
Phylogeny and Evolutionary Timescale of Muscidae (Diptera: Calyptratae) Inferred from Mitochondrial Genomes

 , , ,
, , ,
Abstract
:Simple Summary
Abstract
1. Introduction
2. Materials and Methods
2.1. Sampling and DNA Extraction
2.2. Sequences Assembly and Annotation
2.3. Phylogeny Analyses
2.4. Divergence Time Estimation
3. Results
3.1. Mtgenome Assembly
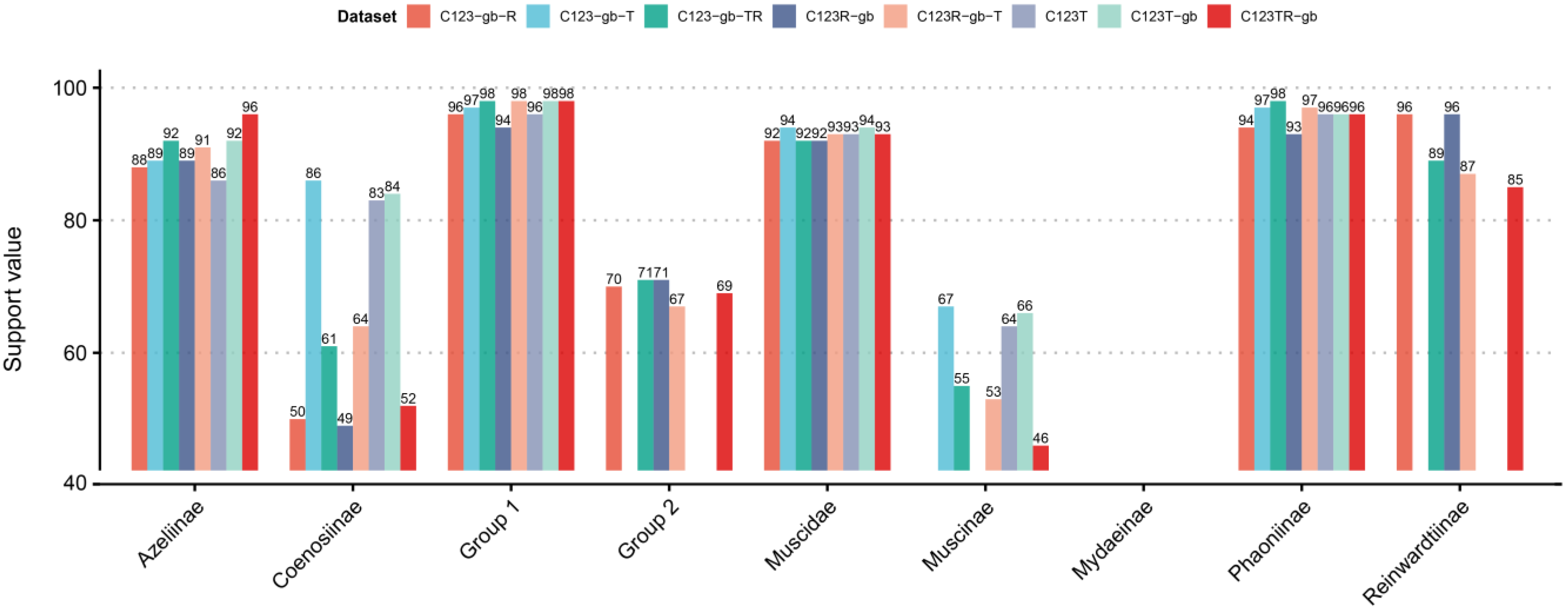
3.2. Phylogenetic Effect of RNA Sequences
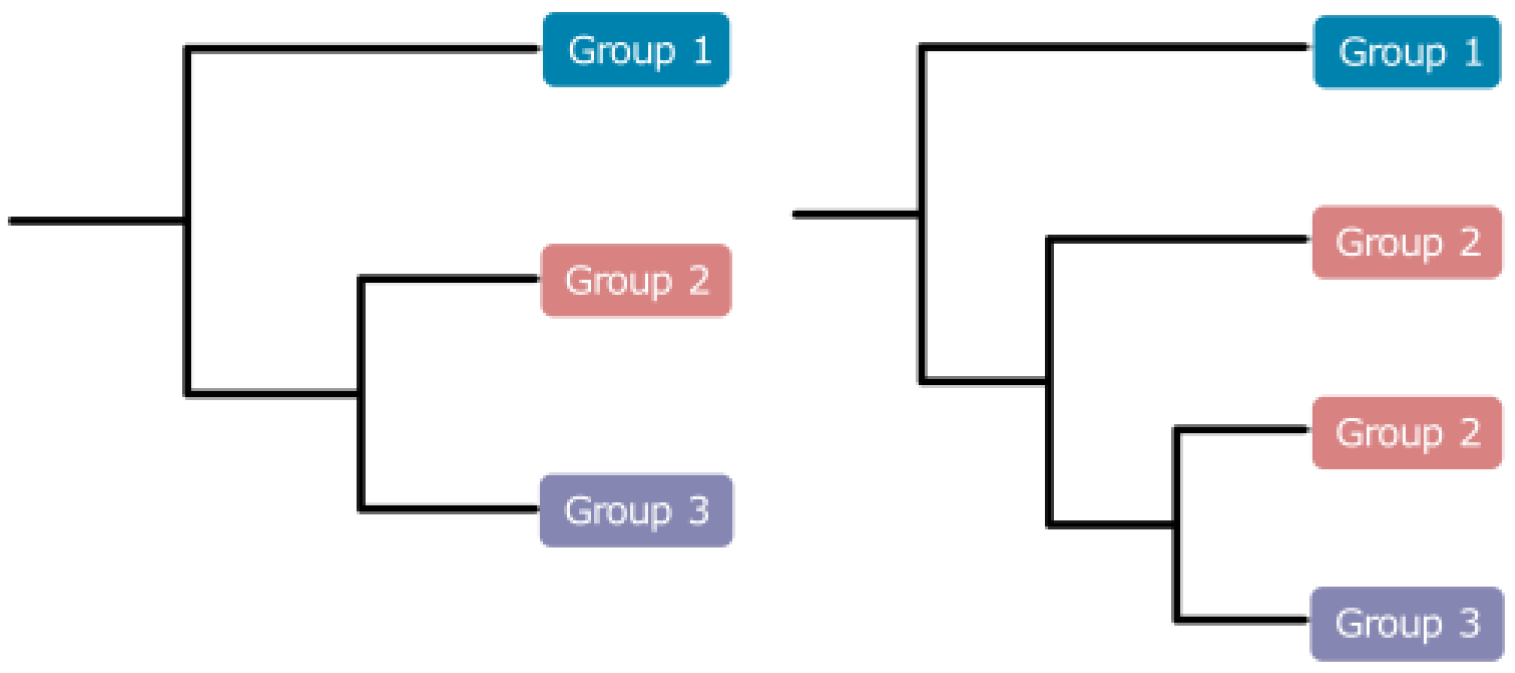
3.3. Phylogenetic Analyses of Muscidae
3.3.1. Phylogenetic Analyses Based on IQ-Tree
3.3.2. The Best Phylogenetic Tree
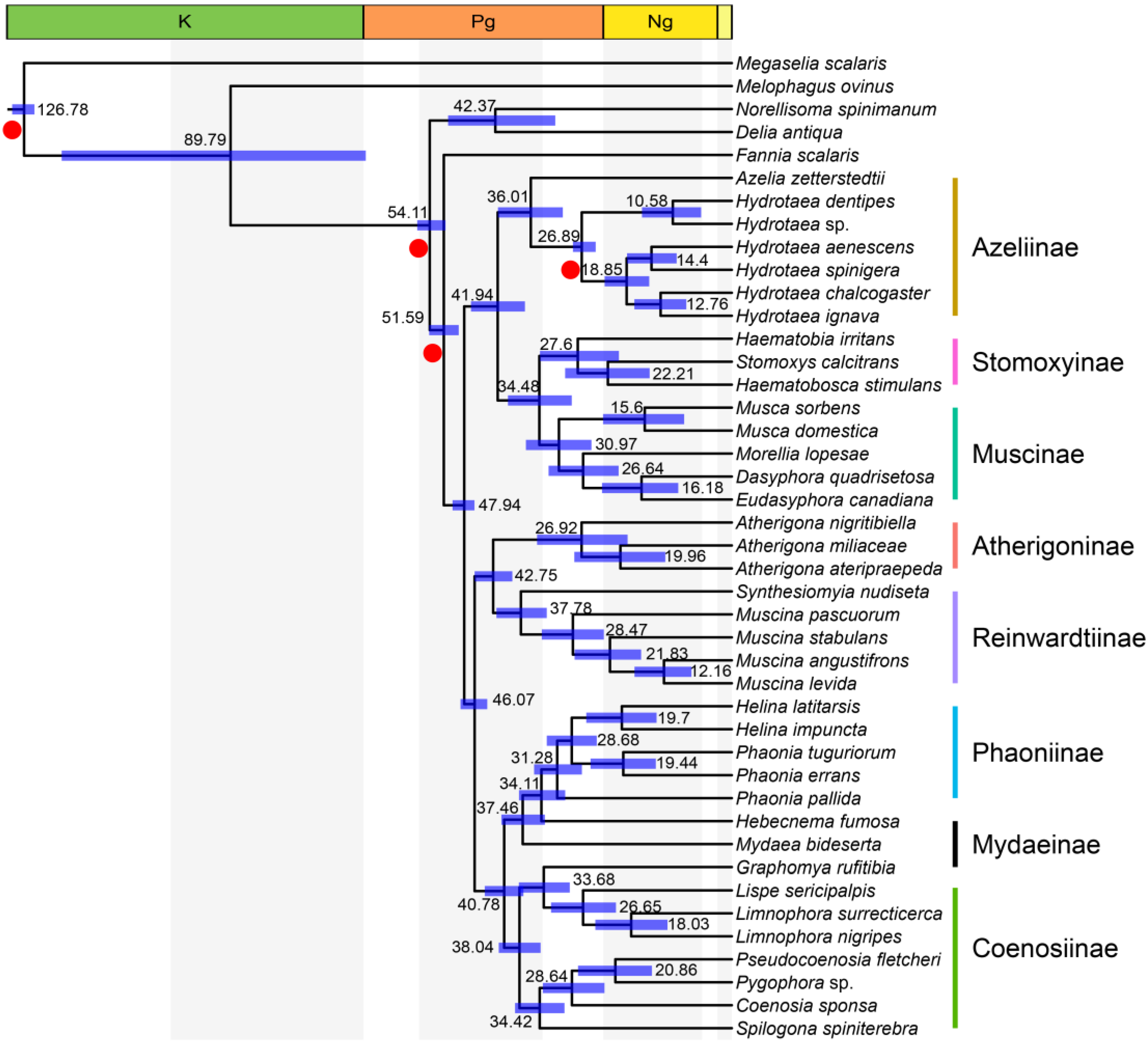
3.4. Time Frame Based on Mtgenomes
4. Discussion
Supplementary Materials
Author Contributions
Funding
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Bánki, O.; Roskov, Y.; Döring, M.; Ower, G.; Vandepitte, L.; Hobern, D.; Remsen, D.; Schalk, P.; DeWalt, R.E.; Keping, M.; et al. Catalogue of Life Checklist, version 21 March 2022; Catalogue of Life: Leiden, The Netherlands, 2022. [CrossRef]
- Ivorra, T.; Martínez-Sánchez, A.; Rojo, S. Review of Synthesiomyia nudiseta (Diptera: Muscidae) as a useful tool in forensic entomology. Int. J. Legal Med. 2021, 135, 2003–2015. [Google Scholar] [CrossRef] [PubMed]
- Skidmore, P. The Biology of the Muscidae of the World; Series Entomologica; Dr W. Junk Publishers: Dordrecht, The Netherlands, 1985; Volume 29, pp. 1–550. [Google Scholar]
- Grzywacz, A.; Hall, M.J.R.; Pape, T.; Szpila, K. Muscidae (Diptera) of forensic importance—An identification key to third instar larvae of the western Palaearctic region and a catalogue of the muscid carrion community. Int. J. Legal Med. 2017, 131, 855–866. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Brown, B.V.; Borkent, A.; Cumming, J.M.; Wood, D.M.; Woodley, N.E.; Zumbado, M.A. Manual of Central American Diptera; NRC Research Press: Ottawa, ON, Canada, 2010; Volume 2, pp. 715–1442. [Google Scholar]
- Marchiori, C.H. Study of the biology and ecology of the Muscidae Family (Insecta: Diptera): Collections. Open Access Res. J. Multidiscip. Stud. 2021, 2, 108–116. [Google Scholar] [CrossRef]
- Cupp, E.W.; Cupp, M.S.; Ribeiro, J.M.C.; Kunz, S.E. Blood-feeding strategy of Haematobia irritans (Diptera: Muscidae). J. Med. Entomol. 1998, 35, 591–595. [Google Scholar] [CrossRef]
- Fessl, B.; Sinclair, B.J.; Kleindorfer, S. The life-cycle of Philornis downsi (Diptera: Muscidae) parasitizing Darwin’s finches and its impacts on nestling survival. Parasitology 2006, 133, 739–747. [Google Scholar] [CrossRef]
- Al-Keri, L.A.; Ghoneim, K. Evaluation of the toxic effects of novaluron on Muscina stabulans (Fallen) (Diptera: Muscidae). Pak. J. Biol. Sci. 2021, 24, 42–52. [Google Scholar] [CrossRef]
- Toto, N.A.; Elhenawy, H.I.; Eltaweil, A.S.; El-Ashram, S.; El-Samad, L.M.; Moussian, B.; El Wakil, A. Musca domestica (Diptera: Muscidae) as a biological model for the assessment of magnetite nanoparticles toxicity. Sci. Total Environ. 2022, 806, 151483. [Google Scholar] [CrossRef]
