
Improved Quality Management of the Indian Meal Moth, Plodia interpunctella (Hübner) (Lepidoptera: Pyralidae) for Enhanced Efficacy of the Sterile Insect Technique


Abstract
:Simple Summary
Abstract
1. Introduction
2. Materials and Methods
2.1. Experimental Procedures
Insect Rearing
2.2. Irradiation Procedure
2.3. Irradiation throughout Ontogenesis
2.3.1. Egg Irradiation
2.3.2. Larval Irradiation
2.3.3. Pupal Irradiation
2.4. Induced Sterility and Mating Competitiveness
2.4.1. Crossing Schedule
2.4.2. Irradiated Eggs
2.4.3. Irradiated Larvae
2.4.4. Irradiated Pupae
2.4.5. Overflooding Ratio
2.5. Low-Temperature Treatment of Male Pupae and Mating Performance
2.6. Flight Cylinder Bioassay
2.7. Assessment of DNA Damage in Cold Preserved Irradiated Pupae
2.8. Statistical Analysis
3. Results
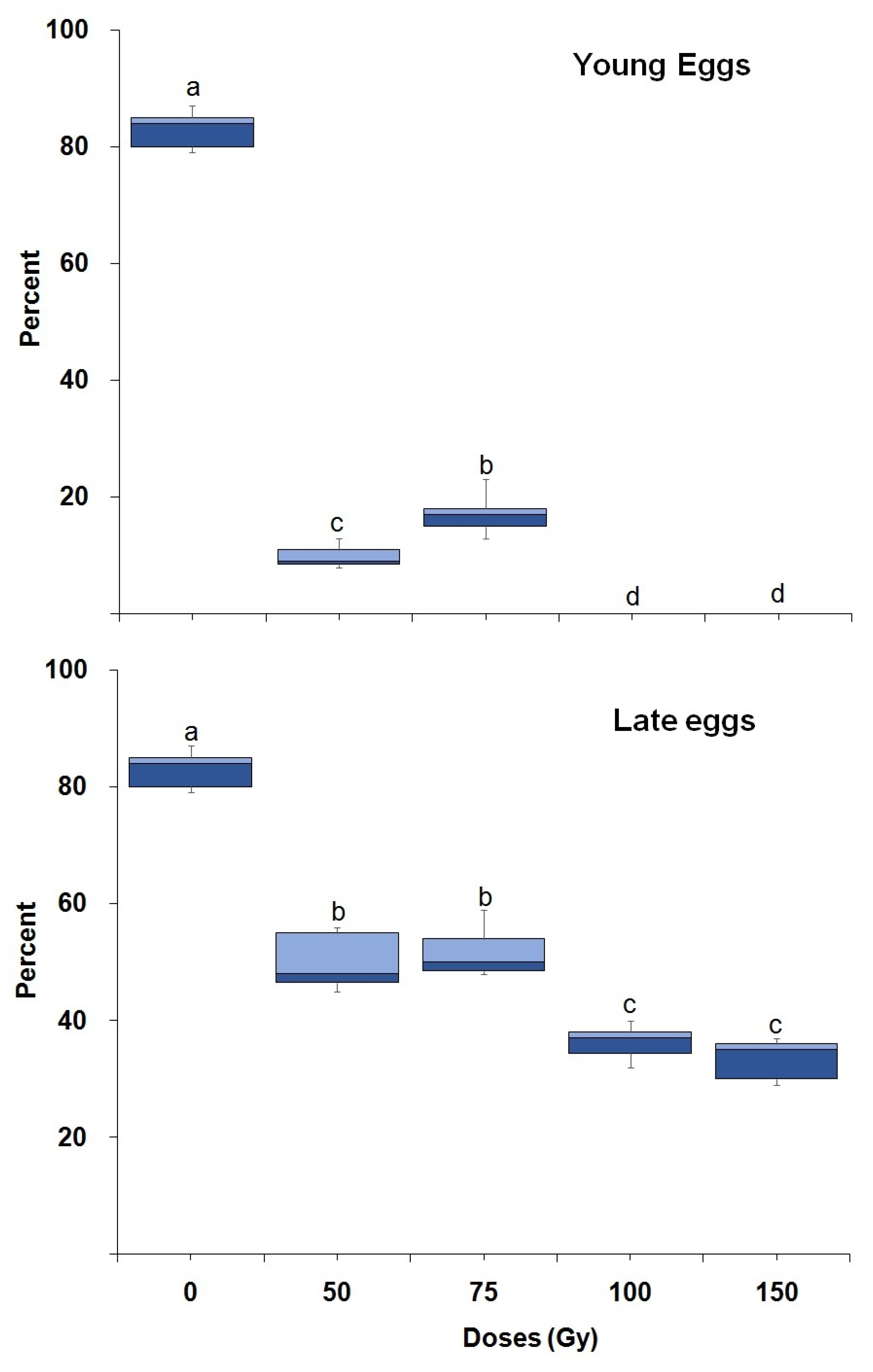
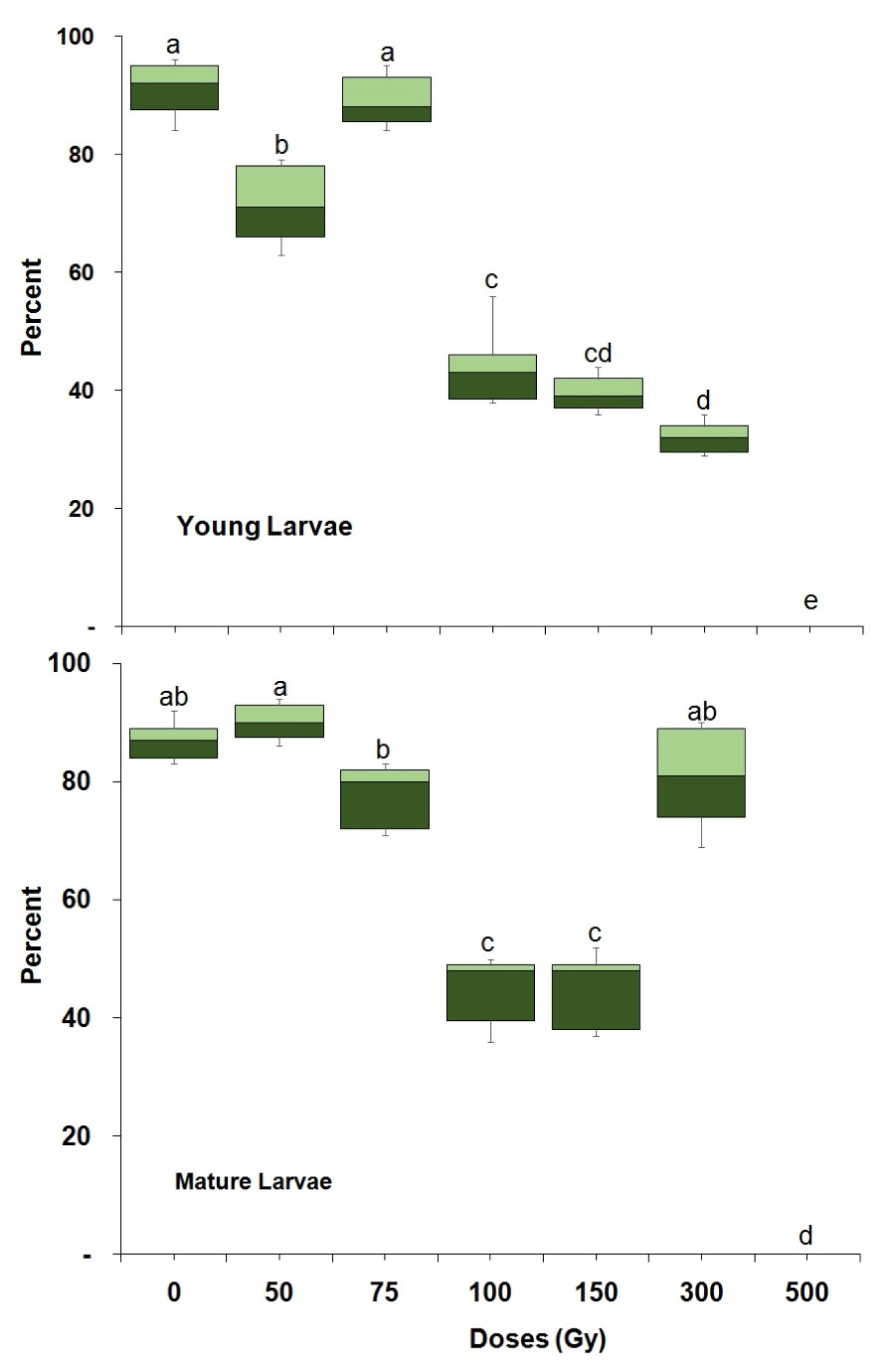
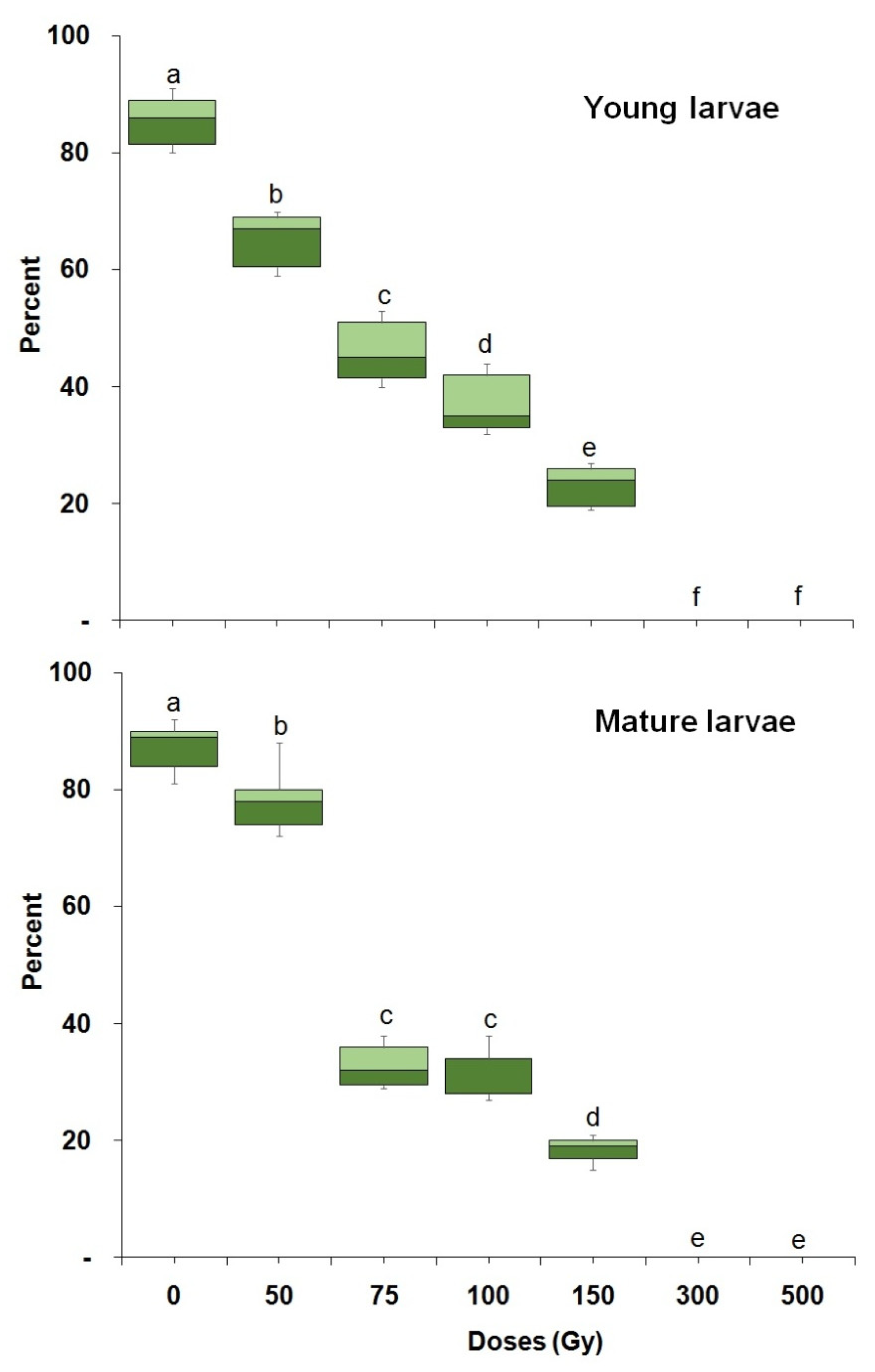
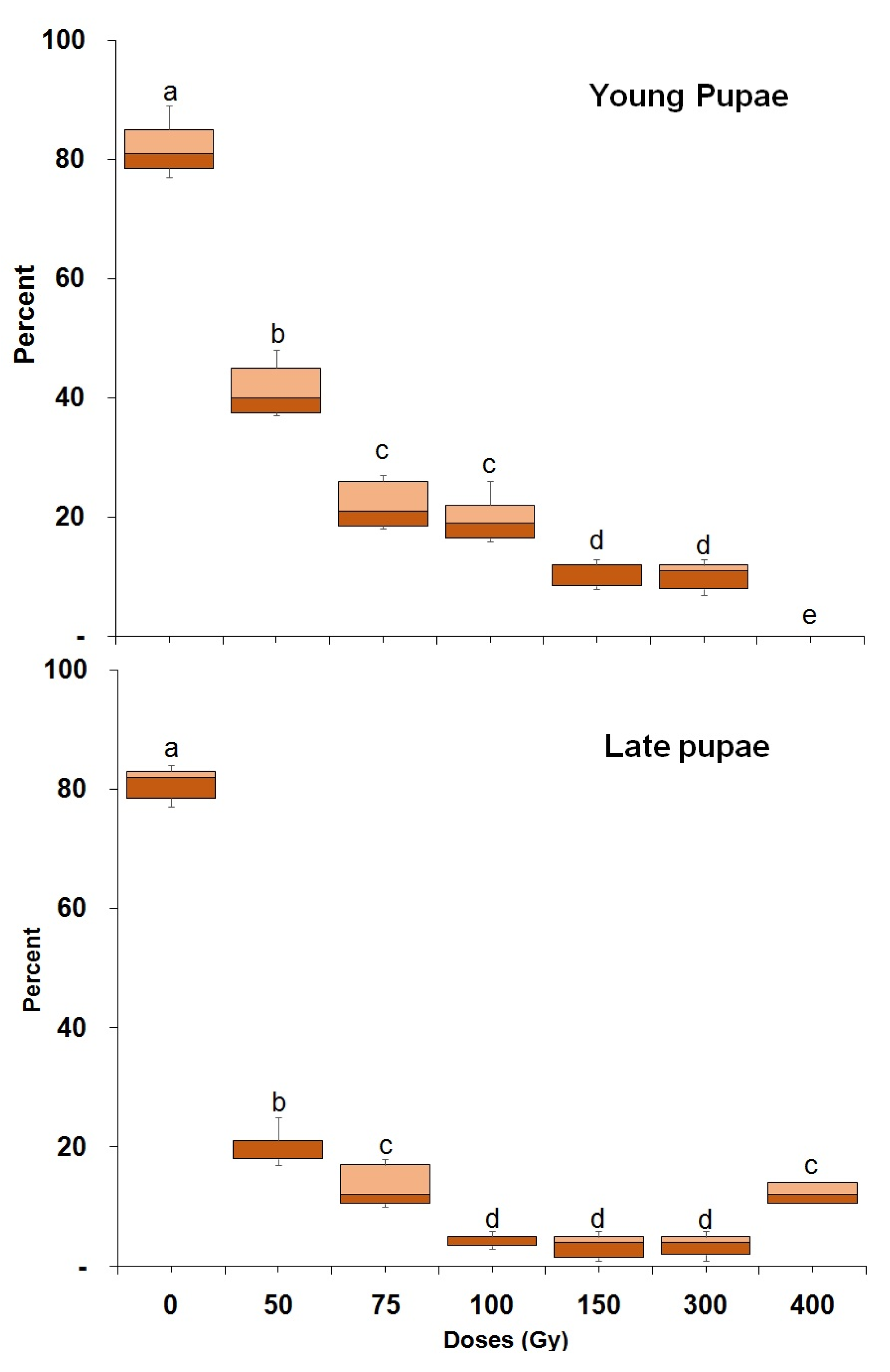
3.1. Radiation Tolerance throughout Ontogeny
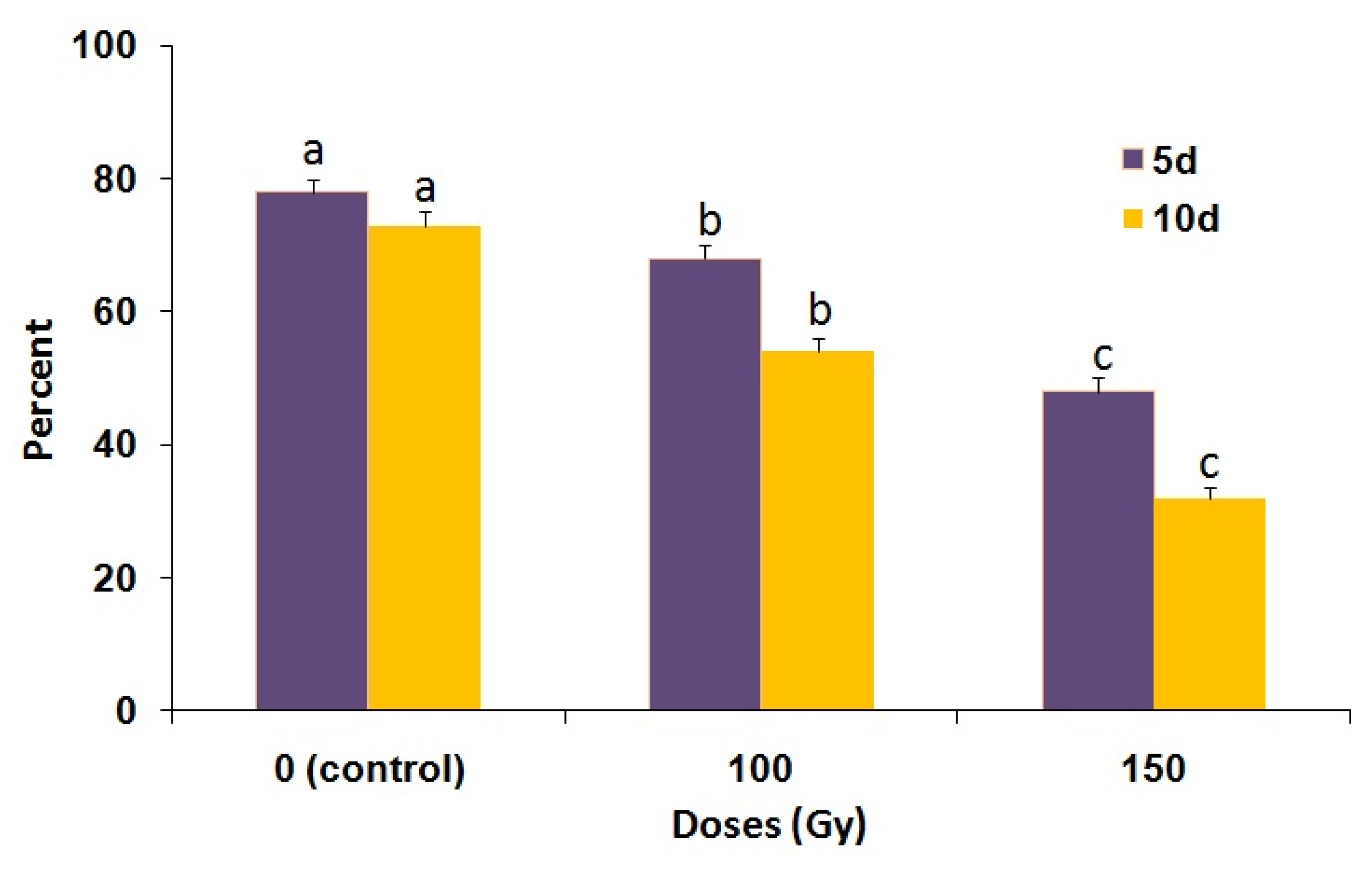
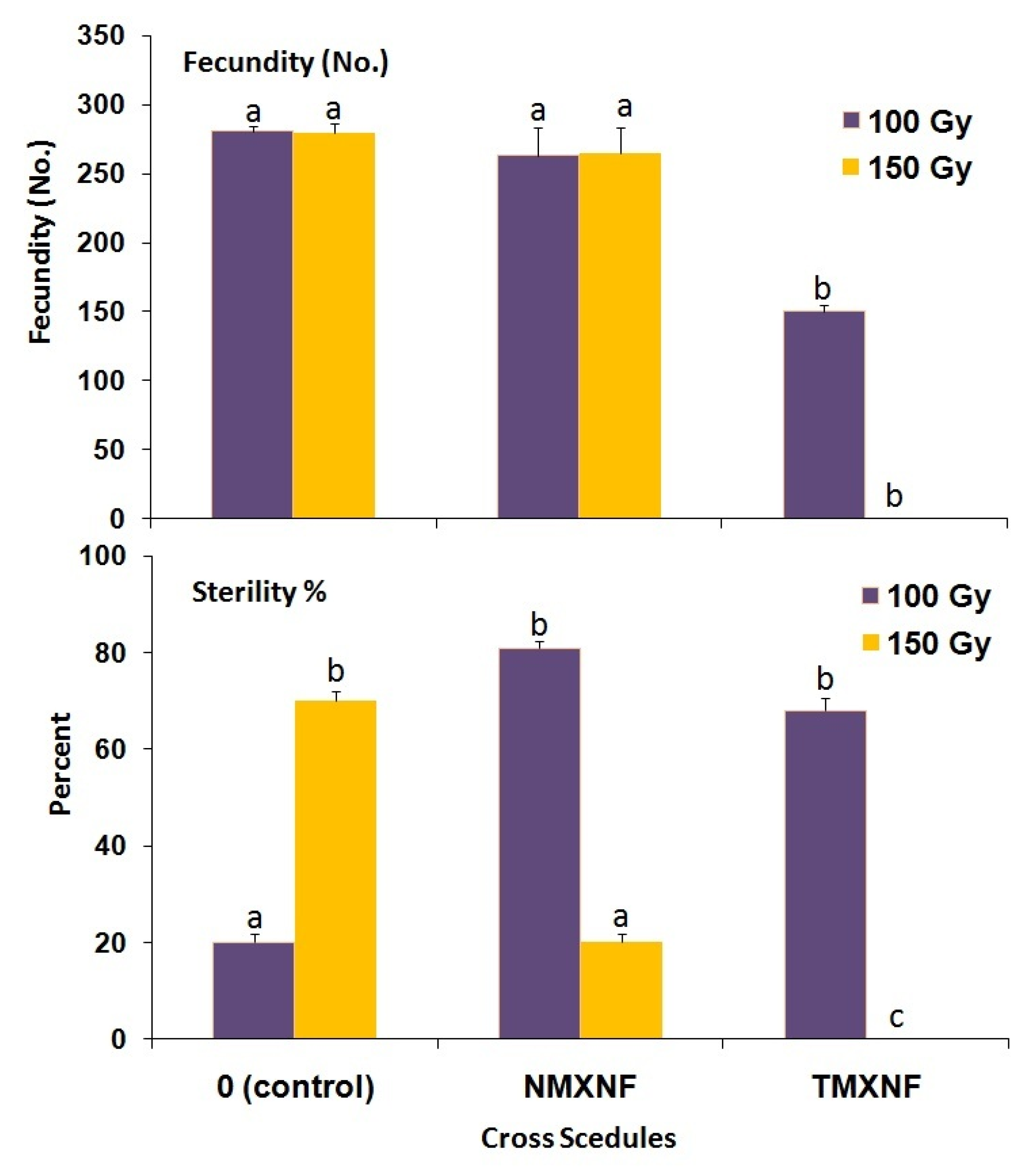
3.2. Induced Sterility and Mating Competitiveness
3.3. Low-Temperature Treatment of Pupae and Male Mating Performance
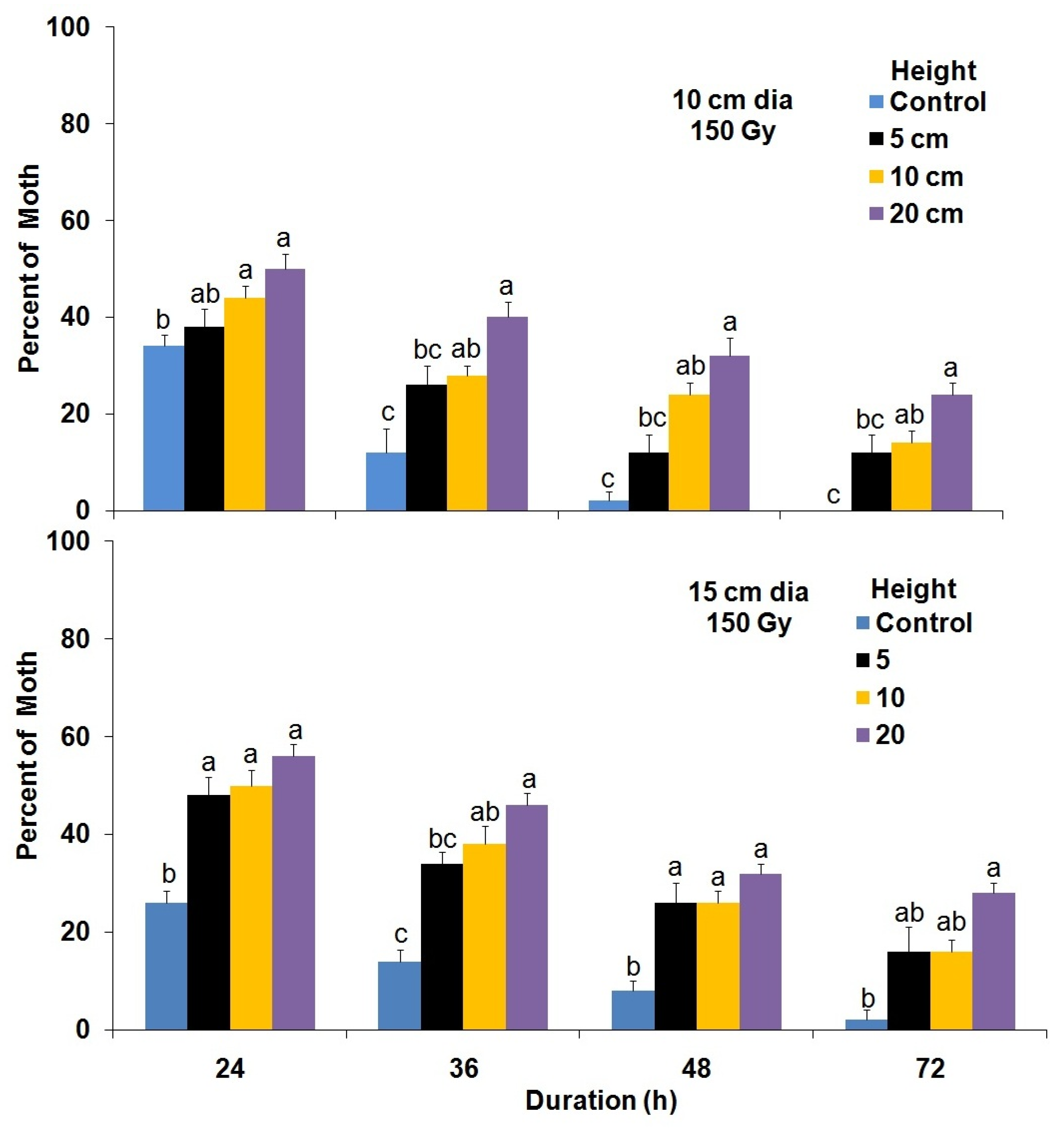
