
Integrative Taxonomy of the Gall Mite Nothopoda todeica n. sp. (Eriophyidae) from the Disjunct Afro-Australasian Fern Todea barbara: Morphology, Phylogeny, and Mitogenomics †

Abstract
:Simple Summary
Abstract
1. Introduction
2. Materials and Methods
3. Results
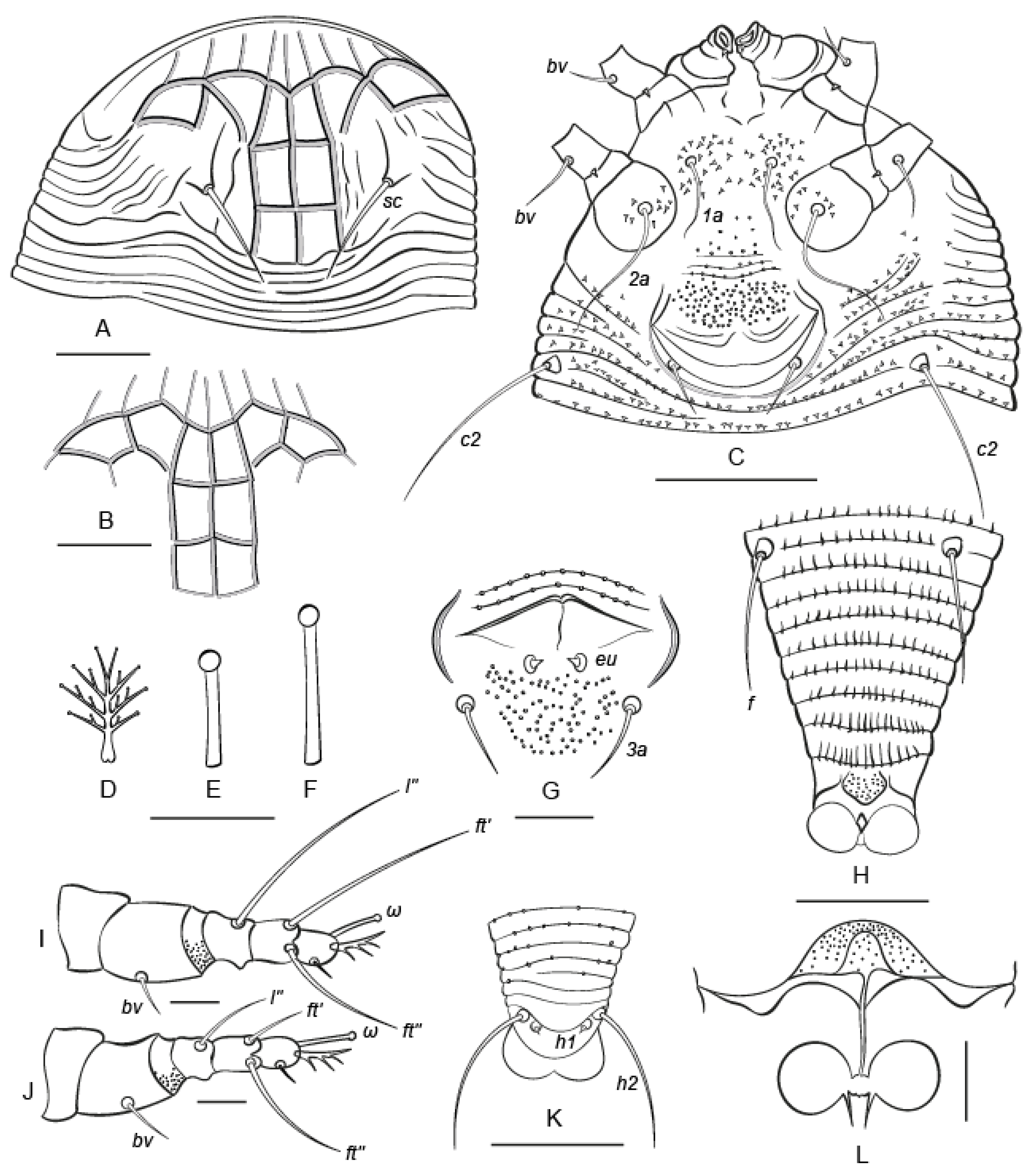
3.1. Taxonomy
Nothopoda todeica n. sp. (Figure 3 and Figure 4)

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Character | N. todeica n. sp. | N. footei | N. natalensis |
|---|---|---|---|
| Median line of prodorsal shield | Entire, simple | Forked anteriorly | Entire, simple |
| Microtubercles on lateral areas of prodorsal shield | Absent | Present | Present |
| Cells along anterior margin of prodorsal shield | 9–10, along whole anterior anterolateral margin | 2–4, only in antero-medial region | 2–4, only in antero-medial region |
| Ornamentation of genital coverflap | Two transverse rows of striae | Two longitudinal striae | 14 transverse striae |
| Host plant | Todea barbara (L.) T. Moore (Osmundaceae) | Nephrolepis sp. (Nephrolepidaceae) | Aspilia natalensis (Sond.) Wild (Asteraceae) |
| Relation to host | Vagrant on lower surface of fronds | in terminal galls on the frond branches | in erineum within dome-like gall |
| Reference | This paper | [54] | [55] |



3.2. Molecular Phylogenetics
3.2.1. Blast Search Results for Cox1 and rDNA Sequence of Nothopoda todeica n. sp.
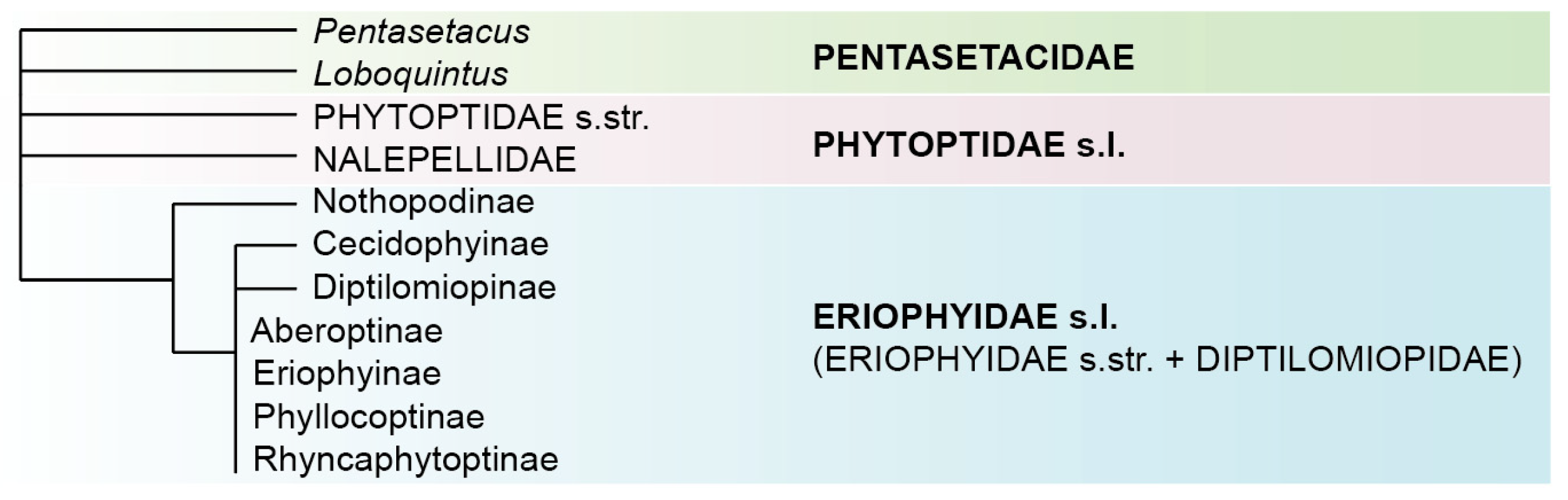
3.2.2. Molecular Phylogenetic Analyses (Figure 6)

3.2.3. Comparative Mitogenomics
4. Discussion
Author Contributions
Funding
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Nuzzaci, G.; Alberti, G. Internal anatomy and physiology. In Eriophyoid Mites: Their Biology, Natural Enemies and Control; World Crop Pests; Lindquist, E.E., Sabelis, M.W., Bruin, J., Eds.; Elsevier Science Publishing: Amsterdam, The Netherlands, 1996; Volume 6, pp. 101–150. [Google Scholar] [CrossRef]
- Sidorchuk, E.A.; Schmidt, A.R.; Ragazzi, E.; Roghi, G.; Lindquist, E.E. Plant-feeding mite diversity in Triassic amber (Acari: Tetrapodili). J. Syst. Palaeontol. 2015, 13, 129–151. [Google Scholar] [CrossRef]
