
Receptivity and Remating Propensity in Female Spodoptera litura (Fabricius) after Mating with an Irradiated Male or Its F1 Male Progeny
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:Simple Summary
Abstract
1. Introduction
2. Materials and Methods
2.1. Insect Rearing
2.2. Insect Irradiation
2.3. Precopulatory, Mating and Reproductive Behavior of Mated Female Moths
2.4. Insemination Quality of Irradiated Moths and F1 Progeny
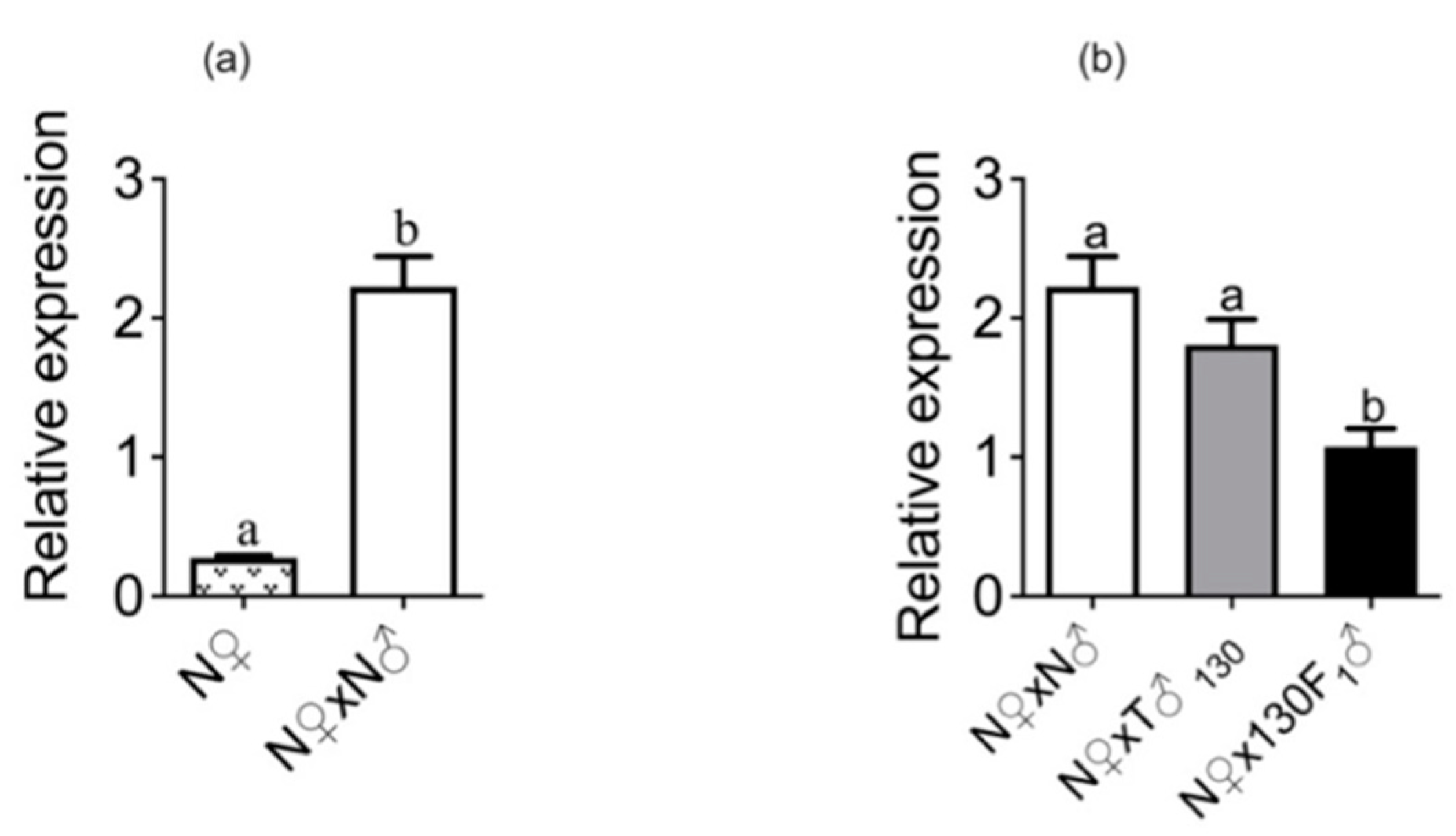
2.5. Expression of Sex Peptide Receptor (SPR) in Mated Females
2.6. Remating Behavior of Female Moths
2.7. Statistical Analysis
3. Results
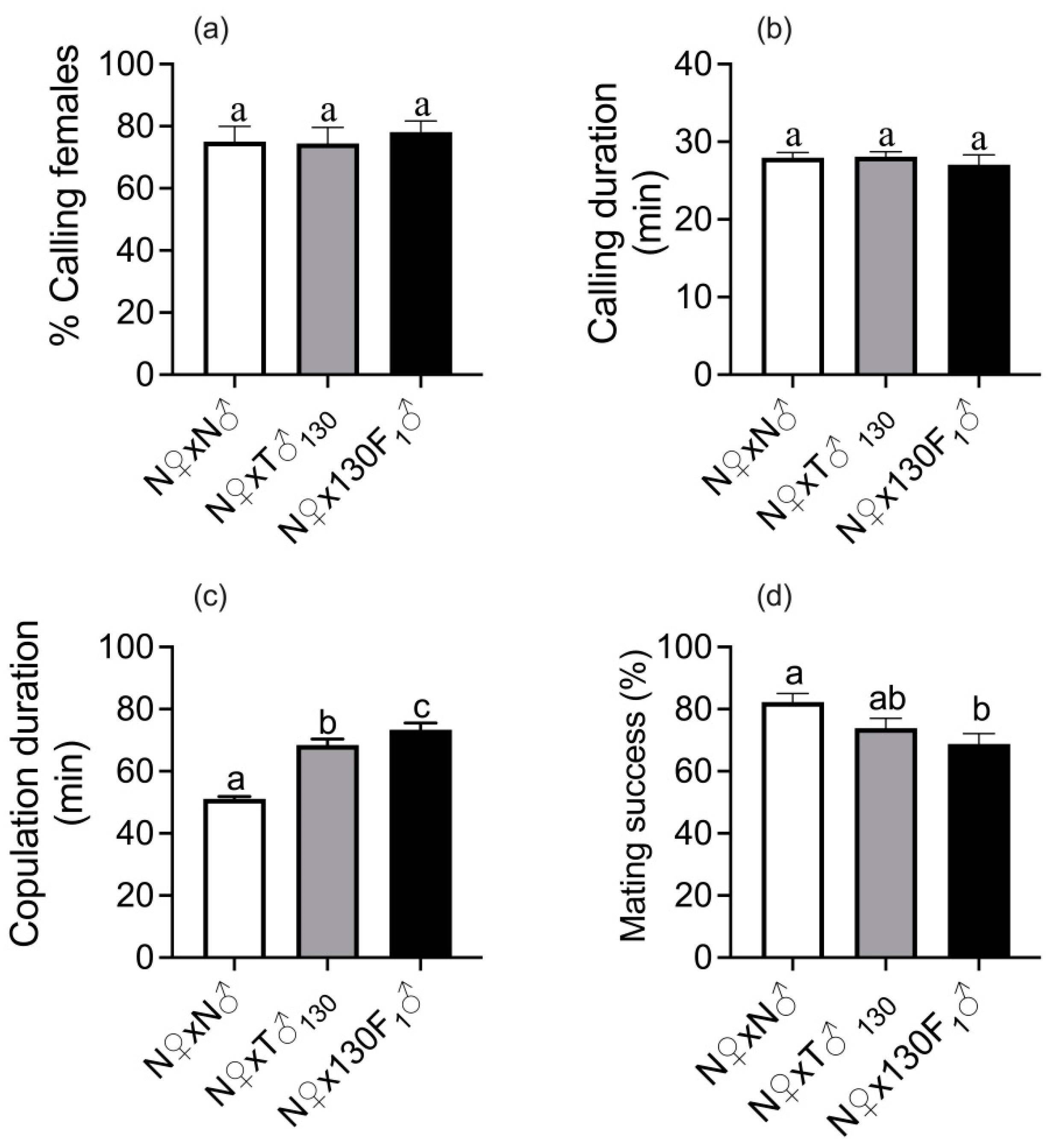
3.1. Female Premating and Mating Behavior after Pairing with Irradiated Males and F1 Male Progeny
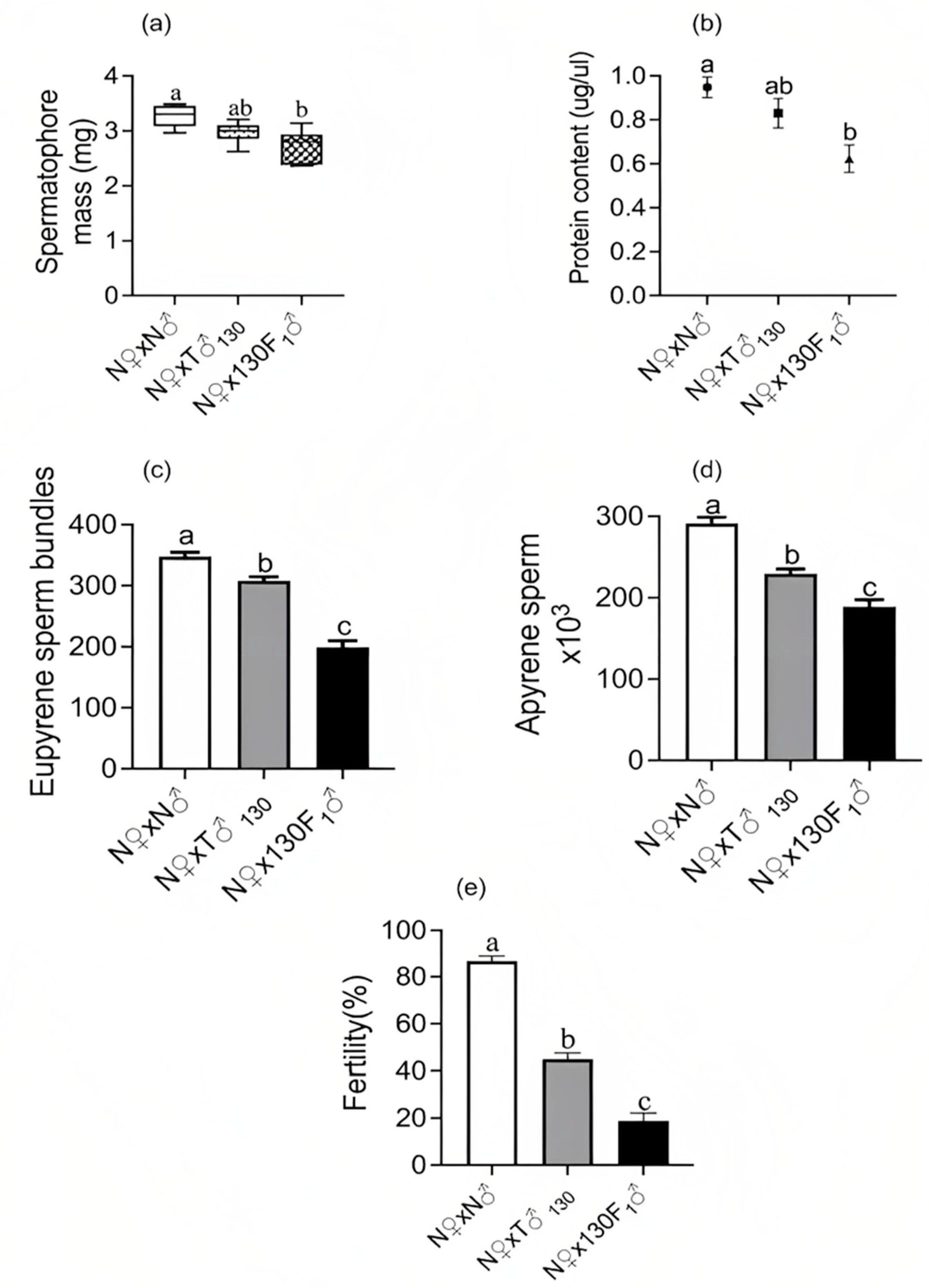
3.2. Insemination Quality of Irradiated Moths and F1 Progeny and Its Correlation with Fertility
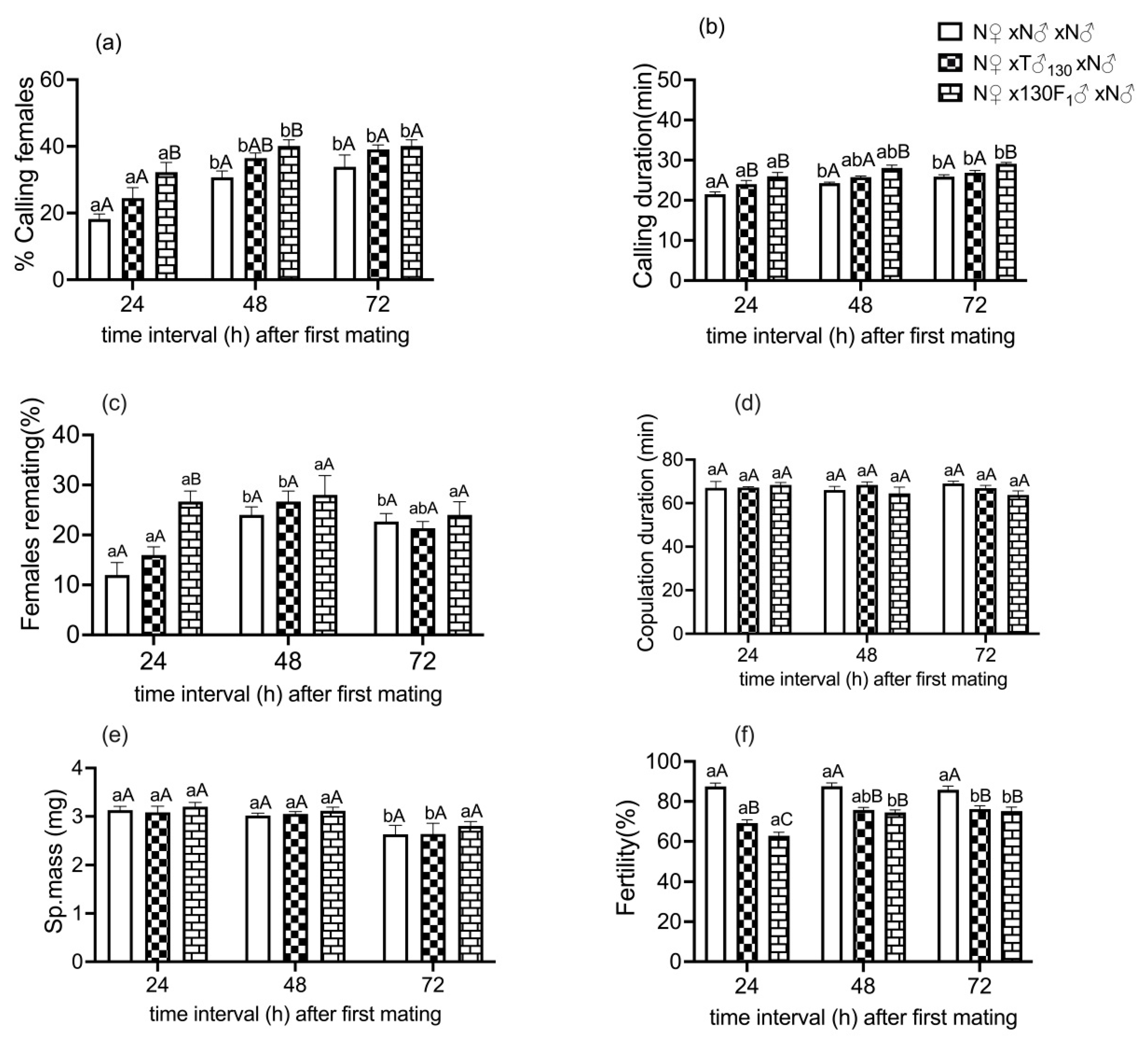
3.3. Remating Behavior of Mated Females in Response to First Mating with Irradiated Males or Their F1 Progeny
4. Discussion
4.1. Premating and Mating Behavior after Pairing with Irradiated Males and F1 Male Progeny
4.2. Insemination Quality of Irradiated Males and F1 Male Progeny and Its Correlation with Fertility
4.3. Remating Behavior in Response to First Mating with Irradiated Males or Their F1 Progeny
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Ahmad, M.; Arif, M.I.; Ahmad, M. Occurrence of insecticide resistance in field populations of Spodoptera litura (Lepidoptera: Noctuidae) in Pakistan. Crop Prot. 2007, 26, 809–817. [Google Scholar] [CrossRef]
- Ahmad, M.; Sayyed, A.H.; Saleem, M.A.; Ahmad, M. Evidence for field evolved resistance to newer insecticides in Spodoptera litura (Lepidoptera: Noctuidae) from Pakistan. Crop Prot. 2008, 27, 1367–1372. [Google Scholar] [CrossRef]
- Peter, C.; David, B.V. Residual toxicity of some insecticides on groundnut to the first and third instar larvae of Spodoptera litura F. (Lepidoptera: Noctuidae). Trop. Pest Manag. 1988, 34, 24–26. [Google Scholar] [CrossRef]
- Zhou, S.Z.; Chen, Z.P.; Xu, Z.F. Effects of three Spodoptera litura control strategies on arthropod diversity and abundance in tobacco agroecosystems in South China. Pak. J. Zool. 2012, 44, 151–157. [Google Scholar]
- Wang, J.J.; Dong, H.G.; Yuan, L.Z. Resistance mechanisms of Spodoptera litura to indoxacarb. J. Plant Protec. 2009, 36, 168–172. [Google Scholar]
- Shad, S.A.; Sayyed, A.H.; Fazal, S.; Saleem, M.A.; Zaka, S.M.; Ali, M. Field evolved resistance to carbamates, organophosphates, pyrethroids, and new chemistry insecticides in Spodoptera litura Fab. (Lepidoptera: Noctuidae). J. Pest Sci. 2012, 85, 153–162. [Google Scholar] [CrossRef]
- Knipling, E.F. Possibilities of insect control or eradication through the use of sexually sterile males. J. Econ. Entomol. 1955, 48, 459–462. [Google Scholar] [CrossRef]
