
Molecular-Based Taxonomic Inferences of Some Spider Mite Species of the Genus Oligonychus Berlese (Acari, Prostigmata, Tetranychidae)


{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:Simple Summary
Abstract
1. Introduction
2. Materials and Methods
2.1. Collection, Preservation, and Processing of Spider Mite Samples
2.2. Mites Identification Based on Morphological Characters
2.3. Molecular Analysis
2.3.1. DNA Extraction and Amplification of ITS2/COI Regions
2.3.2. DNA Sequencing and Analysis
2.3.3. Phylogenetic and Genetic Distances Analyses
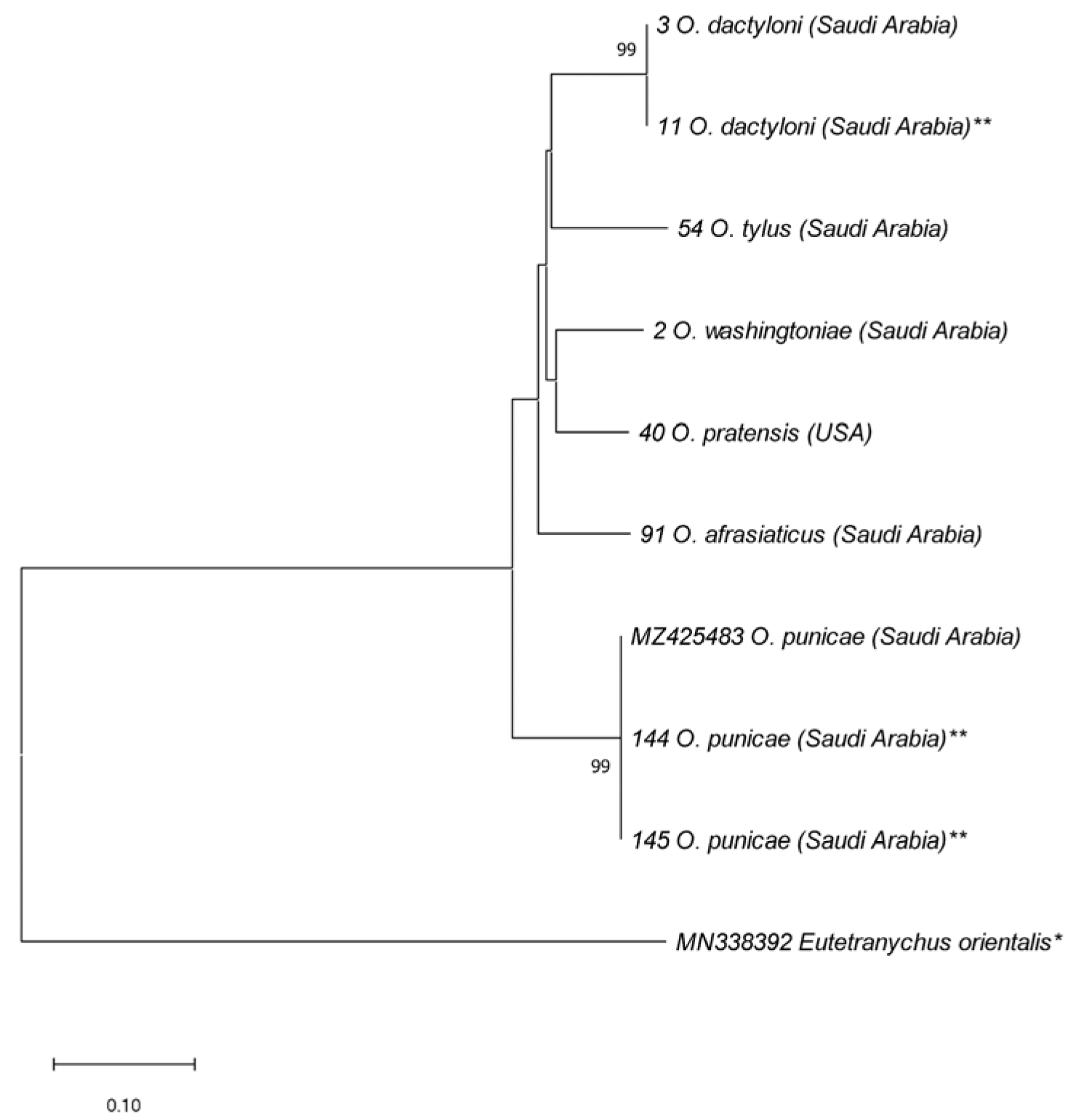
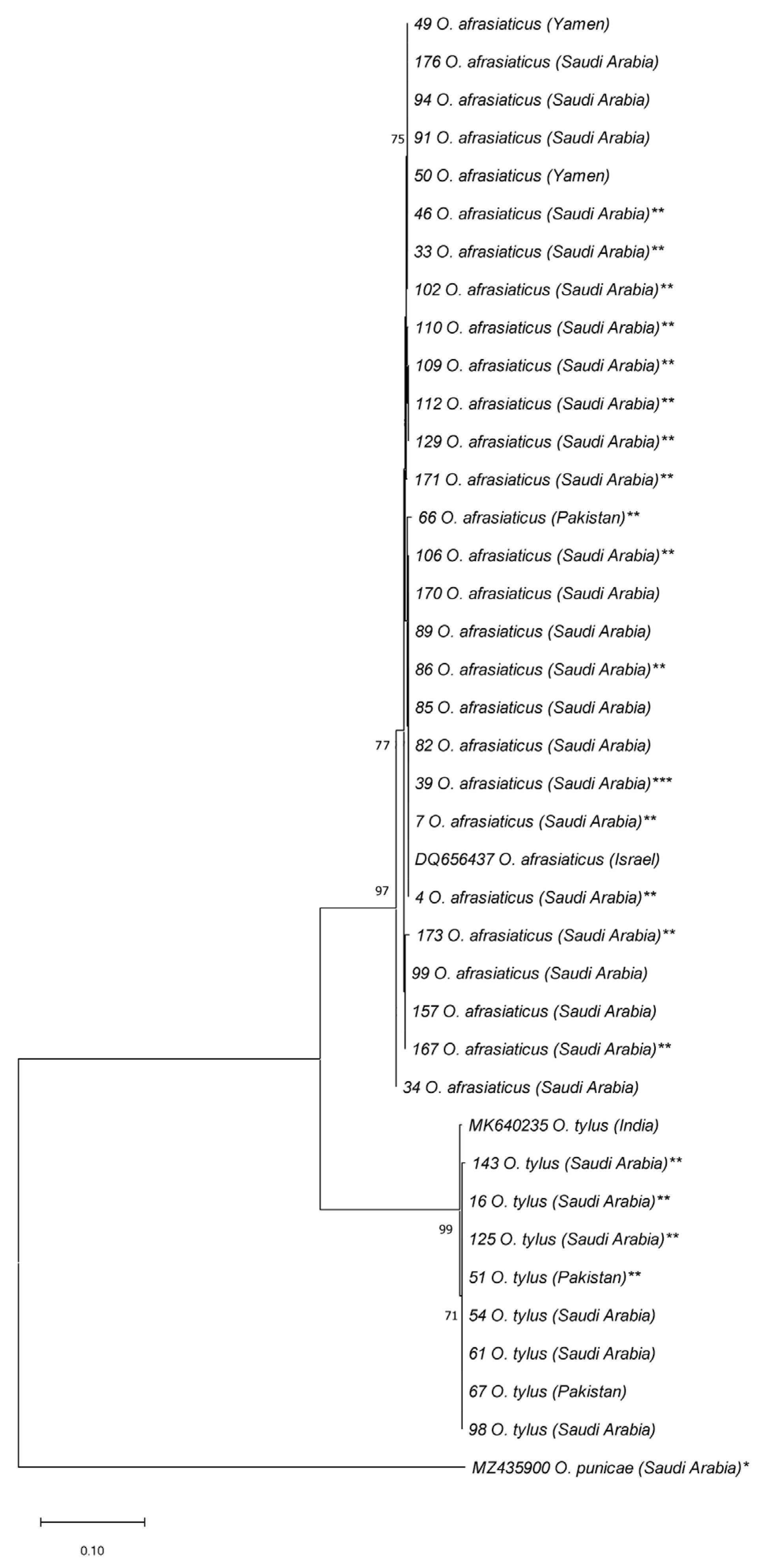
3. Results
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Pritchard, A.E.; Baker, E.W. A Revision of the Spider Mite Family Tetranychidae; Pacific Coast Entomological Society: San Francisco, CA, USA, 1955; p. 472. [Google Scholar]
- Meyer, M.K.P.S. African Tetranychidae (Acari: Prostigmata)—With Reference to the World Genera, Entomology Memoir; Department of Agriculture and Water Supply: Pretoria, South Africa, 1987; pp. 1–175.