- Propistsova, E.A.; Makarova, A.A.; Chetverikov, P.E.; Polilov, A.A. Anatomy of the miniature four-legged mite Achaetocoptes quercifolii (Arachnida: Acariformes: Eriophyoidea). Arthropod Struct. Dev. 2023, 72, 101235. [Google Scholar] [CrossRef] [PubMed]
- Lindquist, E.E. External anatomy and notation of structures. In Eriophyoid Mites: Their Biology, Natural Enemies and Control; World Crop Pests; Lindquist, E.E., Sabelis, M.W., Bruin, J., Eds.; Elsevier Science Publishing: Amsterdam, The Netherlands, 1996; Volume 6, pp. 3–31. [Google Scholar] [CrossRef]
- Bolton, S.J.; Chetverikov, P.E.; Klompen, H. Morphological support for a clade comprising two vermiform mite lineages: Eriophyoidea (Acariformes) and Nematalycidae (Acariformes). Syst. Appl. Acarol. 2017, 22, 1096–1131. [Google Scholar] [CrossRef]
- Bolton, S.J.; Bauchan, G.R.; Chetverikov, P.E.; Ochoa, R.; Klompen, H. A rudimentary sheath for the smallest of “biting” chelicerae: The mouthparts of Cunliffea (Nematalycidae) and a new hypothesis on the origin of the stylet sheath of Eriophyoidea (Acariformes). Int. J. Acarol. 2018, 44, 374–381. [Google Scholar] [CrossRef]
- Klimov, P.B.; OConnor, B.M.; Chetverikov, P.E.; Bolton, S.J.; Pepato, A.R.; Mortazavi, A.L.; Tolstikov, A.V.; Bauchan, G.R.; Ochoa, R. Comprehensive phylogeny of acariform mites (Acariformes) provides insights on the origin of the four-legged mites (Eriophyoidea), a long branch. Mol. Phylogenet. Evol. 2018, 119, 105–117. [Google Scholar] [CrossRef]
- Klimov, P.B.; Chetverikov, P.E.; Dodueva, I.E.; Vishnyakov, A.E.; Bolton, S.J.; Paponova, S.S.; Lutova, L.A.; Tolstikov, A.V. Symbiotic bacteria of the gall-inducing mite Fragariocoptes setiger (Eriophyoidea) and phylogenomic resolution of the eriophyoid position among Acari. Sci. Rep. 2022, 12, 3811. [Google Scholar] [CrossRef]
- Pepato, A.R.; Costa, S.G.D.S.; Harvey, M.S.; Klimov, P.B. One-way ticket to the blue: A large-scale, dated phylogeny revealed asymmetric land-to-water transitions in acariform mites (Acari: Acariformes). Mol. Phylogenet. Evol. 2022, 177, 107626. [Google Scholar] [CrossRef]
- Li, H.S.; Xue, X.F.; Hong, X.Y. Homoplastic evolution and host association of Eriophyoidea (Acari, Prostigmata) conflict with the morphological-based taxonomic system. Mol. Phylogenet. Evol. 2014, 78, 185–198. [Google Scholar] [CrossRef]