- North, D.T. Inherited sterility in Lepidoptera. Annu. Rev. Entomol. 1975, 20, 167–182. [Google Scholar] [CrossRef]
- Bonomi, A.; Bassetti, F.; Gabrieli, P.; Beadell, J.; Falchetto, M.; Scolari, F.; Gomulski, L.M.; Regazzini, E.; Ouma, J.O.; Caccone, A.; et al. Polyandry is a common event in wild populations of the tsetse fly Glossina fuscipes fuscipes and may impact population reduction measures. PLoS Negl. Trop. Dis. 2011, 5, e1190. [Google Scholar] [CrossRef] [Green Version]
- Gutierrez, A.P.; Ponti, L.; Arias, P.A. Deconstructing the eradication of new world screwworm in North America: Retrospective analysis and climate warming effects. Med. Vet. Entomol. 2019, 33, 282–295. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Drummond, B.A. Multiple mating and sperm competition in the Lepidoptera. In Sperm Competition and the Evolution of Animal Mating Systems; Academic Press: Orlando, FL, USA, 1984; pp. 291–370. [Google Scholar]
- Arnqvist, G.; Nilsson, T. The evolution of polyandry: Multiple mating and female fitness in insects. Anim. Behav. 2000, 60, 145–164. [Google Scholar] [CrossRef]
- Sadek, M.M. Polyandry in field-collected Spodoptera littoralis moths and laboratory assessment of the effects of male mating history. Entomol. Exp. Appl. 2001, 98, 165–172. [Google Scholar] [CrossRef]
- Torres-Vila, L.M.; Rodríguez-Molina, M.C.; Gragera, J.; Bielza-Lino, P. Polyandry in Lepidoptera: A heritable trait in Spodoptera exigua Hübner. Heredity 2001, 86, 177–183. [Google Scholar] [CrossRef] [Green Version]
- Xu, J.; Chen, Z.; Gao, B.; Chen, M.; Fu, D.Y.; Chen, P.; Liu, J.H. Bodyweight-Related Polyandry in the Tobacco Cutworm Moth Spodoptera litura (Lepidoptera: Noctuidae). J. Insect Sci. 2019, 19, 9. [Google Scholar] [CrossRef] [Green Version]
- Seth, R.K.; Khan, Z.; Rao, D.K.; Zarin, M. Flight activity and mating behavior of irradiated Spodoptera litura (Lepidoptera: Noctuidae) males and their F1 progeny for use of inherited sterility in pest management approaches. Fla. Entomol. 2016, 99 (Suppl. 1), 119–130. [Google Scholar] [CrossRef] [Green Version]
- Parker, G.A. Sperm competition and its evolutionary effect on copula duration in the fly Scatophaga stercoraria. J. Insect Physiol. 1970, 16, 1301–1328. [Google Scholar] [CrossRef]
- Horng, S.B.; Plant, R.E. Lek mating system and its impact on male annihilation technique. Res. Popul. Ecol. 1993, 35, 183–197. [Google Scholar] [CrossRef]
- Kraaijeveld, K.; Chapman, T. Effects of male sterility on female remating in the Mediterranean fruitfly, Ceratitis capitata. Proc. R. Soc. Lond. Ser. B Biol. Sci. 2004, 271 (Suppl. 4), S209–S211. [Google Scholar] [CrossRef]
- Simmons, L.W.; Shuker, D.M. Sperm competition. In The Evolution of Insect Mating Systems; Oxford University Press: Oxford, UK, 2014; pp. 181–203. [Google Scholar]
- Etman AA, M.; Hooper, G.H.S. Sperm precedence of the last mating in Spodoptera litura. Ann. Entomol. Soc. Am. 1979, 27, 119–120. [Google Scholar] [CrossRef]