- Mushtaq, H.M.S.; Alatawi, F.J.; Kamran, M.; Flechtmann, C.H.W. The genus Oligonychus Berlese (Acari, Prostigmata, Tetranychidae): Taxonomic assessment and a key to subgenera, species groups, and subgroups. ZooKeys 2021, 1079, 89. [Google Scholar] [CrossRef] [PubMed]
- Meyer, M.K.P.S. A Revision of the Tetranychidae of Africa (Acari) with a Key to the Genera of the World, Entomology Memoir; Department of Agricultural Technical Services: Pretoria, South Africa, 1974; pp. 1–291.
- Jeppson, L.R.; Keifer, H.H.; Baker, E.W. Mites Injurious to Economic Plants; University of California Press: Berkeley, CA, USA, 1975; pp. 1–614. [Google Scholar]
- Li, J.; Yi, T.-C.; Guo, J.-J.; Jin, D.-C. Ontogenetic development and redescription of Oligonychus pratensis (Banks, 1912) (Acari: Tetranychidae). Zootaxa 2018, 4486, 349–375. [Google Scholar] [CrossRef] [PubMed]
- Mushtaq, H.M.S.; Kamran, M.; Saleh, A.A.; Alatawi, F.J. Evidence for Reconsidering the Taxonomic Status of Closely Related Oligonychus Species in punicae Complex (Acari: Prostigmata: Tetranychidae). Insects 2023, 14, 3. [Google Scholar] [CrossRef] [PubMed]
- Rahman, K.A.; Sapra, A.N. Mites of the family Tetranychidae from Lyallpur with descriptions of four new species. Indian Acad. Sci. Proc. Sect. B 1940, 11, 177–196. [Google Scholar] [CrossRef]
- Mitrofanov, V.I.; Zapletina, A. A new species of the genus Oligonychus from Azerbaijan. Dokl. Akad. Nauk. Azerbaijan SSR 1973, 29, 50–52. [Google Scholar]
- Meyer, M.K.P.; Ryke, P.A.J. A revision of the spider mites (Acarina: Tetranychidae) of South Africa with descriptions of a new genus and new species. J. Entomol. Soc. S. Afr. 1959, 22, 330–366. [Google Scholar]
- Tseng, Y.H. Mites associated with the conifers of Taiwan, (1) Tetranychoidea, with a checklist of the world. J. Entomol. Natl. Chung Hsien Univ. 1980, 15, 145–168. [Google Scholar]
- Navajas, M.; Gutierrez, J.; Lagnel, J.; Boursot, P. Mitochondrial cytochrome oxidase I in tetranychid mites: A comparison between molecular phylogeny and changes of morphological and life history traits. Bull. Entomol. Res. 1996, 86, 407–417. [Google Scholar] [CrossRef] [Green Version]
- Matsuda, T.; Fukumoto, C.; Hinomoto, N.; Gotoh, T. DNA-based identification of spider mites: Molecular evidence for cryptic species of the genus Tetranychus (Acari: Tetranychidae). J. Econ. Entomol. 2013, 106, 463–472. [Google Scholar] [CrossRef] [Green Version]
- Matsuda, T.; Hinomoto, N.; Singh, R.N.; Gotoh, T. Molecular-based identification and phylogeny of Oligonychus species (Acari: Tetranychidae). J. Econ. Entomol. 2012, 105, 1043–1050. [Google Scholar] [CrossRef] [Green Version]
- Arabuli, T.; Gotoh, T. A new species of spider mite, Oligonychus neocastaneae sp. nov. (Acari: Tetranychidae), from Japan. Zootaxa 2018, 4378, 563–572. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ben-David, T.; Melamed, S.; Gerson, U.; Morin, S. ITS2 sequences as barcodes for identifying and analyzing spider mites (Acari: Tetranychidae). Exp. Appl. Acarol. 2007, 41, 169–181. [Google Scholar] [CrossRef] [PubMed]
- Mirza, J.H.; Kamran, M.; Saleh, A.A.; Alatawi, F.J. Molecular and phenotypic variations in Eutetranychus orientalis (Klein) populations from Saudi Arabia. PLoS ONE 2020, 15, e0233389. [Google Scholar] [CrossRef] [PubMed]
- Navajas, M.; Gutierrez, J.; Williams, M.; Gotoh, T. Synonymy between two spider mite species, Tetranychus kanzawai and T. hydrangeae (Acari: Tetranychidae), shown by ribosomal ITS2 sequences and cross-breeding experiments. Bull. Entomol. Res. 2001, 91, 117–123. [Google Scholar] [PubMed]