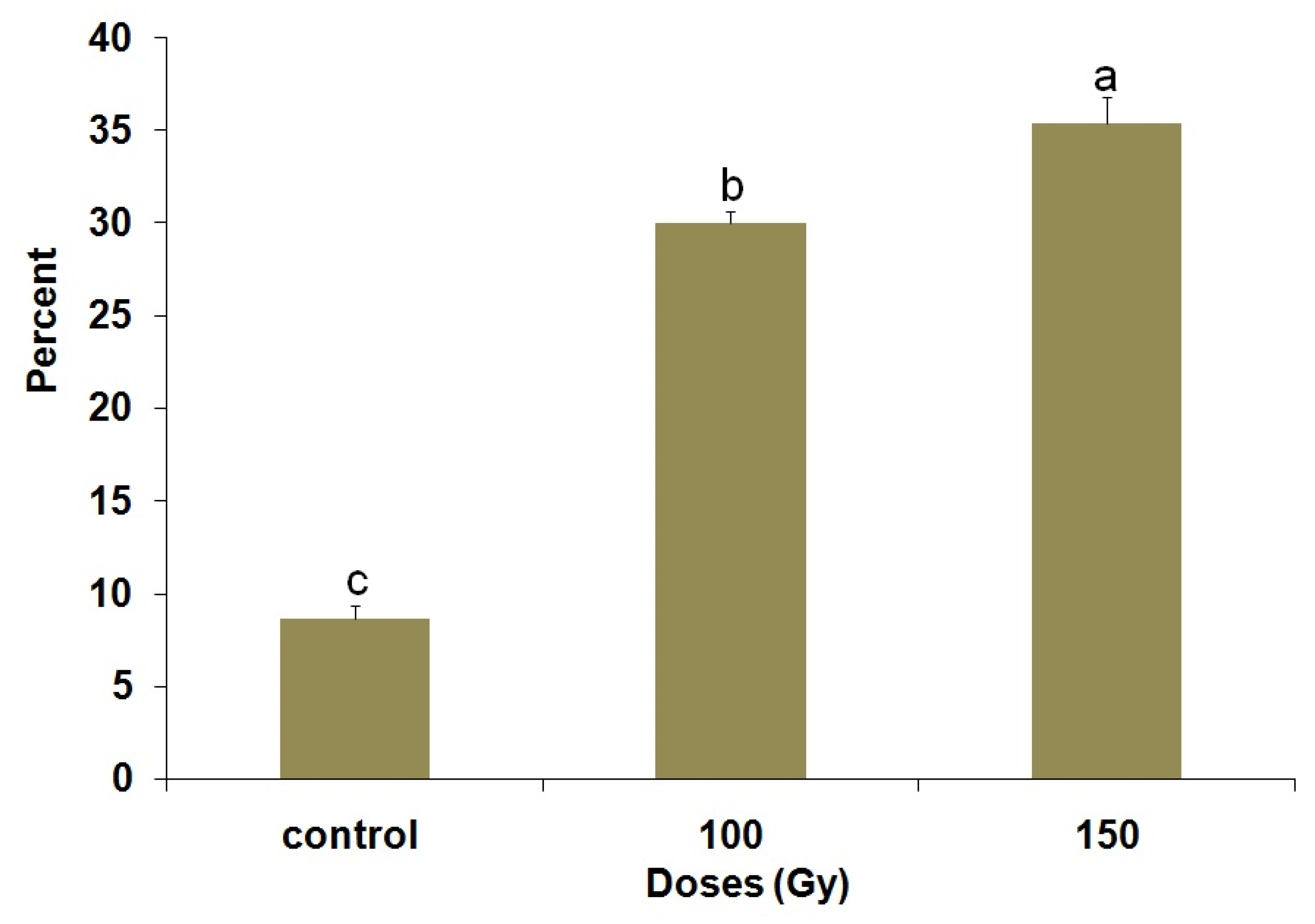
3.4. Flight Cylinder Bioassay
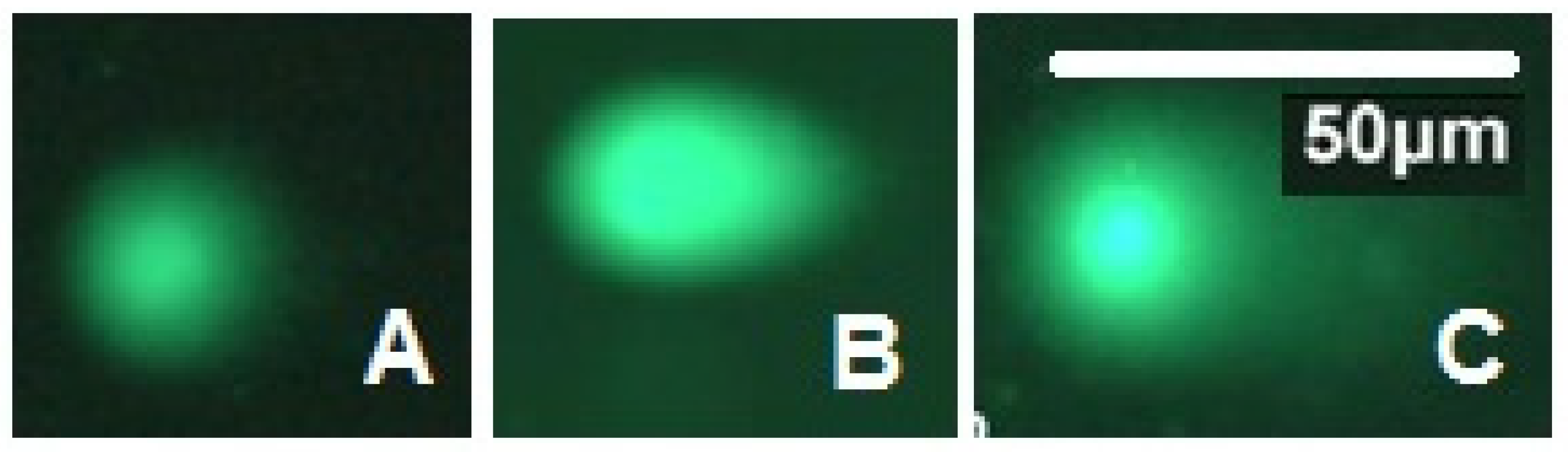
3.5. Assessment of DNA Damage in Cold Treated and Irradiated Pupae
4. Discussion
5. Conclusions
Author Contributions
Funding
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Environmental Protection Agency (EPA). Methyl Bromide; Environmental Protection Agency, United States Department of Agriculture: Washington, DC, USA, 2017. [Google Scholar]
- Phillips, T.W.; Cogan, P.M.; Fadamiro, H.Y. Pheromones. In Alternatives to Pesticides in Stored-Product IPM; Subramanyam, B., Hagstrum, D.W., Eds.; Kluwer Academic Publishers: London, UK, 2000; pp. 273–302. [Google Scholar]
- White, N.D.G.; Leesch, J.H. Chemical control. In Integrated Management of Insects in Stored Products; Subramanyam, B., Hagstrum, D.W., Eds.; Marcel Dekker: New York, UK, USA, 1996; pp. 287–330. [Google Scholar]
- Arthur, F.H.; Phillips, T.W. Stored-product insect pest management and control. In Food Plant Sanitation; Hui, Y.H., Bruinsma, B.L., Gorham, J.R., Nip, W.K., Tong, P.S., Ventresca, P., Eds.; Marcel Dekker: New York, NY, USA, 2003; pp. 341–358. [Google Scholar]