- Seth, R.K.; Rao, D.K.; Reynolds, S.E. Movement of spermatozoa in the reproductive tract of adult male Spodoptera litura: Daily rhythm of sperm descent and the effect of light regime on male reproduction. J. Insect Physiol. 2002, 48, 119–131. [Google Scholar] [CrossRef] [PubMed]
- Gehring, R.D.; Madsen, H.F. Some aspects of the mating and oviposition behavior of the codling moth, Carpocapsa pomonella. J. Econ. Entomol. 1963, 56, 140–143. [Google Scholar] [CrossRef]
- Ouye, M.T.; Graham, H.M.; Richmond, C.A.; Martin, D.F. Mating studies of the pink bollworm. J. Econ. Entomol. 1964, 57, 222–225. [Google Scholar] [CrossRef]
- Angmo, N.; Vimal, N.; Sengupta, M.; Seth, R.K. Effect of male irradiation and its mating status on the remating propensity, insemination quality and reproductive behaviour of male Spodoptera litura (F.). Indian J. Entomol. 2023, 85, 95–102. [Google Scholar]
- Gillott, C. Male accessory gland secretions: Modulators of female reproductive physiology and behavior. Annu. Rev. Entomol. 2003, 48, 163–184. [Google Scholar] [CrossRef] [PubMed]
- Poiani, A. Complexity of seminal fluid: A review. Behav. Ecol. Sociobiol. 2006, 60, 289–310. [Google Scholar] [CrossRef]
- Yu, J.F.; Li, C.; Xu, J.; Liu, J.H.; Ye, H. Male accessory gland secretions modulate female post-mating behavior in the moth Spodoptera litura. J. Insect Behav. 2014, 27, 105–116. [Google Scholar] [CrossRef]
- Obara, Y.; Tateda, H.; Kuwabara, M. Mating Behavior of the Cabbage White Butterfly, Pieris rapae crucivora Boisduval-5-Copulatory Stimuli Inducing Changes of Female Response Patterns. Zool. Mag. 1975, 84, 71–76. [Google Scholar]
- Sugawara, T. Fine structure of the stretch receptor in the bursa copulatrix of the butterfly, Pieris rapae crucivora. Cell Tissue Res. 1981, 217, 23–36. [Google Scholar] [CrossRef]
- Ravi Ram, K.; Wolfner, M.F. Seminal influences: Drosophila Acps and the molecular interplay between males and females during reproduction. Integr. Comp. Biol. 2007, 47, 427–445. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Thibout, E. Stimulation of reproductive activity of females of Acrolepiopsis assectella (Lepidoptera: Hyponomeutoidea) by the presence of eupyrene spermatozoa in the spermatheca. Entomol. Exp. Appl. 1979, 26, 279–290. [Google Scholar] [CrossRef]
- Giebultowicz, J.M.; Raina, A.K.; Uebel, E.C.; Ridgway, R.L. Two-step regulation of sex-pheromone decline in mated gypsy moth females. Arch. Insect Biochem. Physiol. 1991, 16, 95–105. [Google Scholar] [CrossRef]
- Xu, J.; Wang, Q. Seminal fluid reduces female longevity and stimulates egg production and sperm trigger oviposition in a moth. J. Insect Physiol. 2011, 57, 385–390. [Google Scholar] [CrossRef] [PubMed]
- Karube, F.; Kobayashi, M. Presence of eupyrene spermatozoa in vestibulum accelerates oviposition in the silkworm moth, Bombyx mori. J. Insect Physiol. 1999, 45, 947–957. [Google Scholar] [CrossRef]
