
Development of a Diet Production System for Conopomorpha cramerella (Lepidoptera: Gracillariidae), a Major Cocoa Production Pest in Southeast Asia and the Pacific Islands
 , and
, and
Abstract
:Simple Summary
Abstract
1. Introduction
2. Materials and Methods
2.1. Insect Collection and Laboratory Site Descriptions
2.2. Conopomorpha Cramerella Laboratory Rearing and Diet Development
2.2.1. Eggs Collection
2.2.2. Egg Decontamination
2.3. Artificial Diet Materials and Methods
2.3.1. Diets Evaluated
2.3.2. MM1
2.3.3. MM4
2.3.4. pH and Water Activity
2.4. Conopomorpha Cramerella Development on the Artificial Diets
2.4.1. Egg Hatching
2.4.2. Larval Developmental Success
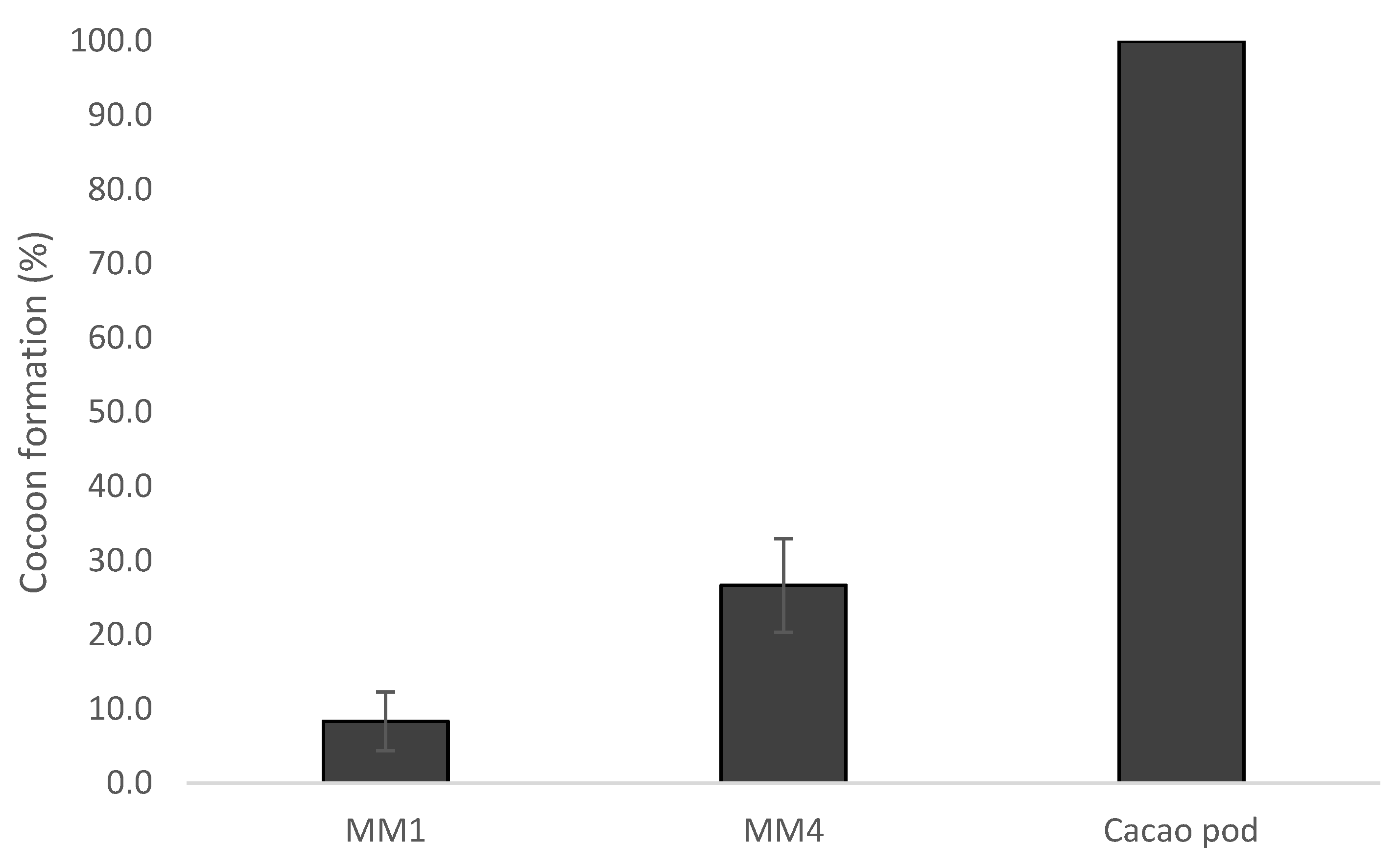
2.4.3. Cocoon Formation Success
2.4.4. Pupation Success
2.4.5. Adult Emergence