- Attia, F.I. Insecticide resistance in Plodia interpunctella (Hubner) (Lepidoptera: Pyralidae) in New South Wales, Australia. J. Aust. Ent. Soc. 1977, 16, 149–152. [Google Scholar] [CrossRef]
- Attia, F.I. Synergism with organophosphorus-resistance strains of lesser grain brorer, Rhyzopertha dominica (F.). In Proceedings of the 1st Australian Stored Grai Pest Control Conference, Melbourne, Australia, 18–22 May 1981. [Google Scholar]
- Zettler, J.L.; McDonald, L.L.; Redlinger, L.M.; Jones, R.D. Plodia interpunctella and Cadra cautella resistance in strains to malathion and synergized pyrethrins. J. Econ. Entomol. 1973, 66, 1049–1050. [Google Scholar] [CrossRef]
- Zettler, J.L. Insecticide resistance in selected stored-product insects infesting peanuts in the Southeastern United States. J. Econ. Entomol. 1982, 75, 359–362. [Google Scholar] [CrossRef]
- Arthur, F.H.; Zetter, J.L.; Halliday, W.R. Insecticide resistance among populations of almond moth and Indian meal moth (Lepidoptera: Pyralidae) in stored peanuts. J. Econ. Entomol. 1988, 81, 1283–1287. [Google Scholar] [CrossRef]
- Sumner, T.J.; Rochester, G.K.; Hall, G. Silicon drift chambers for X-ray astronomy. Nucl. Instrum. Meth. A 1988, 273, 701–705. [Google Scholar] [CrossRef]