- Carvalho, C.J.B.D. Classificação de Muscidae (Diptera): Uma proposta através da análise cladístical. Rev. Bras. Zool. 1989, 6, 627–648. [Google Scholar] [CrossRef] [Green Version]
- Xue, W.; Chao, C. Flies of China; Liaoning Science and Technology Press: Shenyang, China, 1998; Volume 1, pp. 1–1365. [Google Scholar]
- Carvalho, C.J.B.D.; Couri, M.S.; Pont, A.C.; Pamplona, D.; Lopes, S.M. A Catalogue of the Muscidae (Diptera) of the Neotropical Region. Zootaxa 2005, 860, 1–282. [Google Scholar] [CrossRef]
- Fan, Z.D. Diptera Muscidae (I); Science Press: Beijing, China, 2008; Volume 49, pp. 1–1186. [Google Scholar]
- Grzywacz, A.; Wallman, J.F.; Piwczyński, M. To be or not to be a valid genus: The systematic position of Ophyra R.-D. revised (Diptera: Muscidae): The systematic position of Ophyra revised. Syst. Entomol. 2017, 42, 714–723. [Google Scholar] [CrossRef] [Green Version]
- Haseyama, K.L.F.; Wiegmann, B.M.; Almeida, E.A.B.; de Carvalho, C.J.B. Say goodbye to tribes in the new house fly classification: A new molecular phylogenetic analysis and an updated biogeographical narrative for the Muscidae (Diptera). Mol. Phylogenet. Evol. 2015, 89, 1–12. [Google Scholar] [CrossRef] [PubMed]
- Grzywacz, A.; Trzeciak, P.; Wiegmann, B.M.; Cassel, B.K.; Pape, T.; Walczak, K.; Bystrowski, C.; Nelson, L.; Piwczyński, M. Towards a new classification of Muscidae (Diptera): A comparison of hypotheses based on multiple molecular phylogenetic approaches. Syst. Entomol. 2021, 46, 508–525. [Google Scholar] [CrossRef]
- Kutty, S.N.; Pont, A.C.; Meier, R.; Pape, T. Complete tribal sampling reveals basal split in Muscidae (Diptera), confirms saprophagy as ancestral feeding mode, and reveals an evolutionary correlation between instar numbers and carnivory. Mol. Phylogenet. Evol. 2014, 78, 349–364. [Google Scholar] [CrossRef]
- Schuehli, G.S.; Carvalho, C.J.B.D.; Wiegmann, B.M. Molecular phylogenetics of the Muscidae (Diptera:Calyptratae): New ideas in a congruence context. Invertebr. Syst. 2007, 21, 263–278. [Google Scholar] [CrossRef]
- Bae, J.S.; Kim, I.; Sohn, H.D.; Jin, B.R. The mitochondrial genome of the firefly, Pyrocoelia rufa: Complete DNA sequence, genome organization, and phylogenetic analysis with other insects. Mol. Phylogenet. Evol. 2004, 32, 978–985. [Google Scholar] [CrossRef] [PubMed]
- Wang, Y.; Liu, X.; Winterton, S.L.; Yang, D. The first mitochondrial genome for the fishfly subfamily Chauliodinae and implications for the higher phylogeny of Megaloptera. PLoS ONE 2012, 7, e47302. [Google Scholar] [CrossRef] [Green Version]
- Ding, S.; Li, X.; Wang, N.; Cameron, S.L.; Mao, M.; Wang, Y.; Xi, Y.; Yang, D. The phylogeny and evolutionary timescale of Muscoidea (Diptera: Brachycera: Calyptratae) inferred from mitochondrial genomes. PLoS ONE 2015, 10, e0134170. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ren, L.; Shang, Y.; Yang, L.; Shen, X.; Chen, W.; Wang, Y.; Cai, J.; Guo, Y. Comparative analysis of mitochondrial genomes among four species of muscid flies (Diptera: Muscidae) and its phylogenetic implications. Int. J. Biol. Macromol. 2019, 127, 357–364. [Google Scholar] [CrossRef]