- He, Y.; Tanaka, T.; Miyata, T. Eupyrene and apyrene sperm and their numerical fluctuations inside the female reproductive tract of the armyworm, Pseudaletia separata. J. Insect Physiol. 1955, 41, 689–694. [Google Scholar] [CrossRef]
- Snow, J.W.; Jones, R.L.; North, D.T.; Holt, G.G. Effects of irradiation on ability of adult male corn earworms to transfer sperm, and field attractiveness of females mated to irradiated males. J. Econ. Entomol. 1972, 65, 906–908. [Google Scholar] [CrossRef]
- Wiklund, C.; Kaitala, A. Sexual selection for large male size in a polyandrous butterfly: The effect of body size on male versus female reproductive success in Pieris napi. Behav. Ecol. 1995, 6, 6–13. [Google Scholar] [CrossRef]
- Cook, P.A.; Wedell, N. Non-fertile sperm delay female remating. Nature 1999, 397, 486. [Google Scholar] [CrossRef]
- Jin, Z.Y.; Gong, H. Male accessory gland derived factors can stimulate oogenesis and enhance oviposition in Helicoverpa armigera (Lepidoptera: Noctuidae). Arch. Insect Biochem. Physiol. 2001, 46, 175–185. [Google Scholar] [CrossRef] [PubMed]
- Kubli, E. Sex-peptides: Seminal peptides of the Drosophila male. Cell. Mol. Life Sci. 2003, 60, 1689–1704. [Google Scholar] [CrossRef] [PubMed]
- Chen, P.S.; Stumm-Zollinger, E.; Aigaki, T.; Balmer, J.; Bienz, M.; Böhlen, P. A male accessory gland peptide that regulates reproductive behavior of female D. melanogaster. Cell 1988, 54, 291–298. [Google Scholar] [CrossRef] [PubMed]
- Yapici, N.; Kim, Y.J.; Ribeiro, C.; Dickson, B.J. A receptor that mediates the post-mating switch in Drosophila reproductive behaviour. Nature 2008, 451, 33–37. [Google Scholar] [CrossRef]
- Hanin, O.; Azrielli, A.; Applebaum, S.W.; Rafaeli, A. Functional impact of silencing the Helicoverpa armigera sex-peptide receptor on female reproductive behaviour. Insect Mol. Biol. 2012, 21, 161–167. [Google Scholar] [CrossRef]
- Li, C.; Yu, J.F.; Lu, Q.; Xu, J.; Liu, J.H.; Ye, H. Molecular characterization and functional analysis of a putative sex-peptide receptor in the tobacco cutworm Spodoptera litura (Fabricius, 1775) (Lepidoptera: Noctuidae). Austral. Entomol. 2014, 53, 424–431. [Google Scholar] [CrossRef]
- Seth, R.K.; Sharma, V.P. Inherited sterility by substerilizing radiation in Spodoptera litura (Lepidoptera: Noctuidae): Bioefficacy and potential for pest suppression. Fla. Entomol. 2001, 83, 183–193. [Google Scholar] [CrossRef]
- Seth, R.K.; Sehgal, S.S. Partial sterilizing radiation dose effect on the F 1 progeny of Spodoptera litura (Fabr.): Growth, bioenergetics and reproductive competence. In Management of Insect Pests: Nuclear and Related Molecular and Genetic Techniques; International Atomic Energy Agency: Vienna, Austria, 1993. [Google Scholar]
- He, F. BCA (bicinchoninic acid) protein assay. Bio-Protocol 2011, 1, e44. [Google Scholar] [CrossRef]