- Schaafsma, A.W. Resistance to malathion in populations of Indian meal moth, Plodia interpunctella (Lepidoptera: Pyralidae). Proc. Entomol. Soc. Ont. 1990, 121, 101–114. [Google Scholar]
- Subramanyam, B.; Hagstrum, D.W. Sampling. In Integrated Management of Insects in Stored Products; Subramanyam, B., Hagstrum, D.W., Eds.; Marcel Dekker, Inc.: New York, NY, USA, 1995; pp. 135–193. [Google Scholar]
- Johnson, J.A.; Wofford, P.L.; Gill, R.F. Development thresholds and degree-day accumulations of Indian meal moth (Lepidoptera: Pyralidae) on dried fruits and nuts. J. Econ. Entomol. 1995, 88, 734–741. [Google Scholar] [CrossRef]
- Van-Rie, J.; McGaughey, W.H.; Johnson, D.E.; Barnett, B.D.; VanMellaer, T.H. Mechanism of insect resistance to the microbial insecticide Bacillus thuringiensis. Science 1990, 247, 72–74. [Google Scholar] [CrossRef]
- McGaughey, W.H.; Johnson, D.E. Indian meal moth (Lepidoptera: Pyralidae) resistance to different strains and mixtures of Bacillus thuringiensis. J. Econ. Entomol. 1992, 85, 1594–1600. [Google Scholar] [CrossRef]
- Herrero, S.; Oppert, B.; Ferre, J. Different mechanisms of resistance to Bacillus thuringiensis toxins in the Indian meal moth. Appl. Environ. Microb. 2001, 67, 1085–1089. [Google Scholar] [CrossRef] [Green Version]