- Song, F.; Li, H.; Jiang, P.; Zhou, X.; Liu, J.; Sun, C.; Vogler, A.P.; Cai, W. Capturing the phylogeny of Holometabola with mitochondrial genome data and bayesian site-heterogeneous mixture models. Genome Biol. Evol. 2016, 8, 1411–1426. [Google Scholar] [CrossRef]
- Cameron, S.L.; Lambkin, C.L.; Barker, S.C.; Whiting, M.F. A mitochondrial genome phylogeny of Diptera: Whole genome sequence data accurately resolve relationships over broad timescales with high precision. Syst. Entomol. 2007, 32, 40–59. [Google Scholar] [CrossRef]
- Boore, J.L. Animal mitochondrial genomes. Nucleic Acids Res. 1999, 27, 1767–1780. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Cameron, S.L. Insect mitochondrial genomics: Implications for evolution and phylogeny. Annu. Rev. Entomol. 2014, 59, 95–117. [Google Scholar] [CrossRef] [Green Version]
- Zhang, N.; Yu, G.; Li, T.; He, Q.; Zhou, Y.; Si, F.; Ren, S.; Chen, B. The complete mitochondrial genome of Delia antiqua and its implications in dipteran phylogenetics. PLoS ONE 2015, 10, e0139736. [Google Scholar] [CrossRef] [PubMed]
- Ren, L.; Zhang, X.; Li, Y.; Shang, Y.; Chen, S.; Wang, S.; Qu, Y.; Cai, J.; Guo, Y. Comparative analysis of mitochondrial genomes among the subfamily Sarcophaginae (Diptera: Sarcophagidae) and phylogenetic implications. Int. J. Biol. Macromol. 2020, 161, 214–222. [Google Scholar] [CrossRef]
- Zhong, M.; Wang, X.; Liu, Q.; Luo, B.; Wu, C.; Wen, J. The complete mitochondrial genome of the scuttle fly, Megaselia scalaris (Diptera: Phoridae). Mitochondrial DNA Part A DNA Mapp. Seq. Anal. 2016, 27, 182–184. [Google Scholar] [CrossRef] [PubMed]
- Tang, J.M.; Li, F.; Cheng, T.Y.; Duan, D.Y.; Liu, G.H. Comparative analyses of the mitochondrial genome of the sheep ked Melophagus ovinus (Diptera: Hippoboscidae) from different geographical origins in China. Parasitol. Res. 2018, 117, 2677–2683. [Google Scholar] [CrossRef]
- Zhang, J.; Deng, S. The complete mitochondrial genome of Hydrotaea (Ophyra) chalcogaster (Diptera: Muscidae). Mitochondrial DNA Part B Resour. 2018, 3, 961. [Google Scholar] [CrossRef]
- Yan, W.; Shang, Y.; Ren, L.; Zhang, X.; Wang, Y. The complete mitochondrial genome of Hydrotaea dentipes (Diptera: Muscidae). Mitochondrial DNA Part B Resour. 2019, 4, 2044–2045. [Google Scholar] [CrossRef] [Green Version]
- Karagozlu, M.Z.; Park, S.H.; Shin, S.-E.; Kim, C.-B. Complete mitochondrial DNA analysis of Hydrotaea ignava (Insecta, Diptera, Muscidae). Mitochondrial DNA Part B Resour. 2017, 2, 825–826. [Google Scholar] [CrossRef] [Green Version]