- Chetverikov, P.E.; Cvrković, T.; Makunin, A.; Sukhareva, S.; Vidović, B.; Petanović, R. Basal divergence of Eriophyoidea (Acariformes, Eupodina) inferred from combined partial COI and 28S gene sequences and CLSM genital anatomy. Exp. Appl. Acarol. 2015, 67, 219–245. [Google Scholar] [CrossRef]
- Xue, X.F.; Dong, Y.; Deng, W.; Hong, X.Y.; Shao, R. The phylogenetic position of eriophyoid mites (superfamily Eriophyoidea) in Acariformes inferred from the sequences of mitochondrial genomes and nuclear small subunit (18S) rRNA gene. Mol. Phylogenet. Evol. 2017, 109, 271–282. [Google Scholar] [CrossRef]
- Szudarek-Trepto, N.; Kaźmierski, A.; Skoracka, A.; Lewandowski, M.; Dabert, J. Molecular Phylogeny Supports the Monophyly of the Mite Supercohort Eupodides (Acariformes: Trombidiformes) and Greatly Coincides with Traditional Morphological Definition of the Taxon. Ann. Zool. 2022, 72, 757–786. [Google Scholar] [CrossRef]
- Amrine, J.W., Jr.; Stasny, T.A.H.; Flechtmann, C.H.W. Revised Keys to the World Genera of the Eriophyoidea (Acari: Prostigmata); Indira Publishing House: West Bloomfield, MI, USA, 2003; pp. 1–244. [Google Scholar]
- Chetverikov, P.E.; Bertone, M. First rhyncaphytoptine mite (Eriophyoidea, Diptilomiopidae) parasitizing american hazelnut (Corylus americana): Molecular identification, confocal microscopy, and phylogenetic position. Exp. Appl. Acarol. 2022, 88, 75–95. [Google Scholar] [CrossRef] [PubMed]
- Bolton, S.J.; Chetverikov, P.E.; Ochoa, R.; Klimov, P.B. Where Eriophyoidea (Acariformes) belong in the tree of life. Insects, 2023; in press. [Google Scholar] [CrossRef]
- Shevchenko, V.G.; Bagnyuk, I.G.; Sukhareva, S.I. A new family of Pentasetacidae (Acariformes, Tetrapodili) and its role in treatment of the origin and evolution of the group. Zool. Ž. 1991, 70, 47–53. [Google Scholar]
- Chetverikov, P.E.; Petanović, R.U. Description of a new early-derivative mite, Pentasetacus plicatus n. sp. (Acariformes, Eriophyoidea), and remarks on the systematic position of pentasetacines. Zootaxa 2016, 4144, 211–226. [Google Scholar] [CrossRef]