- Lapidot, M.; Saveanu, S.; Padova, R.; Ross, I.; Insect disinfestation by irradiation. Insect Disinfestation of food and Agricultural Products by Irradiation. Proc. IAEA Vienna 1991, 103. Available online: https://citeseerx.ist.psu.edu/document?repid=rep1&type=pdf&doi=91fddb83dda1de8fe3c110b3e0c045614e33ecf8#page=103 (accessed on 1 March 2023).
- Siegwart, M.; Graillot, B.; Lopez, C.B.; Besse, S.; Bardin, M.; Nicot, P.C.; Lopez-Ferber, M. Resistance to bio-insecticides or how to enhance their sustainability: A review. Front. Plant Sci. 2015, 6, 381. [Google Scholar] [CrossRef] [Green Version]
- Marec, F.; Vreysen, M.J.B. Advances and Challenges of Using the Sterile Insect Technique for the Management of Pest Lepidoptera. Insects 2019, 10, 371. [Google Scholar] [CrossRef] [Green Version]
- Dyck, V.A.; Hendrichs, J.; Robinson, A.S. (Eds.) Sterile Insect Technique: Principles and Practice in Area-Wide Integrated Pest Management; CRC Press: Boca Raton, FL, USA, 2021; 1216p. [Google Scholar]
- Carpenter, J.E.; Bloem, S.; Marec, F. Inherited sterility in insects. In Sterile Insect Technique. Principles and Practice in Area-Wide Integrated Pest Management; Dyck, V.A., Hendrichs, J., Robinson, A.S., Eds.; Springer: Dordrecht, The Netherlands, 2005; pp. 115–146. [Google Scholar]