- Sambrook, J.; Russell, D.W. Purification of nucleic acids by extraction with phenol: Chloroform. Cold Spring Harb. Protoc. 2006, 2006, pdb-prot4455. [Google Scholar]
- Livak, K.J.; Schmittgen, T.D. Analysis of relative gene expression data using real-time quantitative PCR and the 2−ΔΔCT method. Methods 2001, 25, 402–408. [Google Scholar] [CrossRef] [PubMed]
- Lu, Q.; Huang, L.Y.; Liu, F.T.; Wang, X.F.; Chen, P.; Xu, J.; Deng, J.Y.; Ye, H. Sex pheromone titre in the glands of Spodoptera litura females: Circadian rhythm and the effects of age and mating. Physiol. Entomol. 2017, 42, 156–162. [Google Scholar] [CrossRef]
- Bonizzoni, M.; Gomulski, L.M.; Bertin, S.; Scolari, F.; Guglielmino, C.R.; Yuval, B.; Gasperi, G.; Malacrida, A.R. Unfaithful Mediterranean fruit fly Ceratitis capitata females: Impact on the SIT? In Area-Wide Control of Insect Pests; Springer: Dordrecht, The Netherlands, 2007; pp. 175–182. [Google Scholar]
- Pérez-Staples, D.; Shelly, T.E.; Yuval, B. Female mating failure and the failure of ‘mating’ in sterile insect programs. Entomol. Exp. Appl. 2013, 146, 66–78. [Google Scholar] [CrossRef] [Green Version]
- Jang, E.B. Physiology of mating behavior in Mediterranean fruit fly (Diptera: Tephritidae): Chemoreception and male accessory gland fluids in female post-mating behavior. Fla. Entomol. 2002, 85, 89–93. [Google Scholar] [CrossRef]
- Vera, M.T.; Wood, R.J.; Cladera, J.L.; Gilburn, A.S. Factors affecting female remating frequency in the Mediterranean fruit fly (Diptera: Tephritidae). Fla. Entomol. 2002, 85, 156–164. [Google Scholar] [CrossRef]
- Parker, A.G.; Vreysen MJ, B.; Bouyer, J.; Calkins, C.O. Sterile insect quality control/assurance. In Sterile Insect Technique: Principles and Practice in Area-Wide Integrated Pest Management; CRC Press: Boca Raton, FL, USA, 2021; pp. 399–440. [Google Scholar]
- Lance, D.R.; McInnis, D.O.; Rendon, P.; Jackson, C.G. Courtship among sterile and wild Ceratitis capitata (Diptera: Tephritidae) in field cages in Hawaii and Guatemala. Ann. Entomol. Soc. Am. 2000, 93, 1179–1185. [Google Scholar] [CrossRef]
- Holt, G.G.; North, D.T. Effects of gamma irradiation on the mechanisms of sperm transfer in Trichoplusia ni. J. Insect Physiol. 1970, 16, 2211–2222. [Google Scholar] [CrossRef]
- Koudelová, J.; Cook, P.A. Effect of gamma radiation and sex-linked recessive lethal mutations on sperm transfer in Ephestia kuehniella (Lepidoptera: Pyralidae). Fla. Entomol. 2001, 84, 172–182. [Google Scholar] [CrossRef]
- Sims, S.R. Aspects of mating frequency and reproductive maturity in Papilio zelicaon. Am. Midl. Nat. 1979, 102, 36–50. [Google Scholar] [CrossRef]
- Vera, M.T.; Cladera, J.L.; Calcagno, G.; Vilardi, J.C.; Mcinnis, D.W. Remating of wild Ceratitis capitata (Diptera: Tephritidae) females in field cages. Ann. Entomol. Soc. Am. 2003, 96, 563–570. [Google Scholar] [CrossRef] [Green Version]