- Gotoh, T.; Araki, R.; Boubou, A.; Migeon, A.; Ferragut, F.; Navajas, M. Evidence of co-specificity between Tetranychus evansi and Tetranychus takafujii (Acari: Prostigmata, Tetranychidae): Comments on taxonomic and agricultural aspects. Int. J. Acarol. 2009, 35, 485–501. [Google Scholar] [CrossRef]
- Zeity, M.; Srinivasa, N.; Gowda, C.C. Are Tetranychus macfarlanei Baker & Pritchard and Tetranychus malaysiensis Ehara (Acari: Tetranychidae) one species? Morphological and molecular evidences for synonymy between these two spider mite species and a note on invasiveness of T. macfarlanei on okra and eggplant in India. Syst. Appl. Acarol. 2017, 22, 467–476. [Google Scholar] [CrossRef]
- Mushtaq, H.M.S.; Kamran, M.; Alatawi, F.J. New species, new records, and re-descriptions of two species of the genus Oligonychus Berlese (Acari: Prostigmata: Tetranychidae) from Saudi Arabia. Syst. Appl. Acarol. 2022, 27, 2568–2582. [Google Scholar] [CrossRef]
- Migeon, A.; Dorkeld, F. Spider Mites Web: A Comprehensive Database for the Tetranychidae. Available online: http://www1.montpellier.inra.fr/CBGP/spmweb (accessed on 5 January 2023).
- Alatawi, F.J.; Kamran, M. Spider mites (Acari: Tetranychidae) of Saudi Arabia: Two new species, new records and a key to all known species. J. Nat. Hist. 2018, 52, 429–455. [Google Scholar] [CrossRef]
- Guzman-Valencia, S.; Santillán-Galicia, M.T.; Guzmán-Franco, A.W.; González-Hernández, H.; Carrillo-Benítez, M.G.; Suárez-Espinoza, J. Contrasting effects of geographical separation on the genetic population structure of sympatric species of mites in avocado orchards. Bull. Entomol. Res. 2014, 104, 610–621. [Google Scholar] [CrossRef]
- Ganjisaffar, F.; Perring, T.M. Prey stage preference and functional response of the predatory mite Galendromus flumenis to Oligonychus pratensis. Biol. Control 2015, 82, 40–45. [Google Scholar] [CrossRef]
- Bolland, H.R.; Gutierrez, J.; Flechtmann, C.H.W. World Catalogue of the Spider Mite Family (Acari: Tetranychidae); Brill Academic Publishers: Leiden, The Netherlands, 1998; p. 392. [Google Scholar]
- Mushtaq, H.M.S.; Alatawi, F.J.; Kamran, M. Keys to World Species of the Genus Oligonychus Berlese (Acari, Prostigmata, Tetranychidae): Taxonomic Notes and New Oligonychus Species, King Saud University: Riyadh, Saudi Arabia, 2023; in prepration.
- Hall, T.A. BioEdit: A user-friendly biological sequence alignment editor and analysis program for Windows 95/98/NT. Proc. Nucleic Acids Symp. Ser. 1999, 41, 95–98. [Google Scholar]
- Kumar, S.; Stecher, G.; Li, M.; Knyaz, C.; Tamura, K. MEGA X: Molecular evolutionary genetics analysis across computing platforms. Mol. Biol. Evol. 2018, 35, 1547–1549. [Google Scholar] [CrossRef] [PubMed]
- Tamura, K.; Nei, M. Estimation of the number of nucleotide substitutions in the control region of mitochondrial DNA in humans and chimpanzees. Mol. Biol. Evol. 1993, 10, 512–526. [Google Scholar]
- Felsenstein, J. Confidence limits on phylogenies: An approach using the bootstrap. Evolution 1985, 39, 783–791. [Google Scholar] [CrossRef] [PubMed]
- Navajas, M.; Gutierrez, J.; Bonato, O.; Bolland, H.R.; Mapangou-Divassa, S. Intraspecific diversity of the cassava green mite Mononychellus progresivus (Acari: Tetranychidae) using comparisons of mitochondrial and nuclear ribosomal DNA sequences and cross-breeding. Exp. Appl. Acarol. 1994, 18, 351–360. [Google Scholar] [CrossRef] [PubMed]