- Ayvaz, A.; Albayrak, A.; Karaborklu, S. Gamma radiation sensitivity of the eggs, larvae and pupae of Indian meal moth Plodia interpunctella (Hübner) (Lepidoptera: Pyralidae). Pest Manag. Sci. 2008, 64, 505. [Google Scholar] [CrossRef]
- Hood-Nowotny, R.; Harari, A.; Seth, R.; Wee, S.-L.; Conlong, D.E.; Suckling, D.M.; Lebdi-Grissa, K.; Simmons, G.; Carpenter, J.E. Stable isotope markers differentiate between mass-reared and wild Lepidoptera in sterile insect technique programs. Fla. Entomol. 2016, 99, 166–176. [Google Scholar] [CrossRef] [Green Version]
- Vreysen, M.J.B.; Robinson, A.S. Ionising radiation and area-wide management of insect pests to promote sustainable agriculture. Agron. Sustain. Dev. 2011, 31, 233–250. [Google Scholar] [CrossRef] [Green Version]
- Vreysen, M.J.B.; Klassen, W.; Carpenter, J.E. Overview of technological advances toward greater efficiency and efficacy in sterile insect-inherited sterility programs against moth pests. Fla. Entomol. 2016, 99, 1–12. [Google Scholar] [CrossRef] [Green Version]
- Knipling, E.F. The Basic Principles of Insect Suppression and Management; Agricultural Handbook 512; United States Department of Agriculture, Agricultural Research Service: Washington, DC, USA, 1979.
