, M.; Khoobdel, M.; Rafinejd, J. Mass rearing of Lucilia sericata Meigen (Diptera: Calliphoridae). Asian Pac. J. Trop. Biomed. 2011, 1, 54–56. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Green, P.W.; Simmonds, M.S.; Blaney, W.M. Diet nutriment and rearing density affect the growth of black blowfly larvae, Phormia regina (Diptera: Calliphoridae). Eur. J. Entomol. 2003, 100, 39–42. [Google Scholar] [CrossRef] [Green Version]
- Prado e Castro, C.; Ameixa, O.M.C.C. Blow flies (Diptera: Calliphoridae) promising candidates as animal feed ingredients. J. Insects Food Feed 2021, 7, 1065–1076. [Google Scholar] [CrossRef]
- Piper, M.D.W.; Blanc, E.; Leitão-Gonçalves, R.; Yang, M.; He, X.; Linford, N.J.; Hoddinott, M.P.; Hopfen, C.; Soultoukis, G.A.; Niemeyer, C.; et al. A holidic medium for Drosophila melanogaster. Nat. Methods 2014, 11, 100–105. [Google Scholar] [CrossRef] [Green Version]
- Matos, M.; Rose, M.R.; Rocha Pité, M.T.; Rego, C.; Avelar, T. Adaptation to the laboratory environment in Drosophila subobscura. J. Evol. Biol. 2000, 13, 9–19. [Google Scholar] [CrossRef]
- Aceituno-Medina, M.; Ordoñez, A.; Carrasco, M.; Montoya, P.; Hernández, E. Mass rearing, quality parameters, and bioconversion in Drosophila suzukii (Diptera: Drosophilidae) for Sterile Insect Technique purposes. J. Econ. Entomol. 2020, 113, 1097–1104. [Google Scholar] [CrossRef]
- Bradley, T.J.; Herbst, D.B. Growth and survival of larvae of Ephydra hians Say (Diptera: Ephydridae) on unialgal diets. Environ. Entomol. 1994, 23, 276–281. [Google Scholar] [CrossRef]
- Holderman, C.J.; Sanchez-Sandoval, U.A.; Ramirez, J.; Smythe, B.G. Laboratory methods for rearing horn flies (Diptera: Muscidae). J. Insect Sci. 2020, 20, 10. [Google Scholar] [CrossRef]
- Byrd, J.H. Laboratory rearing of forensic insects. In Forensic Entomology: The Utility of Arthropods in Legal Investigations; Byrd, J.H., Castner, J.L., Eds.; CRC Press: Boca Raton, FL, USA, 2001; pp. 121–142. [Google Scholar]
- Wang, Y.; Zhang, Y.; Wang, M.; Hu, G.; Fu, Y.; Zhi, R.; Wang, J. Development of Hydrotaea spinigera (Diptera: Muscidae) at constant temperatures and its significance for estimating postmortem interval. J. Med. Entomol. 2021, 58, 56–63. [Google Scholar] [CrossRef]
- Hamidou Leyo, I.; Moussa Ousmane, Z.; Noël, G.; Francis, F.; Caparros Megido, R. Breeding enhancement of Musca domestica L. 1758: Egg load as a measure of optimal larval density. Insects 2021, 12, 956. [Google Scholar] [CrossRef]
- Ardburi, W.; Tangkawanit, U. Effect of larval diets on house fly (Diptera: Muscidae) production and host suitability for a pupal house fly parasitoid (Spalangia gemina) (Hymenoptera: Pteromalidae). J. Med. Entomol. 2022, 59, 1376–1381. [Google Scholar] [CrossRef]
- Lahuatte, P.F.; Lincango, M.P.; Heimpel, G.E.; Causton, C.E. Rearing larvae of the avian nest parasite, Philornis downsi (Diptera: Muscidae), on chicken blood-based diets. J. Insect Sci. 2016, 16, 84. [Google Scholar] [CrossRef] [Green Version]