- Junqueira, A.C.M.; Azeredo-Espin, A.M.L.; Paulo, D.F.; Marinho, M.A.T.; Tomsho, L.P.; Drautz-Moses, D.I.; Purbojati, R.W.; Ratan, A.; Schuster, S.C. Large-scale mitogenomics enables insights into Schizophora (Diptera) radiation and population diversity. Sci. Rep. 2016, 6, 21762. [Google Scholar] [CrossRef] [Green Version]
- Karagozlu, M.Z.; Park, S.H.; Shin, S.-E.; Kim, C.-B. Sequencing and analyzing complete mitochondrial DNA of Muscina angustifrons (Insecta, Diptera, Muscidae). Mitochondrial DNA Part B Resour. 2017, 2, 115–116. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Huang, J.; Ni, Z.; Wang, H.; Zhang, L.; Yan, J.; Bai, S. The complete mitochondrial genome of Muscina pascuorum (Diptera: Muscidae). Mitochondrial DNA Part B Resour. 2021, 6, 110–111. [Google Scholar] [CrossRef] [PubMed]
- Lan, L.; Liu, Y.; Yan, J.; Lin, L.; Zha, L. The complete mitochondrial genome of the flesh fly, Muscina stabulans (Diptera: Muscidae). Mitochondrial DNA Part A DNA Mapp. Seq. Anal. 2016, 27, 4069–4070. [Google Scholar] [CrossRef]
- Li, X.; Wang, Y.; Su, S.; Yang, D. The complete mitochondrial genomes of Musca domestica and Scathophaga stercoraria (Diptera: Muscoidea: Muscidae and Scathophagidae). Mitochondrial DNA Part A DNA Mapp. Seq. Anal. 2016, 27, 1435–1436. [Google Scholar]
- Ma, T.; Huang, J. The complete mitochondrial genome of the bazaar fly, Musca sorbens Wiedemann (Diptera: Muscidae). Mitochondrial DNA Part B Resour. 2018, 3, 436–437. [Google Scholar] [CrossRef] [Green Version]
- Chen, W.; Shang, Y.; Ren, L.; Zhang, X.; Guo, Y. The complete mitochondrial genome of Graphomya rufitibia (Diptera: Muscidae). Mitochondrial DNA Part B Resour. 2018, 3, 403–404. [Google Scholar] [CrossRef] [Green Version]
- Donath, A.; Jühling, F.; Al-Arab, M.; Bernhart, S.H.; Reinhardt, F.; Stadler, P.F.; Middendorf, M.; Bernt, M. Improved annotation of protein-coding genes boundaries in metazoan mitochondrial genomes. Nucleic Acids Res. 2019, 47, 10543–10552. [Google Scholar] [CrossRef]
- Kearse, M.; Moir, R.; Wilson, A.; Stones-Havas, S.; Cheung, M.; Sturrock, S.; Buxton, S.; Cooper, A.; Markowitz, S.; Duran, C.; et al. Geneious Basic: An integrated and extendable desktop software platform for the organization and analysis of sequence data. Bioinformatics 2012, 28, 1647–1649. [Google Scholar] [CrossRef] [Green Version]
- Lowe, T.M.; Chan, P.P. tRNAscan-SE On-line: Integrating search and context for analysis of transfer RNA genes. Nucleic Acids Res. 2016, 44, 54–57. [Google Scholar] [CrossRef]
- Laslett, D.; Canback, B. ARWEN: A program to detect tRNA genes in metazoan mitochondrial nucleotide sequences. Bioinformatics 2008, 24, 172–175. [Google Scholar] [CrossRef] [Green Version]
- Cameron, S.L. How to sequence and annotate insect mitochondrial genomes for systematic and comparative genomics research. Syst. Entomol. 2014, 39, 400–411. [Google Scholar] [CrossRef] [Green Version]
- Zhang, D.; Gao, F.; Jakovlić, I.; Zou, H.; Zhang, J.; Li, W.X.; Wang, G.T. PhyloSuite: An integrated and scalable desktop platform for streamlined molecular sequence data management and evolutionary phylogenetics studies. Mol. Ecol. Resour. 2020, 20, 348–355. [Google Scholar] [CrossRef] [PubMed]