- Chapman, R.F. The Insects Structure and Function; American Elsevier Publishing Company: New York, NY, USA, 1969. [Google Scholar]
- Leopold, R.A. The role of male accessory glands in insect reproduction. Annu. Rev. Entomol. 1976, 21, 199–221. [Google Scholar] [CrossRef]
- Seth, R.K.; Rao, D.K.; Kaur, J.J. Developmental pattern of testes in F 1 progeny of gamma irradiated Spodoptera litura (Fabr.). J. Nucl. Agric. Biol. 2000, 29, 129–141. [Google Scholar]
- Cook, P.A.; Wedell, N. Ejaculate dynamics in butterflies: A strategy for maximizing fertilization success. Proc. R. Soc. Lond. Ser. B Biol. Sci. 1996, 263, 1047–1051. [Google Scholar]
- Strong, F.E.; Sheldahl, J.A.; Hughes, P.R.; Hussein EM, K. Reproductive biology of Lygus hesperus Knight. Hilgardia 1970, 40, 105–147. [Google Scholar] [CrossRef] [Green Version]
- Brent, C.S. Reproductive refractoriness in the western tarnished plant bug (Hemiptera: Miridae). Ann. Entomol. Soc. Am. 2010, 103, 300–306. [Google Scholar] [CrossRef] [Green Version]
- Sugawara, T. Stretch reception in the bursa copulatrix of the butterfly, Pieris rapae crucivora, and its role in behaviour. J. Comp. Physiol. 1979, 130, 191–199. [Google Scholar] [CrossRef]
- Proshold, F.I.; Mastro, V.C.; Bernon, G.L. Sperm transfer by gypsy moths (Lepidoptera: Lymantriidae) from irradiated males: Implication for control by inherited sterility. J. Econ. Entomol. 1993, 86, 1104–1108. [Google Scholar] [CrossRef]
- Taylor, J.R.; Orley, R. Relationship of multiple mating to fertility in Atteva punctella (Lepidoptera: Yponomeutidae). Ann. Entomol. Soc. Am. 1967, 60, 583–590. [Google Scholar] [CrossRef]
- Calkins, C.O.; Parker, A.G. Sterile insect quality. In Sterile insect technique (pp. 269-296). Springer, Dordrechtction. J. Insect Physiol. 2005, 48, 119–131. [Google Scholar]
- Radhakrishnan, P.; Taylor, P.W. Ability of male Queensland fruit flies to inhibit receptivity in multiple mates, and the associated recovery of accessory glands. J. Insect Physiol. 2008, 54, 421–428. [Google Scholar] [CrossRef]
- Wu, T.; Cao, D.H.; Liu, Y.; Yu, H.; Fu, D.Y.; Ye, H.; Xu, J. Mating-Induced Common and Sex-Specific Behavioral, Transcriptional Changes in the Moth Fall Armyworm (Spodoptera frugiperda, Noctuidae, Lepidoptera) in Laboratory. Insects 2023, 14, 209. [Google Scholar] [CrossRef] [PubMed]
- Rafaeli, A. Neuroendocrine control of pheromone biosynthesis in moths. Int. Rev. Cytol. 2002, 213, 49–91. [Google Scholar] [PubMed]
- Xu, J.; Wang, Q.; He, X.Z. Emergence and reproductive rhythms of Ephestia kuehniella (Lepidoptera: Pyralidae). N. Z. Plant Prot. 2008, 61, 277–282. [Google Scholar] [CrossRef] [Green Version]
- Raina, A.K. Male-induced termination of sex pheromone production and receptivity in mated females of Heliothis zea. J. Insect Physiol. 1989, 35, 821–836. [Google Scholar] [CrossRef]