- Klassen, W. Area-wide integrated pest management and the sterile insect technique. In Sterile Insect Technique. Principles and Practice in Area-Wide Integrated Pest Management; Dyck, V.A., Hendrichs, J., Robinson, A.S., Eds.; Springer: Dordrecht, The Netherlands, 2005; pp. 39–68. [Google Scholar]
- Zhang, K.; Fu, H.; Zhu, S.; Li, Z.; Weng, Q.F.; Hu, M.Y. Influence of irradiation on flight ability and dispersal of Conopomorpha sinensis (Lepidoptera: Gracillariidae). Fla. Entomol. 2016, 99, 79–86. [Google Scholar] [CrossRef] [Green Version]
- Dyck, V.A.; Hendrichs, J.; Robinson, A.S. (Eds.) Sterile Insect Technique: Principles and Practice in Area-Wide Integrated Pest Management; Springer: Dordrecht, The Netherlands, 2005; 787p. [Google Scholar]
- Zhang, J.H.; Li, N.; Zhao, H.Y.; Wang, Y.Q.; Yang, X.Q.; Wu, K.M. Sterility of Cydia pomonella by X-ray irradiation as an alternative to gamma radiation for the sterile insect technique. Bull. Ent. Res. 2023, 113, 72–78. [Google Scholar] [CrossRef]
- Hallman, G.J. Control of stored product pests by ionizing radiation. J. Stored Prod. Res. 2013, 52, 36–41. [Google Scholar] [CrossRef]
- ISO/ASTM 51261; Standard Guide for Selection and Calibration of Dosimetry Systems for Radiation Processing. American Society for Testing and Materials: West Conshohocken, PA, USA, 2002.
- Hasan, M.; Khan, A.R. Control of stored-product pests by irradiation. Integr. Pest Manage. Rev. 1998, 3, 15–29. [Google Scholar] [CrossRef]
- Fried, M. Determination of Sterile Insect Competitiveness. J. Econ. Entomol. 1971, 64, 869–872. [Google Scholar] [CrossRef]
- Leanage, K.; Wijayaratne, W.; Burk, C.S. Article persistence of mating suppression of the Indian meal moth Plodia interpunctella in the presence and absence of commercial mating disruption dispensers. Insects 2020, 11, 701. [Google Scholar]
- Carpenter, J.E.; Blomefield, T.L.; Vreysen, M.J.B. A flight cylinder bioassay as a simple, effective quality control test for Cydia pomonella. J. Appl. Entomol. 2012, 136, 711–720. [Google Scholar] [CrossRef]
- Hasan, M.; Todoriki, S.; Miyanoshita, A.; Imamura, T. Detection of gamma radiation-induced DNA damage in maize weevil, Sitophilus zeamais Motschulsky (Coleoptera: Curculionidae) assessed using the comet assay. Int. J. Radiat. Biol. 2008, 84, 815–820. [Google Scholar] [CrossRef] [PubMed]
- Sriussadaporn, C.; Yamamoto, K.; Fukushi, K.; Simazaki, D. Comparison of DNA damage detected by plant comet assay in roadside and non-roadside environments. Mutat. Res. 2003, 541, 31–44. [Google Scholar] [CrossRef]
- Levene, H. Contributions to Probability and Statistics. In Essays in Honor of Harold Hetelling; Olkin, I., Gleser, L.J., Perlman, M.D., Press, S.J., Sampson, A.R., Eds.; Stanford University Press: Palo Alto, CA, USA, 1960; pp. 278–292. [Google Scholar]
- SAS Institute. SAS/STAT 9.2 User’s Guide; SAS Institute: Cary, NC, USA, 2008. [Google Scholar]
- LaChance, L.E. Dominant lethal mutations in insects with holokinetic chromosomes. 2. Irradiation of sperm of cabbage looper. Ann. Entomol. Soc. Am. 1974, 67, 35–39. [Google Scholar] [CrossRef]
- Nayak, M.K.; Daglish, G.J.; Phillips, T.W.; Ebert, P.R. Resistance to the Fumigant Phosphine and Its Management in Insect Pests of Stored Products: A Global Perspective. Ann. Rev. Ent. 2020, 65, 333–350. [Google Scholar] [CrossRef] [Green Version]
- Lampiri, E.; Athanassiou, C.G. Insecticidal Effect of phosphine on eggs of the khapra beetle (Coleoptera: Dermestidae). J. Econ. Ent. 2021, 114, 1389–1400. [Google Scholar] [CrossRef]
- Acheson, D.; Steele, J.H. Food Irradiation: A Public Health Challenge for the 21st Century. Clin. Infect. Dis. 2001, 33, 376–377. [Google Scholar]
- Berg, G.J.; LaChance, L.E. Dominant lethal mutations in insects with holokinetic chromosomes: Irradiation of pink bollworm sperm. Ann. Entomol. Soc. Am. 1976, 69, 971–976. [Google Scholar] [CrossRef]
- Yamada, H.; Dias, V.S.; Parker, A.G.; Maiga, H.; Kraupa, C.; Vreysen, M.J.B.; Mamai, W.; Schetelig, M.F.; Bimbilé, N.S.; Somda, N.S.B.; et al. Radiation dose-rate is a neglected critical parameter in dose–response of insects. Sci. Rep. 2022, 12, 6242. [Google Scholar] [CrossRef]
- Fezza, T.J.; Follett, P.A.; Shelly, T.E. The timing of pupal irradiation and the quality and sterility of oriental fruit flies (Diptera: Tephritidae) for use in sterile insect technique. Appl. Entomol. Zool. 2021, 56, 443–450. [Google Scholar] [CrossRef]
- Jiang, S.; He, L.M.; He, W.; Zhao, H.Y.; Yang, X.M.; Yang, X.Q.; Wu, K.M. Effects of X-ray irradiation on the fitness of the established invasive pest fall armyworm Spodoptera frugiperda. Pest Manag. Sci. 2022, 78, 2806–2815. [Google Scholar] [CrossRef]
- LaChance, L.E.; Graham, C.K. Insect radiosensitivity: Dose curves and dose-fractionation studies of dominant lethal mutations in the mature sperm of 4 insect species. Mutat. Res. 1984, 127, 49–59. [Google Scholar] [CrossRef]