- Chetverikov, P.E.; Rector, B.G.; Tonkel, K.; Dimitri, L.; Cheglakov, D.S.; Romanovich, A.E.; Amrine, J. Phylogenetic position of a new Trisetacus mite species (Nalepellidae) destroying seeds of North American junipers and new hypotheses on basal divergence of Eriophyoidea. Insects 2022, 13, 201. [Google Scholar] [CrossRef]
- Xue, X.F.; Guo, J.F.; Dong, Y.; Hong, X.Y.; Shao, R. Mitochondrial genome evolution and tRNA truncation in Acariformes mites: New evidence from eriophyoid mites. Sci. Rep. 2016, 6, 18920. [Google Scholar] [CrossRef]
- Greenhalgh, R.; Dermauw, W.; Glas, J.J.; Rombauts, S.; Wybouw, N.; Thomas, J.; Alba, J.M.; Pritham, E.J.; Legarrea, S.; Feyereisen, R.; et al. Genome streamlining in a minute herbivore that manipulates its host plant. eLife 2020, 9, e56689. [Google Scholar] [CrossRef]
- Yin, Y.; Yao, L.F.; Zhang, Q.; Hebert, P.D.; Xue, X.F. Using multiple lines of evidence to delimit protogynes and deutogynes of four-legged mites: A case study on Epitrimerus sabinae s.l. (Acari: Eriophyidae). Invertebr. Syst. 2020, 34, 757–768. [Google Scholar] [CrossRef]
- Chase, M.W.; Christenhusz, M.J.M.; Fay, M.F.; Byng, J.W.; Judd, W.S.; Soltis, D.E.; Mabberley, D.J.; Sennikov, A.N.; Soltis, P.S.; Stevens, P.F. An update of the Angiosperm Phylogeny Group classification for the orders and families of flowering plants: APG IV. Bot. J. Linn. Soc. 2016, 181, 1–20. [Google Scholar] [CrossRef]
- Skoracka, A.; Smith, L.; Oldfield, G.; Cristofaro, M.; Amrine, J.W. Host-plant specificity and specialization in eriophyoid mites and their importance for the use of eriophyoid mites as biocontrol agents of weeds. Exp. Appl. Acarol. 2010, 51, 93–113. [Google Scholar] [CrossRef] [PubMed]
- Ferreira, B.G.; Álvarez, R.; Bragança, G.P.; Alvarenga, D.R.; Pérez-Hidalgo, N.; Isaias, R.M. Feeding and other gall facets: Patterns and determinants in gall structure. Bot. Rev. 2019, 85, 78–106. [Google Scholar] [CrossRef]
- Westphal, E. Cecidogenesis and resistance phenomena inmite-induced galls. In Biology of Insect-Induced Galls; Shorthouse, J., Rohfritsch, O., Eds.; Oxford University Press: New York, NY, USA, 1992; pp. 141–156. [Google Scholar]