- Katoh, K.; Standley, D.M. MAFFT multiple sequence alignment software version 7: Improvements in performance and usability. Mol. Biol. Evol. 2013, 30, 772–780. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Castresana, J. Selection of conserved blocks from multiple alignments for their use in phylogenetic analysis. Mol. Biol. Evol. 2000, 17, 540–552. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lanfear, R.; Frandsen, P.B.; Wright, A.M.; Senfeld, T.; Calcott, B. PartitionFinder 2: New methods for selecting partitioned models of evolution for molecular and morphological phylogenetic analyses. Mol. Biol. Evol. 2017, 34, 772–773. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Nguyen, L.-T.; Schmidt, H.A.; von Haeseler, A.; Minh, B.Q. IQ-TREE: A fast and effective stochastic algorithm for estimating maximum-likelihood phylogenies. Mol. Biol. Evol. 2015, 32, 268–274. [Google Scholar] [CrossRef]
- Minh, B.Q.; Nguyen, M.A.T.; von Haeseler, A. Ultrafast Approximation for Phylogenetic Bootstrap. Mol. Biol. Evol. 2013, 30, 1188–1195. [Google Scholar] [CrossRef] [Green Version]
- Guindon, S.; Dufayard, J.-F.; Lefort, V.; Anisimova, M.; Hordijk, W.; Gascuel, O. New algorithms and methods to estimate maximum-likelihood phylogenies: Assessing the performance of PhyML 3.0. Syst. Biol. 2010, 59, 307–321. [Google Scholar] [CrossRef] [Green Version]
- Letunic, I.; Bork, P. Interactive tree of life (iTOL) v5: An online tool for phylogenetic tree display and annotation. Nucleic Acids Res. 2021, 49, 293–296. [Google Scholar] [CrossRef]
- Yang, Z. PAML 4: Phylogenetic analysis by maximum likelihood. Mol. Biol. Evol. 2007, 24, 1586–1591. [Google Scholar] [CrossRef] [Green Version]
- Wang, L.; Lam, T.T.-Y.; Xu, S.; Dai, Z.; Zhou, L.; Feng, T.; Guo, P.; Dunn, C.W.; Jones, B.R.; Bradley, T.; et al. Treeio: An R package for phylogenetic tree input and output with richly annotated and associated data. Mol. Biol. Evol. 2020, 37, 599–603. [Google Scholar] [CrossRef] [PubMed]
- Yu, G.; Smith, D.K.; Zhu, H.; Guan, Y.; Lam, T.T. ggtree: An R package for visualization and annotation of phylogenetic trees with their covariates and other associated data. Methods Ecol. Evol. 2017, 8, 28–36. [Google Scholar] [CrossRef]
- Gearty, W.; AEgit; Yu, G.; Guidoni, K. Deeptime. Available online: https://github.com/willgearty/deeptime (accessed on 26 November 2021).
- Mo, R.-R.; Wang, Y.; Cao, J.-J.; Wang, G.-Q.; Li, W.-H.; Murányi, D. Two complete mitochondrial genomes of the subfamily Chloroperlinae (Plecoptera: Chloroperlidae) and their phylogenetic implications. Arthropod Syst. Phylog. 2022, 80, 155–168. [Google Scholar] [CrossRef]
- Li, X.; Wang, L.; Li, Z.; Hou, P.; Yang, D. The mitochondrial genome of Formicosepsis sp. (Diptera: Cypselosomatidae). Mitochondrial DNA Part B Resour. 2019, 4, 2140–2141. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Brandl, R.; Mann, W.; Sprinzl, M. Mitochondrial tRNA and the phylogenetic position of nematoda. Biochem. Syst. Ecol. 1992, 20, 325–330. [Google Scholar] [CrossRef]