- Salem, A.; Franc, M.; Jacquiet, P.; Bouhsira, E.; Liénard, E. Feeding and breeding aspects of Stomoxys calcitrans (Diptera: Muscidae) under laboratory conditions. Parasite 2012, 19, 309. [Google Scholar] [CrossRef] [Green Version]
- Friesen, K.; Berkebile, D.R.; Zhu, J.J.; Taylor, D.B. Laboratory rearing of stable flies and other muscoid Diptera. J. Vis. Exp. 2018, 138, e57341. [Google Scholar] [CrossRef] [Green Version]
- Dillmann, J.B.; Cossetin, L.; dos Santos Petry, L.; de Souza, T.P.; Monteiro, S.G. An alternative protocol for rearing of stable fly, Stomoxys calcitrans (Diptera: Muscidae) under laboratory conditions. Int. J. Trop. Insect Sci. 2021, 41, 2453–2458. [Google Scholar] [CrossRef]
- Yehuda, B.; Marchaim, U.; Glatman, L.; Drabkin, V.; Chizov-Ginzburg, A.; Mumcuoglu, K.Y.; Gelman, A. Bioconversion of poultry and fish waste by Lucilia sericata and Sarcophaga carnaria larvae. Asian J. Water Environ. Pollut. 2011, 8, 69–75. [Google Scholar]
- Almeida, C.; Rijo, P.; Rosado, C. Bioactive compounds from Hermetia illucens larvae as natural ingredients for cosmetic application. Biomolecules 2020, 10, 976. [Google Scholar] [CrossRef]
- Kim, C.H.; Ryu, J.; Lee, J.; Ko, K.; Lee, J.Y.; Park, K.Y.; Chung, H. Use of black soldier fly larvae for food waste treatment and energy production in Asian countries: A review. Processes 2021, 9, 161. [Google Scholar] [CrossRef]
- Arnone, S.; De Mei, M.; Petrazzuolo, F.; Musmeci, S.; Tonelli, L.; Salvicchi, A.; Defilippo, F.; Curatolo, M.; Bonilauri, P. Black soldier fly (Hermetia illucens L.) as a high-potential agent for bioconversion of municipal primary sewage sludge. Environ. Sci. Pollut. Res. 2022, 29, 64886–64901. [Google Scholar] [CrossRef] [PubMed]
- Campoy, A.; Lutsyk, M.; Pérez-Bañón, C.; Rojo, S. Age-stage two-sex life table analysis of Eristalinus aeneus (Diptera, Syrphidae) reared with two different larval media. Bull. Entomol. Res. 2022, 112, 13–20. [Google Scholar] [CrossRef] [PubMed]
- Ottenheim, M.M.; Holloway, G.J. The effect of diet and light on larval and pupal development of laboratory-reared Eristalis arbustorum (Diptera: Syrphidae). Neth. J. Zool. 1995, 45, 305–314. [Google Scholar]
- Martel, V.; Thireau, J.C.; Régnière, J. Manual inoculation of host larvae with first instar maggots as a rearing technique for the larval parasitoid Actia interrupta (Diptera: Tachinidae). Biocontrol Sci. Technol. 2022, 32, 110–113. [Google Scholar] [CrossRef]
- Bratti, A.; Nettles, W.C., Jr. In vitro rearing of Eucelatoria bryani: Improvements and evaluation of factors affecting efficiency. Entomol. Exp. Appl. 1992, 63, 213–219. [Google Scholar] [CrossRef]
- Dindo, M.L.; Vandicke, J.; Marchetti, E.; Spranghers, T.; Bonte, J.; De Clercq, P. Supplementation of an artificial medium for the parasitoid Exorista larvarum (Diptera: Tachnidae) with hemolymph of Hermetia illucens (Diptera: Stratiomyidae) or Antheraea pernyi (Lepidoptera: Saturniidae). J. Econ. Entomol. 2016, 109, 602–606. [Google Scholar] [CrossRef]
- Cardoza, Y.J.; Epsky, N.D.; Heath, R.R. Biology and development of Lespesia aletiae (Diptera: Tachinidae) in two lepidopteran species in the laboratory. Fla. Entomol. 1997, 80, 289–300. [Google Scholar] [CrossRef]