- Rendón, P.; McInnis, D.; Lance, D.; Stewart, J. Medfly (Diptera: Tephritidae) genetic sexing: Large-scale field comparison of males-only and bisexual sterile fly releases in Guatemala. J. Econ. Entomol. 2004, 97, 1547–1553. [Google Scholar] [CrossRef]
- Marec, F.; Neven, L.G.; Robinson, A.S.; Vreysen, M.; Goldsmith, M.R.; Nagaraju, J.; Franzn, G. Development of genetic sexing strains in Lepidoptera: From traditional to transgenic approaches. J. Econ. Entomol. 2005, 98, 248–259. [Google Scholar] [CrossRef]
- Goldsmith, M.R.; Shimada, T.; Abe, H. The genetics and genomics of the silkworm, Bombyx mori. Ann. Rev. Entomol. 2005, 50, 71–100. [Google Scholar] [CrossRef]
- Haque, A.; Islam, S.; Bari, A.; Hossain, A.; Athanassiou, C.G.; Hasan, M. Cold storage-mediated rearing of Trichogramma evanescens Westwood on eggs of Plodia interpunctella (Hübner) and Galleria mellonella L. PLoS ONE 2021, 16, e0253287. [Google Scholar] [CrossRef]
- Karimi, N.; Ahmadi, M.; Imani, S.; Marouf, A. Integrated effect of gamma radiation and temperature on Plodia interpunctella (Lepidoptera: Pyralidae). J. Crop Prot. 2022, 11, 201–209. [Google Scholar]
- Athanassiou, C.G.; Arthur, F.H.; Hartzer, K.L. Efficacy of low temperatures for the control of all life stages of Plodia interpunctella and Liposcelis bostrychophila. J. Pest. Sci. 2018, 91, 1363–1369. [Google Scholar] [CrossRef]











{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Stages | N | LC50 (Gy) (95% Fiducial Limits) | LC99 (Gy) (95% Fiducial Limits) | Slope ± SE | Intercept ± SE | χ2 Values |
|---|---|---|---|---|---|---|
| Young eggs | 1280 | 21.37 (13.67–39.22) | 154.84 (98.90–216.34) | 0.66 ± 0.12 | −27.73 ± 20.77 | 151.88 (p < 0.001) (df,10) |
| Late eggs | 1280 | 59.36 (30.42–76.11) | 206.18 (159.27–315.10) | 1.23 ± 0.16 | −44.67 ± 17.58 | 36.26 (p < 0.001) (df,10) |
| Young larvae | 1800 | 51.48 (37.75–61.78) | 299.29 (227.85–486.96) | 0.03 ± 0.56 | 165.33 ± 55.83 | 55.01 (p < 0.001) (df,16) |
| Mature larvae | 1800 | 58.89 (49.59–66.29) | 230.87 (193.38–302.78) | 0.06 ± 0.55 | 163.71 ± 54.89 | 42.62 (p < 0.001) (df,16) |
| Young pupae | 2100 | 26.20 (19.34–41.08) | 360.53 (211.24–437.14) | 0.09 ± 0.37 | 98.14 ± 31.21 | 72.29 (p < 0.001) (df,16) |
| Late pupae | 2100 | 16.19 (11.27–25.31) | 343.92 (296.16–448.02) | 0.06 ± 0.55 | −63.18 ± 26.31 | 39.18 (p < 0.001) (df,16) |
| Cross Schedules | Late Eggs | Mature Larvae | Pupae | ||||||
|---|---|---|---|---|---|---|---|---|---|
| Fecundity (No.) | Sterility% | CI* | Fecundity (No.) | Sterility% | CI | Fecundity (No.) | Sterility% | CI | |
| UF × UM | 186.20 ± 2.62 a | 18.40 ± 1.2 b | 173.73 ± 3.73 a | 18.80 ± 1.5 c | 284.80 ± 5.99 a | 19.40 ± 1.33 b | |||
| TM × UF | 158.00 ± 4.71 c | 92.40 ± 1.4 a | 0.75 | 125.00 ± 3.02 b | 95.40 ± 1.0 b | 1.00 | 166.20 ± 7.44 c | 100 ± 00 a | 1.01 |
| TM × TF | 137.80 ± 2.36 b | 87.60 ± 1.3 a | 99.20 ± 3.82 c | 100 ± 00 a | 146.20 ± 8.17 b | 100 ± 00 a | |||
| Doses (Gy) | Flooding Ratio Im:Um:Uf | Late Eggs | Mature Larvae | Pupae | ||||||
|---|---|---|---|---|---|---|---|---|---|---|
| Fecundity (No.) | Sterility% | CI* | Fecundity (No.) | Sterility% | CI | Fecundity (No.) | Sterility% | CI | ||
| 0:1:1 | 256.80 ± 4.08 a | 20.00 ± 0.84 c | - | 240.00 ± 4.64 a | 16.80 ± 1.89 c | - | 272.60 ± 5.62 a | 20.20 ± 0.86 d | - | |
| 1:0:1 | 121.30 ± 2.39 b | 51.60 ± 1.97 b | - | 139.20 ± 5.66 b | 47.40 ± 2.39 b | - | 193.80 ± 4.04 b | 48.40 ± 3.57 c | - | |
| 100 | 1:1:1 | 114.60 ± 3.24 c | 88.40 ± 2.61 a | 0.37 | 121.80 ± 3.94 c | 92.00 ± 2.94 a | 0.28 | 55.00 ± 4.86 b | 83.60 ± 2.93 a | 0.42 |
| 5:1:1 | 107.90 ± 4.24 d | 93.40 ± 1.97 a | 0.93 | 102.40 ± 4.54 c | 96.20 ± 1.07 a | 0.70 | 50.20 ± 2.23 c | 98.20 ± 0.80 a | 1.00 | |
| 0:1:1 | 256.40 ± 1.73 a | 20.60 ± 0.68 d | - | 268.40 ± 4.88 a | 18.20 ± 0.86 c | - | 280.40 ± 6.21 a | 17.00 ± 0.71 c | - | |
| 1:0:1 | 109.60 ± 3.55 b | 63.80 ± 1.28 c | - | 127.80 ± 3.39 b | 43.20 ± 3.32 b | - | 179.60 ± 3.85 b | 58.20 ± 4.74 b | - | |
| 150 | 1:1:1 | 98.20 ± 4.96 c | 87.60 ± 2.95 b | 0.29 | 115.60 ± 2.98 c | 94.20 ± 1.69 a | 0.43 | 149.00 ± 3.78 c | 99.20 ± 0.38 a | 0.21 |
| 5:1:1 | 87.80 ± 3.60 c | 100 ± 00 d | 0.72 | 113.80 ± 3.64 d | 100 ± 00 a | 1.0 | 127.80 ± 5.43 d | 100 ± 00 a | 0.53 | |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Hasan, M.M.; Hossain, M.A.; Athanassiou, C.G. Improved Quality Management of the Indian Meal Moth, Plodia interpunctella (Hübner) (Lepidoptera: Pyralidae) for Enhanced Efficacy of the Sterile Insect Technique. Insects 2023, 14, 344. https://doi.org/10.3390/insects14040344
Hasan MM, Hossain MA, Athanassiou CG. Improved Quality Management of the Indian Meal Moth, Plodia interpunctella (Hübner) (Lepidoptera: Pyralidae) for Enhanced Efficacy of the Sterile Insect Technique. Insects. 2023; 14(4):344. https://doi.org/10.3390/insects14040344
Chicago/Turabian StyleHasan, Md. Mahbub, Md. Akhtar Hossain, and Christos G. Athanassiou. 2023. "Improved Quality Management of the Indian Meal Moth, Plodia interpunctella (Hübner) (Lepidoptera: Pyralidae) for Enhanced Efficacy of the Sterile Insect Technique" Insects 14, no. 4: 344. https://doi.org/10.3390/insects14040344