- Song, S.-N.; Tang, P.; Wei, S.-J.; Chen, X.-X. Comparative and phylogenetic analysis of the mitochondrial genomes in basal hymenopterans. Sci. Rep. 2016, 6, 20972. [Google Scholar] [CrossRef] [Green Version]
- Kim, H.; Lee, S. A molecular phylogeny of the tribe Aphidini (Insecta: Hemiptera: Aphididae) based on the mitochondrial tRNA/COII, 12S/16S and the nuclear EF1α genes. Syst. Entomol. 2008, 33, 711–721. [Google Scholar] [CrossRef]
- Couri, M.S.; Carvalho, C.J.B. Systematic relations among Philornis Meinert, Passeromyia Rodhain & Villeneuve and allied genera (Diptera, Muscidae). Braz. J. Biol. 2003, 63, 223–232. [Google Scholar]
- Kutty, S.N.; Pape, T.; Pont, A.; Wiegmann, B.M.; Meier, R. The Muscoidea (Diptera: Calyptratae) are paraphyletic: Evidence from four mitochondrial and four nuclear genes. Mol. Phylogenet. Evol. 2008, 49, 639–652. [Google Scholar] [CrossRef]
- Kutty, S.N.; Pape, T.; Wiegmann, B.M.; Meier, R. Molecular phylogeny of the Calyptratae (Diptera: Cyclorrhapha) with an emphasis on the superfamily Oestroidea and the position of Mystacinobiidae and McAlpine’s fly. Syst. Entomol. 2010, 35, 614–635. [Google Scholar] [CrossRef]
- Kutty, S.N.; Meusemann, K.; Bayless, K.M.; Marinho, M.A.T.; Pont, A.C.; Zhou, X.; Misof, B.; Wiegmann, B.M.; Yeates, D.; Cerretti, P.; et al. Phylogenomic analysis of Calyptratae: Resolving the phylogenetic relationships within a major radiation of Diptera. Cladistics 2019, 35, 605–622. [Google Scholar] [CrossRef]
- Gregor, F.; Rozkošný, R.; Bartak, M.; Vaňhara, J. The Muscidae (Diptera) of Central Europe. Folia Facultatis Scientiarum Naturalium Universitatis Masarykianae Brunensis; Masaryk University: Brno, Czech Republic, 2002; Volume Biologia 107, pp. 1–280. [Google Scholar]
- Hennig, W. Vorarbeiten zu einem phylogenetischen system der Muscidae (Diptera: Cyclorrhapha). Stuttg. Beiträge Nat. 1965, 141, 1–100. [Google Scholar]
- Pont, A.C. Family Muscidae. In Catalogue of the Palearctic Diptera; Akadémiai Kiadó: Budapest, Hungary, 1986; Volume 11, pp. 57–215. [Google Scholar]
- Sorokina, V.S.; Ovtshinnikova, O.G. The position of the Azeliinae in the Muscidae (Diptera) based on musculature of the male terminalia. ZooKeys 2020, 975, 87–110. [Google Scholar] [CrossRef]
- Savage, J.; Wheeler, T.A. Phylogeny of the Azeliini (Diptera: Muscidae). Stud. Dipterol. 2004, 11, 259–299. [Google Scholar]
- Carvalho, C.J.B.D. Muscidae (Diptera) of the Neotropical Region: Taxonomy; Editora UFPR: Curitiba, Brazil, 2002; pp. 1–287. [Google Scholar]
- Baldacchino, F.; Muenworn, V.; Desquesnes, M.; Desoli, F.; Charoenviriyaphap, T.; Duvallet, G. Transmission of pathogens by Stomoxys flies (Diptera, Muscidae): A review. Parasite 2013, 20, 26. [Google Scholar] [CrossRef] [Green Version]