- Rodriguez-del-Bosque, L.A.; Smith, J.W., Jr. Rearing and biology of Lydella jalisco (Diptera: Tachinidae), a parasite of Eoreuma loftini (Lepidoptera: Pyralidae) from Mexico. Ann. Entomol. Soc. Am. 1996, 89, 88–95. [Google Scholar] [CrossRef]
- Nagarkatti, S.; Rao, V.P. Biology of and rearing technique for Sturmiopsis parasitica (Curr.) (Diptera, Tachinidae), a parasite of graminaceous borers in Africa. Bull. Entomol. Res. 1975, 65, 165–170. [Google Scholar] [CrossRef]
- Aluja, M.; Sivinski, J.; Ovruski, S.; Guillen, L.; López, M.; Cancino, J.; Torres-Anaya, A.; Gallegos-Chan, G.; Ruíz, L. Colonization and domestication of seven species of native New World hymenopterous larval-prepupal and pupal fruit fly (Diptera: Tephritidae) parasitoids. Biocontrol Sci. Technol. 2009, 19 (Suppl. S1), 49–79. [Google Scholar] [CrossRef]
- Orozco-Dávila, D.; Quintero, L.; Hernández, E.; Solís, E.; Artiaga, T.; Hernández, R.; Ortega, C.; Montoya, P. Mass rearing and sterile insect releases for the control of Anastrepha spp. pests in Mexico–A review. Entomol. Exp. Appl. 2017, 164, 176–187. [Google Scholar] [CrossRef] [Green Version]
- Castro-López, C.; Pascacio-Villafán, C.; Aluja, M.; García, H.S.; González-Córdova, A.F.; Vallejo-Cordoba, B.; Hernández-Mendoza, A. Safety assessment of the potential probiotic bacterium Limosilactobacillus fermentum J23 using the Mexican fruit fly (Anastrepha ludens Loew, Diptera: Tephritidae) as a novel in vivo model. Probiotics Antimicrob. Proteins 2022. advance online publication. [Google Scholar] [CrossRef]
- Rasgado-Marroquín, M.A.; Velázquez-Dávila, E.; De la Cruz-De la Cruz, J.A.; Laparra, R.A.; Villareal, L.C.S.; Tejeda, M.T. A new diet for a new facility development of a starter-finalizer diet system for rearing colonies of the Ceratitis capitata Vienna 8 strain at a new facility of Mexico’s Moscamed program. In Area-Wide Management of Fruit Fly Pests; Pérez-Staples, D., Díaz-Flescher, F., Montoya, P., Vera, M.T., Eds.; CRC Press: Boca Raton, FL, USA, 2019; pp. 217–232. [Google Scholar]
- Chang, C.L.; Albrecht, C.; El-Shall, S.S.; Kurashima, R. Adult reproductive capacity of Ceratitis capitata (Diptera: Tephritidae) on a chemically defined diet. Ann. Entomol. Soc. Am. 2001, 94, 702–706. [Google Scholar] [CrossRef] [Green Version]
- Wiegers, G.L.; Dullemans, A.M.; Wijbenga, J. The rearing of Tipula oleracea L.(Dipt., Tipulidae). J. Appl. Entomol. 1992, 114, 410–414. [Google Scholar] [CrossRef]

Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Pascacio-Villafán, C.; Cohen, A.C. How Rearing Systems for Various Species of Flies Benefit Humanity. Insects 2023, 14, 553. https://doi.org/10.3390/insects14060553
Pascacio-Villafán C, Cohen AC. How Rearing Systems for Various Species of Flies Benefit Humanity. Insects. 2023; 14(6):553. https://doi.org/10.3390/insects14060553
Chicago/Turabian StylePascacio-Villafán, Carlos, and Allen Carson Cohen. 2023. "How Rearing Systems for Various Species of Flies Benefit Humanity" Insects 14, no. 6: 553. https://doi.org/10.3390/insects14060553