- Khaing, T.M.; Shim, J.K.; Lee, K.Y. Molecular identification of four Panonychus species (Acari: Tetranychidae) in Korea, including new records of P. caglei and P. mori. Entomol. Res. 2015, 45, 345–353. [Google Scholar] [CrossRef]
- Yang, B.; Cai, J.; Cheng, X. Identification of astigmatid mites using ITS2 and COI regions. Parasitol. Res. 2011, 108, 497–503. [Google Scholar] [CrossRef]
- Navia, D.; Mendonca, R.S.; Ferragut, F.; Miranda, L.C.; Trincado, R.C.; Michaux, J.; Navajas, M. Cryptic diversity in Brevipalpus mites (Tenuipalpidae). Zool. Scr. 2013, 42, 406–426. [Google Scholar] [CrossRef]
- Ma, E.P.; Yuan, Y.L. New species and new records of tetranychid mites from China I. (Acarina: Tetranychidae). Acta Zootaxonomica Sin. 1980, 5, 42–45. [Google Scholar]
- Sivakumar, K.; Kunchithapatham, R. Phytophagous mite diversity of Tetranychus spp. and Oligonychus spp. (Acari: Tetranychidae) found on different host plants in Coimbatore district and surrounding regions. Trends Biosci. 2014, 7, 4113–4117. [Google Scholar]
- Chaudhri, W.M.; Akbar, S.; Rasool, A. Taxonomic Studies of the Mites Belonging to the Families Tenuipalpidae, Tetranychidae, Tuckerellidae, Caligonellidae, Stigmaeidae and Phytoseiidae, PL-480 Project on Mites; University of Agriculture: Lyallpur, Pakistan, 1974; p. 250. [Google Scholar]
- Xie, L.; Hong, X.-y.; Xue, X.-f. Population genetic structure of the two spotted spider mite (Acari: Tetranychidae) from China. Ann. Entomol. Soc. Am. 2006, 99, 959–965. [Google Scholar] [CrossRef]
- Ge, M.-K.; Sun, E.-T.; Jia, C.-N.; Kong, D.-D.; Jiang, Y.-X. Genetic diversity and differentiation of Lepidoglyphus destructor (Acari: Glycyphagidae) inferred from inter-simple sequence repeat (ISSR) fingerprinting. Syst. Appl. Acarol. 2014, 19, 491–498. [Google Scholar] [CrossRef]
- Yuan, M.-L.; Wei, D.-D.; Zhang, K.; Gao, Y.-Z.; Liu, Y.-H.; Wang, B.-J.; Wang, J.-J. Genetic Diversity and Population Structure of Panonychus citri (Acari: Tetranychidae), in China Based on Mitochondrial COI Gene Sequences. Mol. Entomol. 2010, 103, 2204–2213. [Google Scholar] [CrossRef] [PubMed]
- Ros, V.I.D.; Breeuwer, J.A.J. Spider mite (Acari: Tetranychidae) mitochondrial COI phylogeny reviewed: Host plant relationships, phylogeography, reproductive parasites and barcoding. Exp. Appl. Acarol. 2007, 42, 239–262. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Matsuda, T.; Kozaki, T.; Ishii, K.; Gotoh, T. Phylogeny of the spider mite sub-family Tetranychinae (Acari: Tetranychidae) inferred from RNA-Seq data. PLoS ONE 2018, 13, e0203136. [Google Scholar] [CrossRef] [PubMed]








Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Mushtaq, H.M.S.; Saleh, A.A.; Kamran, M.; Alatawi, F.J. Molecular-Based Taxonomic Inferences of Some Spider Mite Species of the Genus Oligonychus Berlese (Acari, Prostigmata, Tetranychidae). Insects 2023, 14, 192. https://doi.org/10.3390/insects14020192
Mushtaq HMS, Saleh AA, Kamran M, Alatawi FJ. Molecular-Based Taxonomic Inferences of Some Spider Mite Species of the Genus Oligonychus Berlese (Acari, Prostigmata, Tetranychidae). Insects. 2023; 14(2):192. https://doi.org/10.3390/insects14020192
Chicago/Turabian StyleMushtaq, Hafiz Muhammad Saqib, Amgad A. Saleh, Muhammad Kamran, and Fahad Jaber Alatawi. 2023. "Molecular-Based Taxonomic Inferences of Some Spider Mite Species of the Genus Oligonychus Berlese (Acari, Prostigmata, Tetranychidae)" Insects 14, no. 2: 192. https://doi.org/10.3390/insects14020192