- Kingan, T.G.; Bodnar, W.M.; Raina, A.K.; Shabanowitz, J.; Hunt, D.F. The loss of female sex pheromone after mating in the corn earworm moth Helicoverpa zea: Identification of a male pheromonostatic peptide. Proc. Natl. Acad. Sci. USA 1995, 92, 5082–5086. [Google Scholar] [CrossRef] [PubMed]
- Xu, J.; Shi, M.R.; Fu, D.Y.; Yu, H.; Chen, P.; Ye, H. Male accessory gland factors increased sex pheromone titres in the glands of Spodoptera litura female moths by inhibiting calling behavior. bioRxiv 2019, 2019, 872150. [Google Scholar]
- Mossinson, S.; Yuval, B. Regulation of sexual receptivity of female Mediterranean fruit flies: Old hypotheses revisited and a new synthesis proposed. J. Insect Physiol. 2003, 49, 561–567. [Google Scholar] [CrossRef] [PubMed]
- Proshold, F.; Bernon, G.L. Multiple mating in laboratory-reared gypsy moths (Lepidoptera: Lymantriidae). J. Econ. Entomol. 1994, 87, 661–666. [Google Scholar] [CrossRef]
- Saour, G.; Makee, H. Effect of gamma irradiation on sperm utilization in twice-mated female Phthorimaea operculella Zeller (Lep., Gelechiidae). J. Appl. Entomol. 1999, 123, 513–517. [Google Scholar] [CrossRef]
- Landeta-Escamilla, A.; Hernández, E.; Arredondo, J.; Díaz-Fleischer, F.; Pérez-Staples, D. Male irradiation affects female remating behavior in Anastrepha serpentina (Diptera: Tephritidae). J. Insect Physiol. 2016, 85, 17–22. [Google Scholar] [CrossRef] [PubMed]
- Shelly, T. Ability of sterile males to inhibit female remating in the melon fly Zeugodacus cucurbitae (Diptera: Tephritidae). Fla. Entomol. 2019, 102, 278–280. [Google Scholar]
- Abraham, S.; Liendo, M.C.; Devescovi, F.; Peralta, P.A.; Yusef, V.; Ruiz, J.; Cladera, J.L.; Vera, M.T.; Segura, D.F. Remating behavior in Anastrepha fraterculus (Diptera: Tephritidae) females is affected by male juvenile hormone analog treatment but not by male sterilization. Bull. Entomol. Res. 2013, 103, 310–317. [Google Scholar] [CrossRef]
- Arredondo, J.; Tejeda, M.T.; Ruiz, L.; Meza, J.S.; Pérez-Staples, D. Timing of irradiation and male mating history effects on female remating in Anastrepha ludens (Diptera: Tephritidae). Fla. Entomol. 2017, 100, 566–570. [Google Scholar] [CrossRef] [Green Version]
- Krüger, A.P.; Schlesener, D.C.; Martins, L.N.; Wollmann, J.; Deprá, M.; Garcia, F.R. Radiation effects on Drosophila suzukii (Diptera: Drosophilidae) reproductive behaviour. J. Appl. Entomol. 2019, 143, 88–94. [Google Scholar] [CrossRef]
- Ocampo, V.R. Effect of a substerilizing dose of radiation on the mating competitiveness of male and on the mating propensity of female Helicoverpa armigera (Lepidoptera: Noctuidae). Fla. Entomol. 2001, 84, 194–198. [Google Scholar] [CrossRef]
- Eberhard, W. Female Control: Sexual Selection by Cryptic Female Choice; Princeton University Press: Princeton, NJ, USA, 1996; Volume 69. [Google Scholar]
- Thornhill, R.; Alcock, J. The evolution of insect mating systems. In The Evolution of Insect Mating Systems; Harvard University Press: Cambridge, MA, USA, 2013. [Google Scholar]