- Ma, T.; Huang, J. Characterization of the complete mitochondrial genome of Hydrotaea spinigera (Diptera: Muscidae) with phylogenetic implications. J. Asia-Pac. Entomol. 2018, 21, 1416–1423. [Google Scholar] [CrossRef]
- McAlpine, J.F.; Petersen, B.V.; Shewell, G.E.; Teskey, H.J.; Vockeroth, J.R.; Wood, D.M. Manual of Nearctic Diptera; Research Branch Agriculture Canada: Ontario, ON, Canada, 1993; Volume 2, pp. 675–1332. [Google Scholar]
- Ma, Z.Y.; Xue, W.Q.; Feng, Y. Fauna Sinica Insecta. Diptera Muscidae (II). Phaoniinae (I); Science Press: Beijing, China, 2002; Volume 26, p. 411. [Google Scholar]
- Karagozlu, M.Z.; Choi, T.-J.; Park, S.H.; Shin, S.E.; Kim, C.-B. Complete mitochondrial genome of a forensically important muscid, Hydrotaea chalcogaster (Diptera: Muscidae), with notes on its phylogenetic position. J. Asia-Pac. Entomol. 2018, 21, 1129–1136. [Google Scholar] [CrossRef]
- Dsouli, N.; Delsuc, F.; Michaux, J.; De Stordeur, E.; Couloux, A.; Veuille, M.; Duvallet, G. Phylogenetic analyses of mitochondrial and nuclear data in haematophagous flies support the paraphyly of the genus Stomoxys (Diptera: Muscidae). Infect. Genet. Evol. 2011, 11, 663–670. [Google Scholar] [CrossRef] [PubMed]
- Zachos, J.C.; Dickens, G.R.; Zeebe, R.E. An early Cenozoic perspective on greenhouse warming and carbon-cycle dynamics. Nature 2008, 451, 279–283. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bohaty, S.M.; Zachos, J.C. Significant Southern Ocean warming event in the late middle Eocene. Geology 2003, 31, 1017–1020. [Google Scholar] [CrossRef]
- Boscolo Galazzo, F.; Thomas, E.; Pagani, M.; Warren, C.; Luciani, V.; Giusberti, L. The middle Eocene climatic optimum (MECO): A multiproxy record of paleoceanographic changes in the southeast Atlantic (ODP Site 1263, Walvis Ridge): MECO repercussions in the SE Atlantic. Paleoceanography 2014, 29, 1143–1161. [Google Scholar] [CrossRef]
- Löwenberg-Neto, P.; Carvalho, C.J.B. Neotropical endemism and dispersal events between tropical and extra-tropical regions underlay the reticulate assemblages of muscid flies. J. Biogeogr. 2020, 47, 1574–1584. [Google Scholar] [CrossRef]





{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Subfamily | Tribe | Organism | Accession Number | Reference |
|---|---|---|---|---|
| Outgroup | Delia antiqua (Meigen, 1826) | NC_028226 | [28] | |
| Fannia scalaris (Fabricius, 1794) | NC_053661 | [29] | ||
| Megaselia scalaris (Loew, 1866) | KF974742 | [30] | ||
| Melophagus ovinus (Linnaeus, 1758) | MH024396 | [31] | ||
| Norellisoma spinimanum (Fallén, 1819) | NC_050316 | - | ||
| Atherigoninae | Atherigoniini | Atherigona ateripraepeda He, Huang et Fang, 2007 | OP528680 | This study |
| Atherigona miliaceae Malloch, 1925 | OP528691 | This study | ||
| Atherigona nigritibiella Fan et Liu, 1982 | OP528682 | This study | ||
| Azeliinae | Azeliini | Azelia zetterstedtii Rondani, 1866 | OP528689 | This study |
| Hydrotaea aenescens (Wiedemann, 1830) | NC_042952 | [23] | ||