- Parris, B.S. Circum-Antarctic continental distribution patterns in pteridophyte species. Brittonia 2001, 53, 270–283. [Google Scholar] [CrossRef]
- Bomfleur, B.; Escapa, I. A silicified Todea trunk (Osmundaceae) from the Eocene of Patagonia. PalZ 2019, 93, 543–548. [Google Scholar] [CrossRef]
- Carvalho, M.R.; Wilf, P.; Hermsen, E.J.; Gandolfo, M.A.; Cúneo, N.R.; Johnson, K.R. First record of Todea (Osmundaceae) in South America, from the early Eocene paleorainforests of Laguna del Hunco (Patagonia, Argentina). Am. J. Bot. 2013, 100, 1831–1848. [Google Scholar] [CrossRef]
- Bomfleur, B.; Grimm, G.W.; McLoughlin, S. Osmunda pulchella sp. nov. from the Jurassic of Sweden—Reconciling molecular and fossil evidence in the phylogeny of modern royal ferns (Osmundaceae). BMC Evol. Biol. 2015, 15, 126. [Google Scholar] [CrossRef]
- Amrine, J.W., Jr.; Manson, D.C.M. Preparation, mounting and descriptive study of eriophyoid mites. In Eriophyoid Mites: Their Biology, Natural Enemies and Control; World Crop Pests; Lindquist, E.E., Sabelis, M.W., Bruin, J., Eds.; Elsevier Science Publishing: Amsterdam, The Netherlands, 1996; Volume 6, pp. 383–396. [Google Scholar] [CrossRef]
- Chetverikov, P.E. Video projector: A digital replacement for camera lucida for drawing mites and other microscopic objects. Syst. Appl. Acarol. 2016, 21, 1278–1280. [Google Scholar] [CrossRef]
- Katoh, K.; Misawa, K.; Kuma, K.; Miyata, T. MAFFT: A novel method for rapid multiple sequence alignment based on fast Fourier transformation. Nucleic Acids Res. 2002, 30, 3059–3066. [Google Scholar] [CrossRef]
- Katoh, K.; Rozewicki, J.; Yamada, K.D. MAFFT online service: Multiple sequence alignment, interactive sequence choice and visualization. Brief. Bioinform. 2017, 20, 1160–1166. [Google Scholar] [CrossRef]
- Pepato, A.R.; da Rocha, C.E.; Dunlop, J.A. Phylogenetic position of the acariform mites: Sensitivity to homology assessment under total evidence. BMC Evol. Biol. 2010, 10, 235. [Google Scholar] [CrossRef]
- Castresana, J. Selection of conserved blocks from multiple alignments for their use in phylogenetic analysis. Mol. Biol. Evol. 2000, 17, 540–552. [Google Scholar] [CrossRef]