2.4.6. Adult Survival
2.4.7. Sex Ratio
2.5. Statistical Analysis
3. Results and Discussion
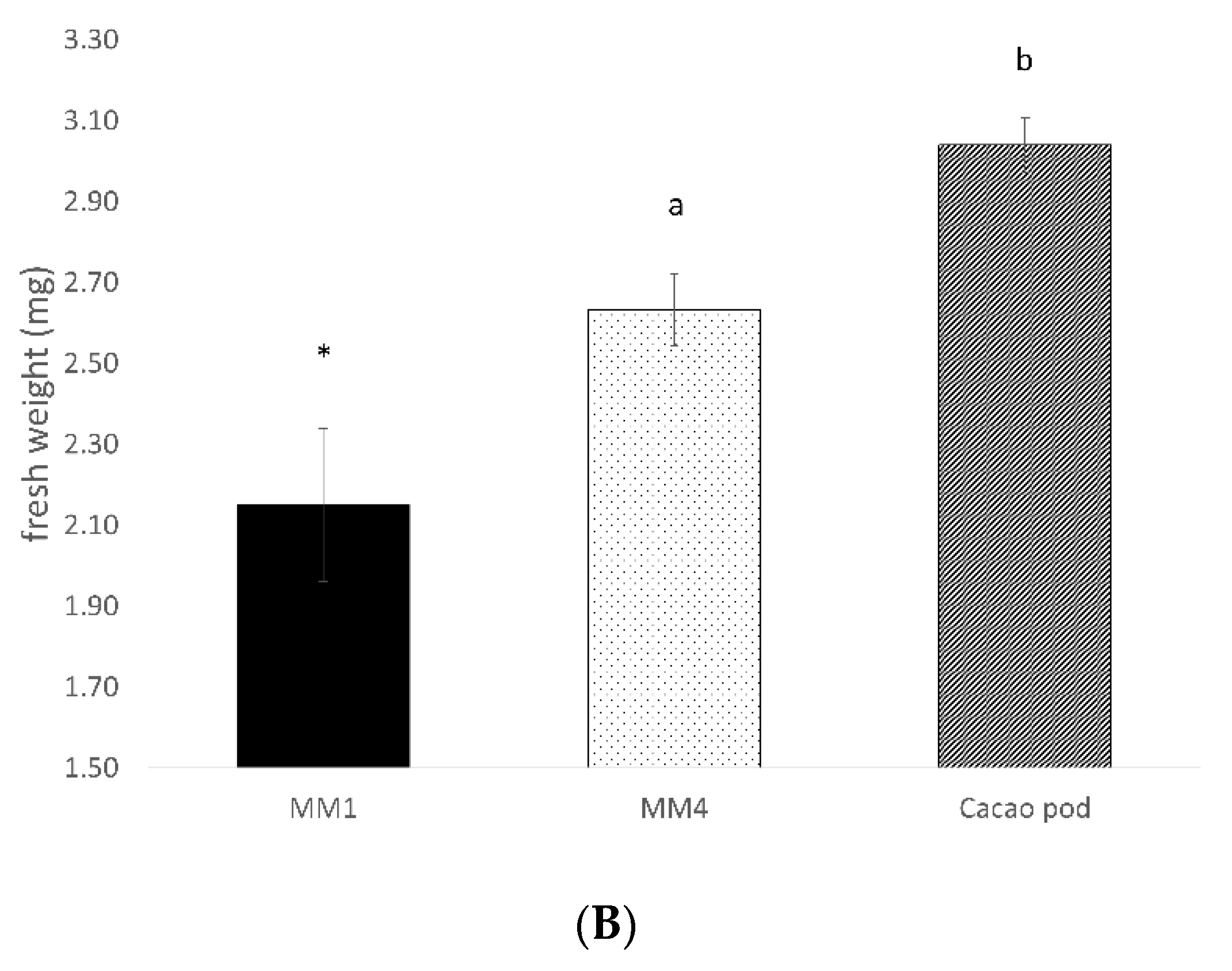
3.1. Diet Development
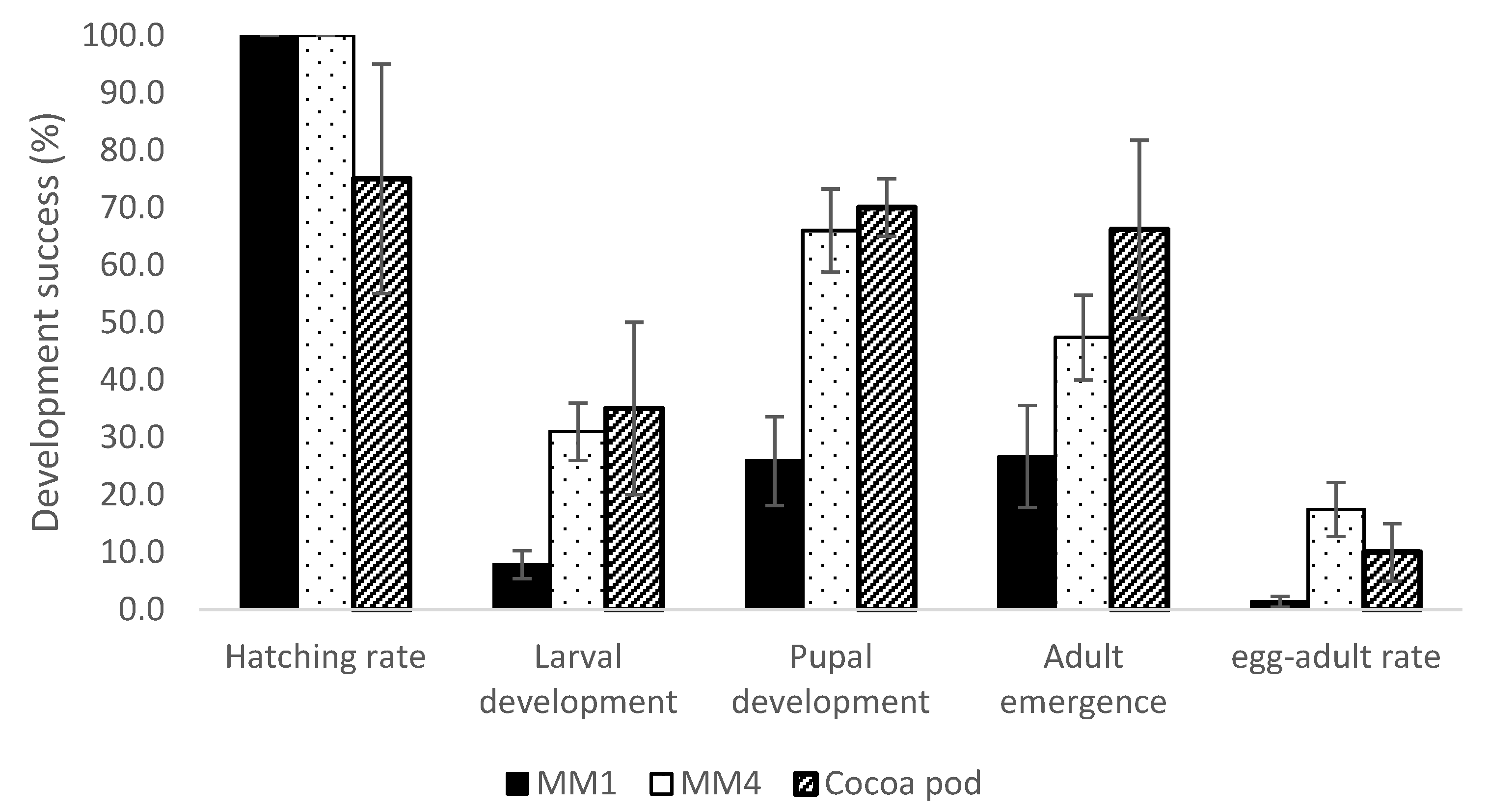
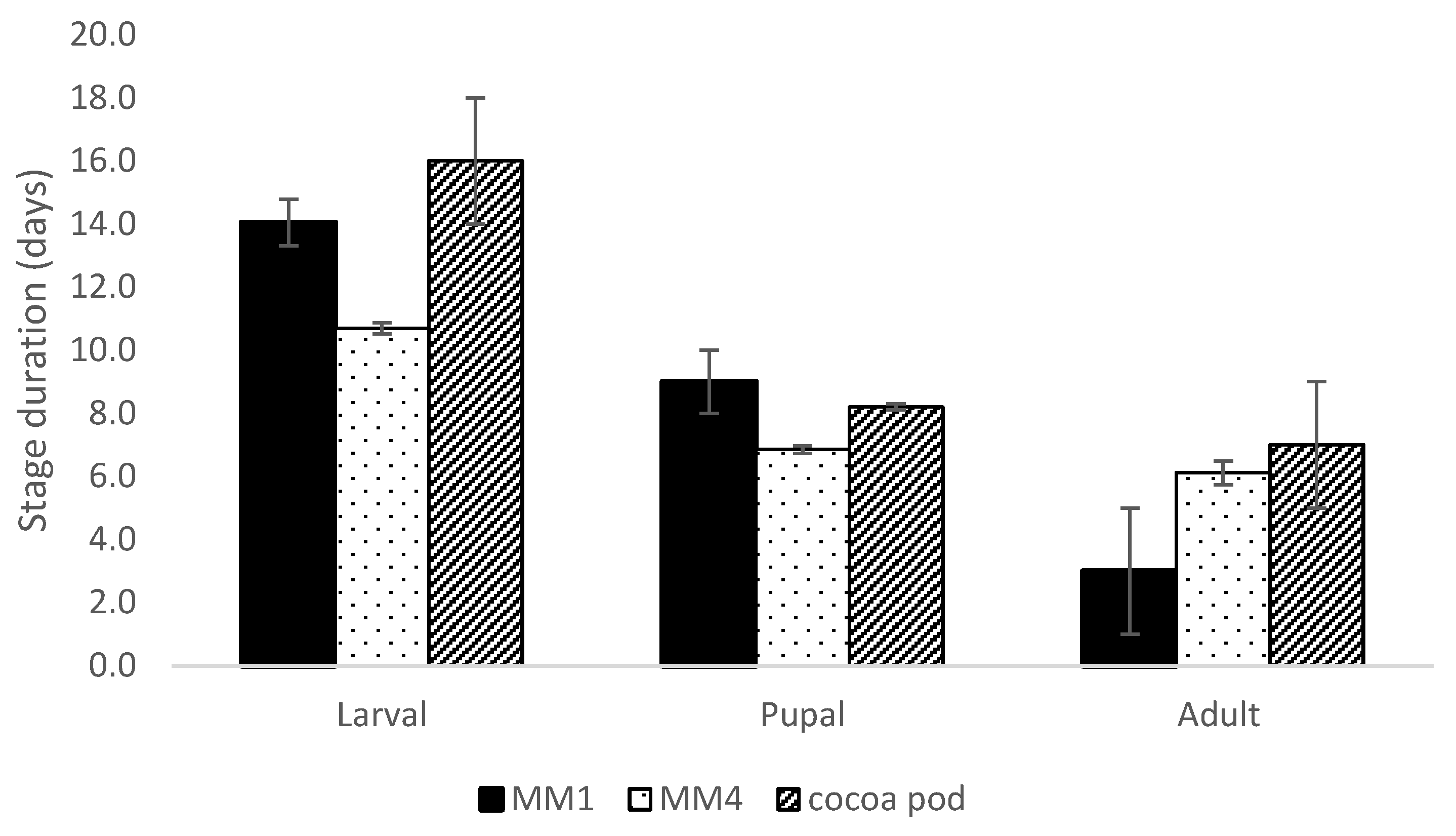
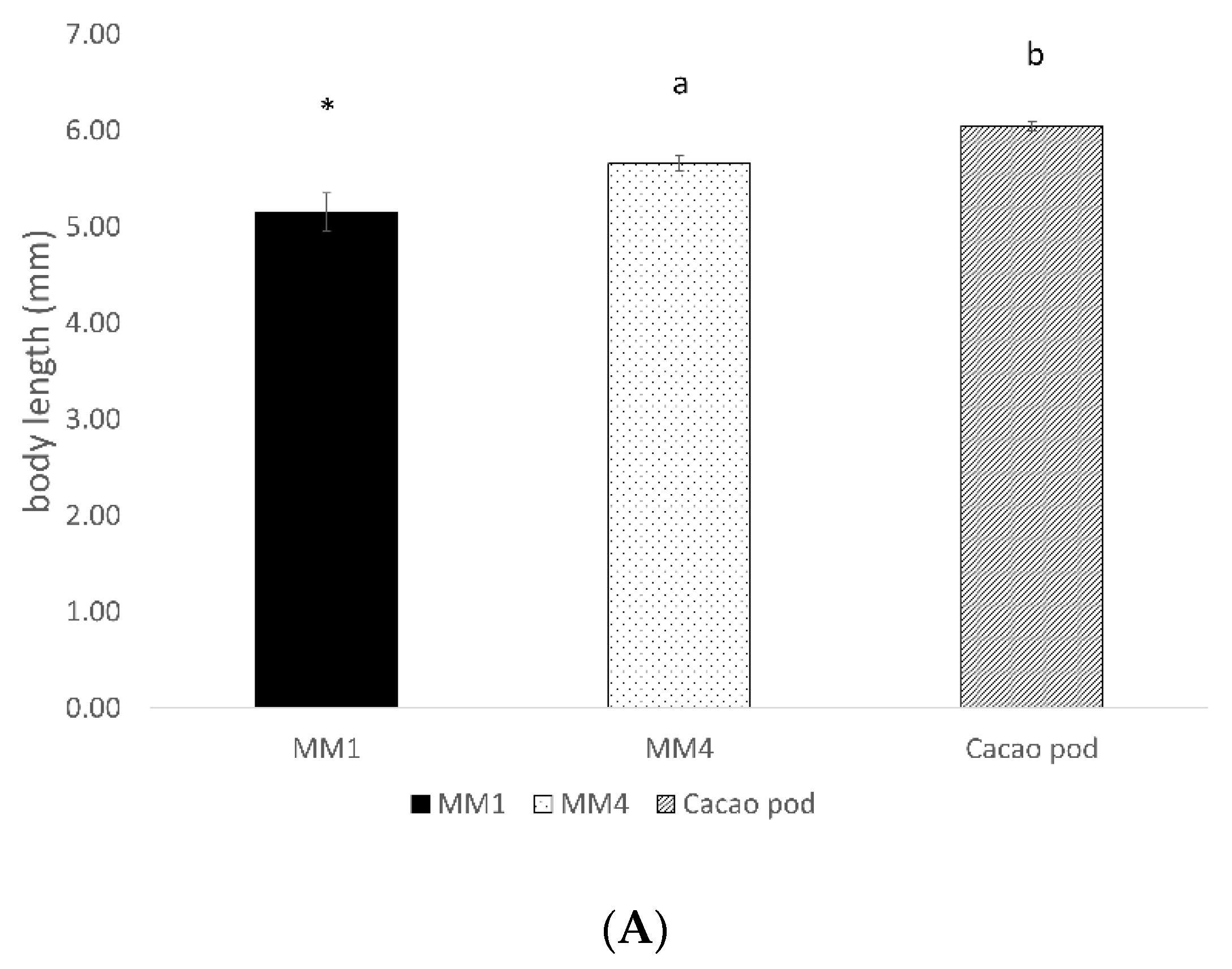
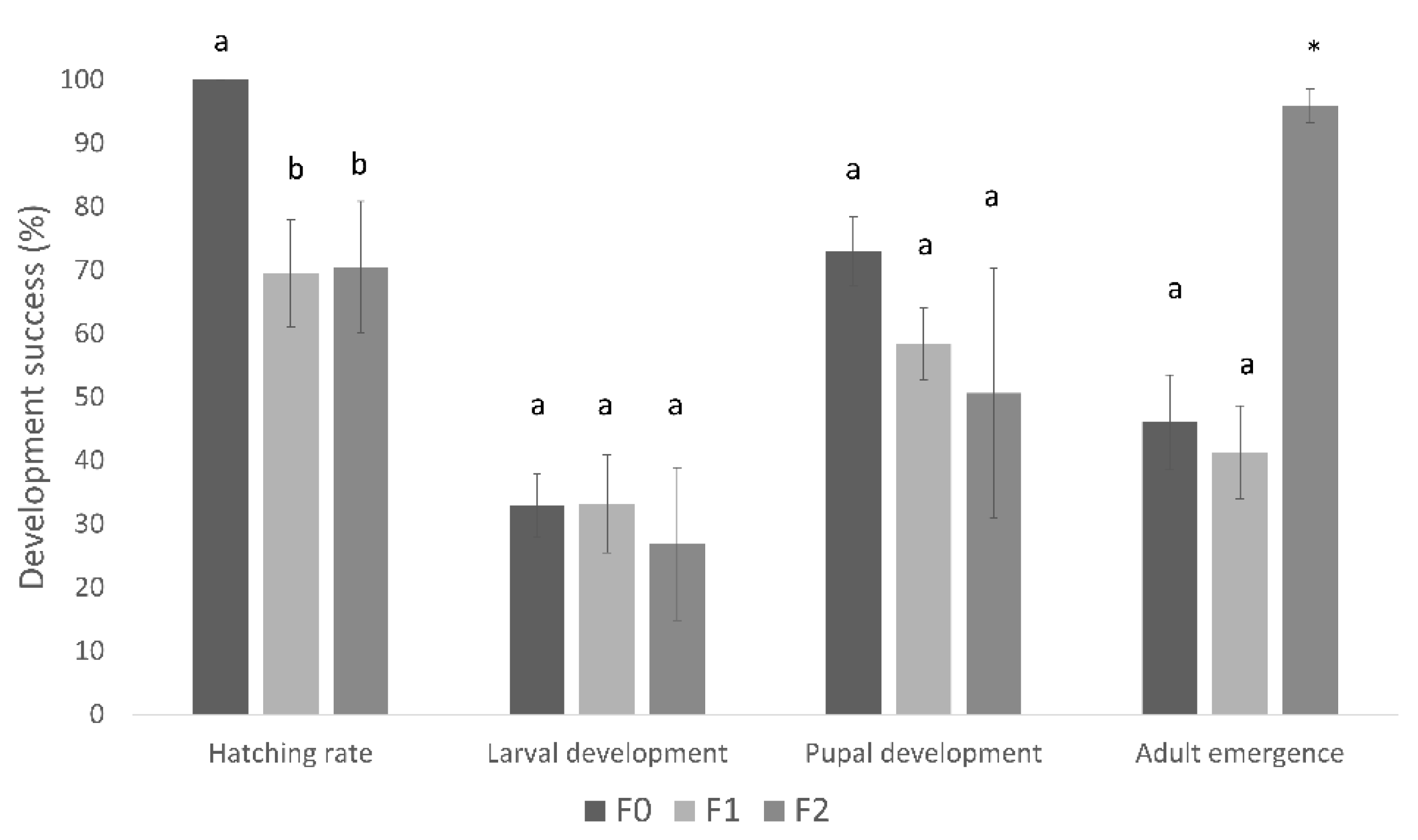
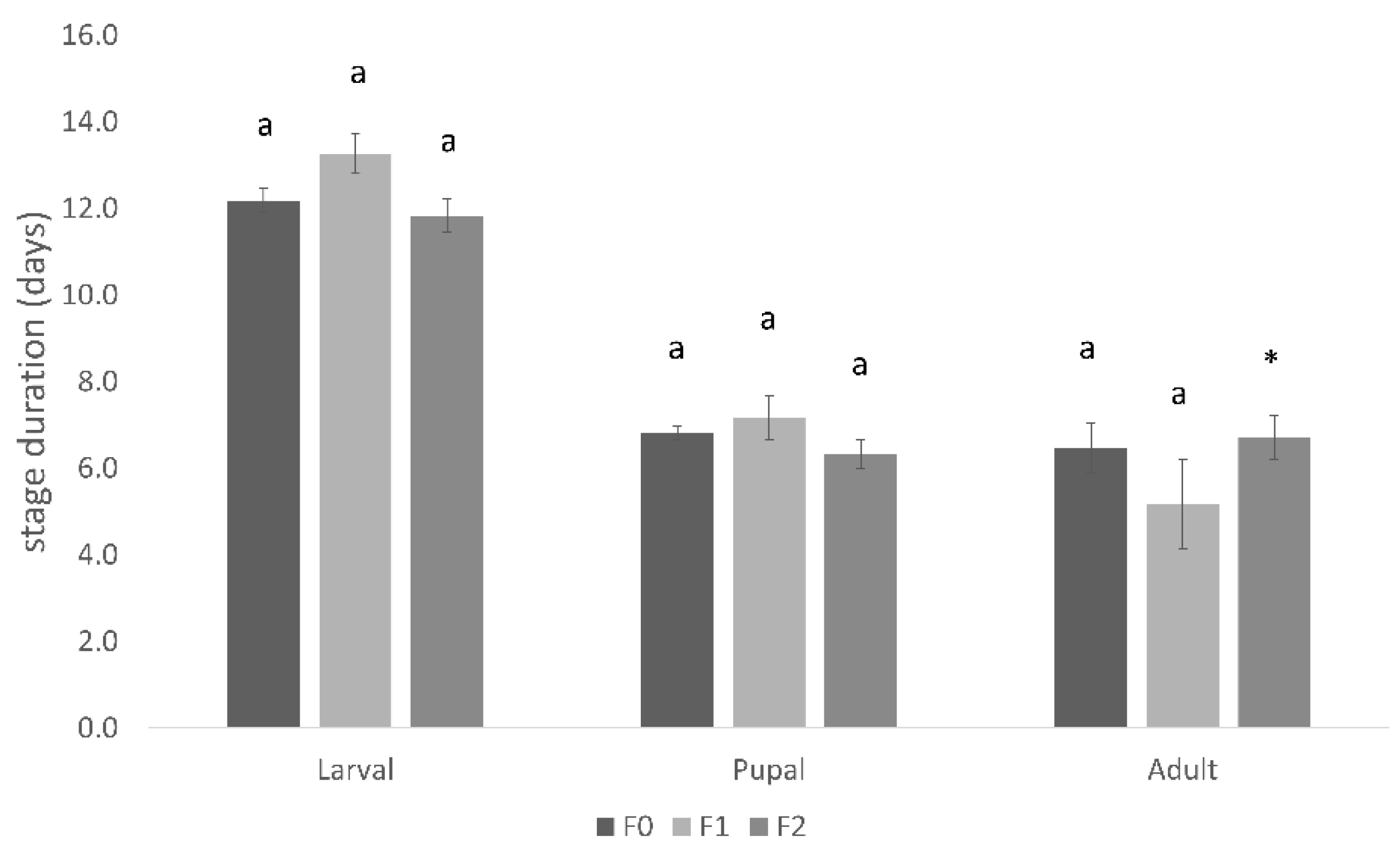
3.2. Conopomorpha Cramerella Development on the Artificial Diets
Author Contributions
Funding
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Niego, A.G.T.; Lambert, C.; Mortimer, P.; Thonklang, N.; Rapio, S.; Grosse, M.; Schrey, H.; Charria-Giron, E.; Walker, A.; Hyde, K.D.; et al. The contribution of fungi to the global economy. Fungal Divers. 2023, 1–43. [Google Scholar] [CrossRef]
- Posada, F.J.; Virdiana, I.; Navies, M.; Pava-Ripoll, M.; Hebbar, P. Sexual dimorphism of pupae and adults of the cocoa pod borer, Conopomorpha cramerella. J. Insect Sci. 2011, 11, 52. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- De Prins, J.; De Prins, W. Global Taxonomic Database of Gracillariidae (Lepidoptera). August 2018, Volume 7. Available online: https://www.gracillariidae.net/ (accessed on 20 May 2022).