- Harvey, G.T. Mean weight and rearing performance of successive egg clusters of eastern spruce budworm (Lepidoptera: Tortricidae). Can. Entomol. 1977, 109, 487–496. [Google Scholar] [CrossRef]
- Suzuki, Y. Adult longevity and reproductive potential of the small cabbage white, Pieris rapae crucivora Boisduval (Lepidoptera: Pieridae). Appl. Entomol. Zool. 1978, 13, 312–313. [Google Scholar] [CrossRef] [Green Version]
- Laturney, M.; van Eijk, R.; Billeter, J.C. Last male sperm precedence is modulated by female remating rate in Drosophila melanogaster. Evol. Lett. 2018, 2, 180–189. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hook, K.A. Female remating decisions and a shorter inter-mating interval diminish last-male sperm precedence. Behav. Ecol. Sociobiol. 2017, 71, 121. [Google Scholar] [CrossRef]
- Birkhead, T.R.; Møller, A.P. Numbers and size of sperm storage tubules and the duration of sperm storage in birds: A comparative study. Biol. J. Linn. Soc. 1992, 45, 363–372. [Google Scholar] [CrossRef]
- Lessells, C.M.; Birkhead, T.R. Mechanisms of sperm competition in birds: Mathematical models. Behav. Ecol. Sociobiol. 1990, 27, 325–337. [Google Scholar] [CrossRef]
- Birkhead, T.R.; Biggins, J.D. Sperm competition mechanisms in birds: Models and data. Behav. Ecol. 1998, 9, 253–260. [Google Scholar] [CrossRef] [Green Version]
- Simmons, L.W. The evolution of polyandry: An examination of the genetic incompatibility and good-sperm hypotheses. J. Evol. Biol. 2001, 14, 585–594. [Google Scholar] [CrossRef]
- Manier, M.K.; Belote, J.M.; Berben, K.S.; Lüpold, S.; Ala-Honkola, O.; Collins, W.F.; Pitnick, S. Rapid diversification of sperm precedence traits and processes among three sibling Drosophila species. Evolution 2013, 67, 2348–2362. [Google Scholar] [CrossRef] [Green Version]
- Seth, R.K.; Kaur, J.J.; Rao, D.K.; Reynolds, S.E. Sperm transfer during mating, movement of sperm in the female reproductive tract, and sperm precedence in the common cutworm Spodoptera litura. Physiol. Entomol. 2002, 27, 1–14. [Google Scholar] [CrossRef]




Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Angmo, N.; Sengupta, M.; Vimal, N.; Seth, R.K. Receptivity and Remating Propensity in Female Spodoptera litura (Fabricius) after Mating with an Irradiated Male or Its F1 Male Progeny. Insects 2023, 14, 651. https://doi.org/10.3390/insects14070651
Angmo N, Sengupta M, Vimal N, Seth RK. Receptivity and Remating Propensity in Female Spodoptera litura (Fabricius) after Mating with an Irradiated Male or Its F1 Male Progeny. Insects. 2023; 14(7):651. https://doi.org/10.3390/insects14070651
Chicago/Turabian StyleAngmo, Nilza, Madhumita Sengupta, Neha Vimal, and Rakesh Kumar Seth. 2023. "Receptivity and Remating Propensity in Female Spodoptera litura (Fabricius) after Mating with an Irradiated Male or Its F1 Male Progeny" Insects 14, no. 7: 651. https://doi.org/10.3390/insects14070651