- Talavera, G.; Castresana, J. Improvement of phylogenies after removing divergent and ambiguously aligned blocks from protein sequence alignments. Syst. Biol. 2007, 56, 564–577. [Google Scholar] [CrossRef] [PubMed]
- Dereeper, A.; Guignon, V.; Blanc, G.; Audic, S.; Buffet, S.; Chevenet, F.; Dufayard, J.F.; Guindon, S.; Lefort, V.; Lescot, M.; et al. Phylogeny.fr: Robust phylogenetic analysis for the non-specialist. Nucleic Acids Res. 2008, 36, W465–W469. [Google Scholar] [CrossRef] [PubMed]
- Minh, B.Q.; Schmidt, H.A.; Chernomor, O.; Schrempf, D.; Woodhams, M.D.; von Haeseler, A.; Lanfear, R. IQ-TREE 2: New Models and Efficient Methods for Phylogenetic Inference in the Genomic Era. Mol. Biol. Evol. 2020, 37, 1530–1534. [Google Scholar] [CrossRef] [PubMed]
- Kalyaanamoorthy, S.; Minh, B.Q.; Wong, T.K.F.; von Haeseler, A.; Jermiin, L.S. ModelFinder: Fast model selection for accurate phylogenetic estimates. Nat. Methods 2017, 14, 587–589. [Google Scholar] [CrossRef]
- FastQC: A Quality Control Tool for High throughput Sequence Data. Available online: https://www.bioinformatics.babraham.ac.uk/projects/fastqc/ (accessed on 23 March 2023).
- Chen, S.; Huang, T.; Zhou, Y.; Han, Y.; Xu, M.; Gu, J. AfterQC: Automatic filtering, trimming, error removing and quality control for fastq data. BMC Bioinform. 2017, 18, 80. [Google Scholar] [CrossRef]
- Nurk, S.; Meleshko, D.; Korobeynikov, A.; Pevzner, P.A. metaSPAdes: A new versatile metagenomic assembler. Genome Res. 2017, 27, 824–834. [Google Scholar] [CrossRef]
- Jin, J.-J.; Yu, W.-B.; Yang, J.-B.; Song, Y.; dePamphilis, C.W.; Yi, T.-S.; Li, D.-Z. GetOrganelle: A fast and versatile toolkit for accurate de novo assembly of organelle genomes. Genome Biol. 2020, 21, 241. [Google Scholar] [CrossRef]
- Meng, G.; Li, Y.; Yang, C.; Liu, S. MitoZ: A toolkit for animal mitochondrial genome assembly, annotation and visualization. Nucleic Acids Res. 2019, 47, e63. [Google Scholar] [CrossRef]
- Okonechnikov, K.; Golosova, O.; Fursov, M. Unipro UGENE: A unified bioinformatics toolkit. Bioinformatics 2012, 28, 1166–1167. [Google Scholar] [CrossRef]
- Seemann, T. Barrnap 0.7: Rapid Ribosomal RNA Prediction. 2013. Available online: https://github.com/tseemann/barrnap (accessed on 23 March 2022).