- Azhar, I.; Long, G.E. Effect of cocoa pod age on egg distribution and egg parasitism of the cocoa pod borer in Malaysia. Entomol. Exp. Et Appl. 1996, 81, 81–89. [Google Scholar] [CrossRef]
- Posada, F.; Vega, F.E. Establishment of the fungal entomopathogen Beauveria bassiana (Ascomycota: Hypocreales) as an endophyte in cocoa seedling (Theobroma cacao). Mycologia 2005, 97, 1195–1200. [Google Scholar] [CrossRef] [PubMed]
- Lim, G.T. Biology, ecology and control of cocoa pod borer Conopomorpha cramerella (Snellen). In Cocoa Pest and Disease Management in Southeast Asia and Australasia; Keane, P.J., Putter, C.A.J., Eds.; Food and Agriculture Organization of the United Nations: Rome, Italy, 1992; pp. 85–100. [Google Scholar]
- Valenzuela, I.; Bin Purung, H.; Roush, R.T.; Hamilton, A.J. Practical yield loss models for infestation of cocoa with cocoa pod borer moth, Conopomorpha cramerella (Snellen). Crop Prot. 2014, 66, 19–28. [Google Scholar] [CrossRef]
- Day, R.K. Effect of cocoa pod borer, Conopomorpha cramerella, on cocoa yield and quality in Sabah, Malaysia. Crop Prot. 1986, 8, 332–339. [Google Scholar] [CrossRef]
- Bradley, J.D. Identity of the South-East Asian cocoa moth Conopomorpha cramerella (Snellen) (Lepidoptera: Gracillariidae) with descriptions of three allied new species. Bull. Ent. Res. 1986, 76, 41–51. [Google Scholar] [CrossRef]
- Alias, A.; Lamin, K.; Lee, M.T. The use of the egg parasitoid Trichogrammatoidea bactrae Fumata Nagaraja, for the control of the cocoa pod borer Conopomorpha cramerella Snellen: Research progress, constraints and prospects. In Biological Control in the Tropics; Hong, L.W., Sastroutomo, S.S., Caunter, I.G., Ali, J., Yeang, L.K., Vijaysegaran, S., Sen, Y.H., Eds.; CAB International: Sengalor, Malaysia, 1999; pp. 48–51. [Google Scholar]
- Lee, C.H. Planting Cocoa—Challenges and Realities. Malaysian Cocoa Board. 2013, pp. 1–19. Available online: https://www.scribd.com/document/341162814/PLANTING-COCOA-CHALLENGES-AND-REALITY-IN-MALAYSIA-pdf (accessed on 25 May 2023).
- Neilson, J. Global markets, farmers and the state: Sustaining profits in the Indonesian cocoa sector. Bull. Indones. Econ. Stud. 2007, 43, 227–250. [Google Scholar] [CrossRef]
- Leppla, N.; Davis, F.; Schneider, J. Introduction. In Principles and Procedures for Rearing High Quality Insects; Schneider, J., Ed.; Mississippi State University: Mississippi, MS, USA, 2009; pp. 3–8. [Google Scholar]
- Marelli, J.-P. Welcome message. In Increasing the Resilience of Cacao to Major Pest and Disease Threats in the 21st Century; Proceedings of an Asia-Pacific Regional Cocoa Ipm Symposium, Aciar Proceedings Series, No. 149; Niogret, J., Sanchez, V., Marelli, J.-P., Eds.; Australian Centre for International Agricultural Research: Canberra, Australia, 2020; p. 10. [Google Scholar]
- Vreysen, M.J.; Klassen, W.; Carpenter, J.E. Overview of technological advances toward greater efficiency and efficacy in sterile insect-inherited sterility programs against moth pests. Fla. Ent. 2016, 99, 1–13. [Google Scholar] [CrossRef] [Green Version]
- Furtek, D.; Kasran, R.; Chia, T.; Jonchiul, L.; Mohammed, A.; Hartney, V. Artificial diet for cocoa pod borer (Conopomorpha cramerella Snellen): Biotechnology research by the Malaysian cocoa board. In Proceeding of the International Workshop on New Technologies and Cocoa Breeding; Malaysian Cocoa Board: Kota Kinabalu, Malaysia, 2001; pp. 153–154. [Google Scholar]
- Santoso, D.; Chaidamsari, T.; Wiryadiputra, S.; de Maagd, R.A. Activity of Bacillus thuringiensis toxins against cocoa pod borer larvae. Pest Manag. Sci. 2004, 60, 733–736. [Google Scholar] [CrossRef] [PubMed]
- Awang, A.; Karim, R.; Furtek, D.; Board, M.C. Artificial diet for cocoa pod borer moth, Conopomorpha cramerella Snellen. In 4th Malaysian International Cocoa Conference; Malaysian Cocoa Board: Kuala Lumpur, Malaysia, 2006; pp. 293–300. [Google Scholar]
- Cohen, A.C. Insect Diets: Science and Technology; CRC Press: Boca Raton, FL, USA, 2015. [Google Scholar]