- Quinlan, A.R.; Hall, I.M. BEDTools: A flexible suite of utilities for comparing genomic features. Bioinformatics 2010, 26, 841–842. [Google Scholar] [CrossRef] [PubMed]
- Altschul, S.F.; Madden, T.L.; Schäffer, A.A.; Zhang, J.; Zhang, Z.; Miller, W.; Lipman, D.J. Gapped BLAST and PSI-BLAST: A new generation of protein database search programs. Nucleic Acids Res. 1997, 25, 3389–3402. [Google Scholar] [CrossRef] [PubMed]
- Bernt, M.; Donath, A.; Jühling, F.; Externbrink, F.; Florentz, C.; Fritzsch, G.; Pütz, J.; Middendorf, M.; Stadler, P.F. MITOS: Improved de novo metazoan mitochondrial genome annotation. Mol. Phylogenet. Evol. 2013, 69, 313–319. [Google Scholar] [CrossRef] [PubMed]
- Laslett, D.; Canbäck, B. ARWEN, a program to detect tRNA genes in metazoan mitochondrial nucleotide sequences. Bioinformatics 2008, 24, 172–175. [Google Scholar] [CrossRef] [PubMed]
- Chetverikov, P.E.; Craemer, C.; Gankevich, V.D.; Vishnyakov, A.E.; Zhuk, A.S. A New Webbing Aberoptus Species from South Africa Provides Insight in Silk Production in Gall Mites (Eriophyoidea). Diversity 2023, 15, 151. [Google Scholar] [CrossRef]
- Lv, A.; Deng, X.; Zhao, Y.; Du, S.; Tan, M.; Wang, G. Three new species Nothopoda Keifer (Acari, Nothopodinae) from Guangxi, China. Int. J. Acarol. 2022, 48, 619–627. [Google Scholar] [CrossRef]
- Keifer, H.H. Eriophyoid Studies C-1; Agricultural Research Service, USDA: Washington, DC, USA, 1969; pp. 1–20.
- Meyer, M.S.; Ueckermann, E.A. Afrotropical Eriophyoidea: On some species of the subfamily Nothopodinae (Acari: Eriophyidae). Acarologia 1997, 38, 57–68. [Google Scholar]
- GBIF, Global Biodiversity Information Facility. Available online: https://www.gbif.org (accessed on 23 March 2023).



| Character | N. todeica n. sp. | N. camelliae |
|---|---|---|
| Shape of microtubercles on coxal plates | Spine-shaped | Subspherical |
| Microtubercles on dorsal opisthosomal annuli | Absent | First five dorsal annuli with microtubercles, all other annuli with microtubercles present only midway |
| Microtubercles on ventral opisthosomal annuli | All ventral annuli completely microtuberculated | Microtubercules form three longitudinal bands separated by smooth areas |
| Microgranulations on femora I and II | Present only distally on ventral femora | Present on entire femora surface |
| Short thin lines within cells between ridges of prodorsal shield | Absent | Distinct, numerous |
| Host plant | Todea barbara (L.) T. Moore (Osmundaceae) | Camellia oleifera Abel (Theaceae) |
| Relation to host | Vagrant on lower surface of fronds | Vagrant on lower leaf surface |
| Reference | This paper | [53] |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Chetverikov, P.E.; Craemer, C.; Gankevich, V.D.; Zhuk, A.S. Integrative Taxonomy of the Gall Mite Nothopoda todeica n. sp. (Eriophyidae) from the Disjunct Afro-Australasian Fern Todea barbara: Morphology, Phylogeny, and Mitogenomics. Insects 2023, 14, 507. https://doi.org/10.3390/insects14060507
Chetverikov PE, Craemer C, Gankevich VD, Zhuk AS. Integrative Taxonomy of the Gall Mite Nothopoda todeica n. sp. (Eriophyidae) from the Disjunct Afro-Australasian Fern Todea barbara: Morphology, Phylogeny, and Mitogenomics. Insects. 2023; 14(6):507. https://doi.org/10.3390/insects14060507
Chicago/Turabian StyleChetverikov, Philipp E., Charnie Craemer, Vladimir D. Gankevich, and Anna S. Zhuk. 2023. "Integrative Taxonomy of the Gall Mite Nothopoda todeica n. sp. (Eriophyidae) from the Disjunct Afro-Australasian Fern Todea barbara: Morphology, Phylogeny, and Mitogenomics" Insects 14, no. 6: 507. https://doi.org/10.3390/insects14060507