- Cohen, A.C. Design, Operation and Control of Insect Rearing Systems: Science, Technology and Infrastructure; CRC Press: Boca Raton, FL, USA, 2021; ISBN 9780203702956. [Google Scholar]
- Rockland, L.B.; Nishi, S.K. Fundamentals of water activity. Food. Technol. 1980, 34, 42–59. [Google Scholar]
- Ngomane, N.C.; Pieterse, E.; Woods, M.J.; Conlong, D.E. Formulation of Artificial Diets for Mass-Rearing Eldana saccharina Walker (Lepidoptera: Pyralidae) Using the Carcass Milling Technique. Insects 2022, 13, 316. [Google Scholar] [CrossRef] [PubMed]
- Chambers, D.L.; Ashely, E.G. Putting quality control in insect rearing. In Advances and Challenges in Insect Rearing; King, E.G., Leppla, N.C., Eds.; Agricultural Research Service: New Orleans, LA, USA, 1984; pp. 256–260. [Google Scholar]
- Agoston, J.; Tóth-Horgosi, P.; Kiss, T.; Taskovics, Z.T. Assessment of nutrient content of tomato hybrids for processing. Acta Univ. Sapentiae-Agric. Environ. 2017, 9, 63–69. [Google Scholar] [CrossRef] [Green Version]
- Catherall, J.; Maree, A. Yield, pH and Sugar Content of Tomato Varieties Grown in Unheated Greenhouses in the North Cariboo; Cariboo Agricultural Research Alliance; Climate Action Initiative, BC Agriculture and Food: Victoria, BC, Canada, 2016. [Google Scholar]
- Day, R.K. Control of the Cocoa Pod Borer (Conopomorpha cramerella). Ph.D. Thesis, University of London, London, UK, 1985. [Google Scholar]
- Wardojo, S. The cocoa pod borer–A major hindrance to cocoa development. Indones. Agric. Res. Dev. J. 1980, 2, 1–9. [Google Scholar]
- Vanialingam, T.; Easaw, P.T.; Irshad, M.; Manals, J.; Cruz, P. Early work in the control of the cocoa pod borer Acrocercops cramerella in the Philippines. In Proceedings of the Eight International Cocoa Research Conference, Cartagena, Colombia, 18–23 October 1981; Cocoa Producers Alliance: Lagos, Nigeria, 1981; pp. 345–351. [Google Scholar]
- Wessel-Riemens, P.C. The cocoa moth pod borer (Acrocercops cramerella Sn.). Review and Translation of Research in Indonesia. ICCO Cocoa Research Policy Committee. 1981, pp. 1900–1918. Available online: chrome-extension://efaidnbmnnnibpcajpcglclefindmkaj/https://edepot.wur.nl/468165 (accessed on 25 May 2023).
- McMahon, P.; Iswanto, A.; Susilo, A.W.; Sulistyowati, E.; Wahab, A.; Imron, M.; Purwantara, A.; Mufrihati, E.; Dewi, V.S.; Lambert, S.; et al. On-farm selection for quality and resistance to pest/diseases of cocoa in Sulawesi: (i) performance of selections against cocoa pod borer, Conopomorpha cramerella. Int. J. Pest Manag. 2009, 55, 325–337. [Google Scholar] [CrossRef]
- Niogret, J.; Ekayanti, A.; Ingram, K.; Lambert, S.; Kendra, P.E.; Alborn, H.; Epsky, N.D. Development and behavioral ecology of Conopomorpha cramerella (Lepidoptera: Gracillariidae). Fla. Ent. 2019, 102, 382–387. [Google Scholar] [CrossRef] [Green Version]








{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Group | Ingredients | Source | Unit | Diet MM1 | Diet MM4 |
|---|---|---|---|---|---|
| A | Fresh tomato | Tomoni Local Fresh Market, Indonesia | g (%) | 500 (40.0) | - |
| Canned tomato | CIRIO 1856, Pelati, Peeled plum Tomatoes, Savena, Italy | g (%) | - | 500 (42.8) | |
| Fresh egg yolk | Tomoni Local Fresh Market, Indonesia | g (%) | 100 (8.0) | 50 (4.3) | |
| B | Tapioca flour | Papioka, Real Organic Tapioca (Gluten Free), Pati, Indonesia | g (%) | 15 (1.2) | 15 (1.3) |
| Lecithin granules | NOW, Non-GMO Lecithin Granules, IL, USA | g (%) | 15 (1.2) | 15 (1.3) | |
| Cellulose | Sigma Aldrich, Beijing, China | g (%) | 30 (2.4) | 30 (2.6) | |
| Brewer’s Yeast | Sigma Aldrich, Sao Paulo, Brazil. | g (%) | 15 (1.2) | 3.75 (0.3) | |
| Chickpea flour | Bob’s Red Mill, Stone Ground, Non-GMO Chickpea flour, Oregon, USA | g (%) | - | 7.5 (0.6) | |
| Red kidney bean flour | Lingkar Organik, Tepung Kacang Merah Organik, Yogyakarta, Indonesia | g (%) | - | 7.5 (0.6) | |
| C | Myo-inositol | Duchefa Biochemie, BH Haarlem, The Netherlands | g (%) | 1 (0.1) | 1 (0.1) |
| Choline chloride | Sigma Aldrich, Beijing, China | g (%) | 1 (0.1) | 1 (0.1) | |
| Vitamin fortification mix for Lepidoptera | Frontier Scientific, Newark, DE, USA | g (%) | - | 0.85 (0.1) | |
| White Sugar | Gulaku, Lampung, Indonesia | g (%) | - | 3.3 (0.3) | |
| Sterile RO water | Onelab Waterone, Deionized water, double RO, EDI, ultrafilter, UV. Jawa Timur, Indonesia water, | mL (%) | 20 (1.6) | 50 (4.3) | |
| D | Flaxseed oil | Nature’s Gift, Omega Gold, Sabah, Malaysia | mL (%) | 40 (3.2) | |
| Linseed oil | Winsor and Newton, London, UK | mL (%) | 10 (0.9) | ||
| E | Phytagel | Sigma Aldrich, Beijing, China | g (%) | 30 (2.4) | 20 (1.7) |
| Sterile RO water | As above | mL (%) | 380 (30.4) | 450 (38.5) | |
| F | Ascorbic acid | Swanson, Pure Vitamin C powder, ND, USA. | g (%) | 1.7 (0.1) | 0.85 (0.1) |
| Sterile RO water | As above | mL (%) | 100 (8.0) | - | |
| G | Antimicrobial blend * | mL (%) | - | 2.5 (0.2) | |
| Total Diet Volume | mL (%) | 1248.7 (100) | 1168.25 (100) |
| MM1 | MM4 | Cocoa | |
|---|---|---|---|
| pH | 5.7 | 4.8 | 5.04 |
| Water content (%) | 80.2 | 84.5 | 88.3 |
| Water activity | 0.988 | 0.985 | |
| Ash (%) | 3.9 | 4.3 | 5.8 |
| Fat (%) | 35.9 | 16.8 | 0.9 |
| Protein | 14.1 | 12.6 | 8.2 |
| Total Carbohydrates | 46.5 | 66.5 | 84.6 |
| Total Dietary Fiber | 32.7 | 57.4 | |
| Total Sterols | 83.8 | 66.5 | 78.5 |
| Saturated Fatty Acids | 6.1 | 3.6 | * |
| Total Cis Unsaturated Fatty Acids | 24.9 | 12.1 | * |
| Monounsaturated Fatty Acids | 9.0 | 5.0 | * |
| Polyunsaturated Fatty Acids | 15.9 | 7.0 | * |
| Trans Fatty Acids | 0.0 | 0.0 | * |
| Omega 3 Fatty Acids | 10.7 | 2.6 | * |
| Omega 6 Fatty Acids | 5.9 | 4.7 | * |
| Omega 9 Fatty Acids | 8.4 | 4.7 | * |
| Total Fatty Acids | 32.4 | 16.3 | * |
| 16:0 Palmitic | 4.7 | 2.9 | * |
| 16:1 Palmitoleic | 0.5 | 0.2 | * |
| 18:0 Stearic g | 1.7 | 0.9 | * |
| 9c 18:1 Oleic | 8.4 | 4.7 | * |
| 18:2 Linoleic | 5.7 | 4.6 | * |
| 18:3 Gamma Linolenic | 0.0 | 0.0 | * |
| 18:3 Alpha Linolenic | 10.7 | 2.6 | * |
| 20:4 Arachidonic (n6) | 0.2 | 0.2 | * |
| 22:6 Docosahexaenoic | 0.0 | 0.0 | * |
| Total 18:1 trans | 0.0 | 0.0 | * |
| Total 18:1 cis | 0.0 | 0.0 | * |
| Amino Acids | |||
| Aspartic Acid | 1.27 | 1.39 | 0.76 |
| Threonine | 0.58 | 0.48 | 0.22 |
| Serine | 0.83 | 0.66 | 0.25 |
| Glutamic Acid | 2.09 | 2.52 | 0.58 |
| Proline | 0.48 | 0.39 | 0.30 |
| Glycine | 0.45 | 0.36 | 0.24 |
| Alanine | 0.68 | 0.54 | 0.31 |
| Valine | 0.68 | 0.52 | 0.30 |
| Isoleucine | 0.63 | 0.47 | 0.23 |
| Leucine | 0.97 | 0.77 | 0.38 |
| Tyrosine | 0.50 | 0.39 | 0.21 |
| Phenylalanine | 0.55 | 0.47 | 0.23 |
| Lysine | 0.87 | 0.64 | 0.35 |
| Histidine | 0.28 | 0.25 | 0.14 |
| Arginine | 0.75 | 0.79 | 0.23 |
| Cystine | 0.19 | 0.13 | * |
| Methionine | 0.33 | 0.22 | 0.09 |
| Tryptophan | 0.15 | 0.12 | * |
| Cholesterol | 0.58 | 0.39 | * |
| Campesterol | 0.02 | 0.01 | * |
| Stigmasterol | 0.01 | 0.01 | 0.02 |
| Beta Sitosterol | 0.03 | 0.02 | 0.06 |
| Brassicasterol | 0.00 | 0.00 | * |
| Other Sterols/Stanols | 0.03 | 0.03 | * |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Niogret, J.; Savantil, A.B.; Ekayanti, A.; Jaus, M.P.; Wulan, W.; Mitzo, E.; Marelli, J.-P.; Conlong, D. Development of a Diet Production System for Conopomorpha cramerella (Lepidoptera: Gracillariidae), a Major Cocoa Production Pest in Southeast Asia and the Pacific Islands. Insects 2023, 14, 708. https://doi.org/10.3390/insects14080708
Niogret J, Savantil AB, Ekayanti A, Jaus MP, Wulan W, Mitzo E, Marelli J-P, Conlong D. Development of a Diet Production System for Conopomorpha cramerella (Lepidoptera: Gracillariidae), a Major Cocoa Production Pest in Southeast Asia and the Pacific Islands. Insects. 2023; 14(8):708. https://doi.org/10.3390/insects14080708
Chicago/Turabian StyleNiogret, Jerome, Anisah Binti Savantil, Arni Ekayanti, Mavis Peter Jaus, Wulan Wulan, Elviah Mitzo, Jean-Philippe Marelli, and Desmond Conlong. 2023. "Development of a Diet Production System for Conopomorpha cramerella (Lepidoptera: Gracillariidae), a Major Cocoa Production Pest in Southeast Asia and the Pacific Islands" Insects 14, no. 8: 708. https://doi.org/10.3390/insects14080708