| Hydrotaea chalcogaster (Wiedemann, 1824) | NC_041089 | [32] | ||
| Hydrotaea dentipes (Fabricius, 1805) | NC_047403 | [33] | ||
| Hydrotaea ignava (Harris, 1780) | NC_037195 | [34] | ||
| Hydrotaea sp. Robineau-Desvoidy, 1830 | KT272841 | [35] | ||
| Hydrotaea spinigera Hennig, 1962 | NC_042951 | [23] | ||
| Reinwardtiini | Muscina angustifrons (Loew, 1858) | NC_034805 | [36] | |
| Muscina levida (Harris, 1780) | NC_029487 | [35] | ||
| Muscina pascuorum (Meigen, 1826) | NC_053670 | [37] | ||
| Muscina stabulans (Fallén, 1817) | NC_026292 | [38] | ||
| Synthesiomyia nudiseta (van der Wulp, 1883) | NC_042953 | [23] | ||
| Coenosiinae | Coenosiini | Coenosia sponsa Xue et Tong, 2004 | OP528683 | This study |
| Pseudocoenosia fletcheri (Malloch, 1919) | OP528679 | This study | ||
| Pygophora sp. Schiner, 1868 | OP528684 | This study | ||
| Limnophorini | Limnophora nigripes (Robineau Desvoidy, 1830) | OP528687 | This study | |
| Limnophora surrecticerca Xue et Zhang, 1998 | OP528690 | This study | ||
| Spilogona spiniterebra (Stein, 1907) | OP528685 | This study | ||
| Lispini | Lispe sericipalpis Stein, 1904 | OP528692 | This study | |
| Muscinae | Muscini | Dasyphora quadrisetosa Zimin, 1951 | OP528694 | This study |
| Eudasyphora canadiana Cuny, 1980 | KT272852 | [35] | ||
| Morellia lopesae Pamplona, 1986 | KT272863 | [35] | ||
| Musca domestica Linnaeus, 1758 | NC_024855 | [39] | ||
| Musca sorbens Wiedemann, 1830 | NC_037910 | [40] | ||
| Stomoxyini | Haematobia irritans (Linnaeus, 1758) | NC_007102 | - | |
| Haematobosca stimulans (Meigen, 1824) | MT410787 | - | ||
| Stomoxys calcitrans (Linnaeus, 1758) | OP528693 | This study | ||
| Mydaeinae | Mydaeini | Graphomya rufitibia Stein, 1918 | NC_038210 | [41] |
| Hebecnema fumosa (Meigen, 1826) | OP528688 | This study | ||
| Mydaea bideserta Xue et Wang, 1992 | OP528681 | This study | ||
| Phaoniinae | Phaoniini | Helina impuncta (Fallén, 1823) | MT410825 | - |
| Helina latitarsis Ringdahl, 1924 | MT410783 | - | ||
| Phaonia errans (Meigen, 1826) | MT920423 | - | ||
| Phaonia pallida (Fabricius, 1787) | MT584137 | - | ||
| Phaonia tuguriorum (Scopoli, 1763) | MT410813 | - |
| Dataset Name | Protein-Coding Genes | Partition Scheme | tRNAs | rRNAs | |
|---|---|---|---|---|---|
| 1 | C123-P1 | all three codons | by gene | ||
| 2 | C123-P2 | all three codons | by locus | ||
| 3 | C123T-P1 | all three codons | by gene | 22 tRNAs | |
| 4 | C123T-P2 | all three codons | by locus | 22 tRNAs | |
| 5 | C123R-P1 | all three codons | by gene | 2 rRNAs | |
| 6 | C123R-P2 | all three codons | by locus | 2 rRNAs | |
| 7 | C123TR-P1 | all three codons | by gene | 22 tRNAs | 2 rRNAs |
| 8 | C123TR-P2 * | all three codons | by locus | 22 tRNAs | 2 rRNAs |
| 9 | C12-P2 | remove 3rd codon | by locus | ||
| 10 | C12T-P2 | remove 3rd codon | by locus | 22 tRNAs | |
| 11 | C12R-P2 | remove 3rd codon | by locus | 2 rRNAs | |
| 12 | C12TR-P2 | remove 3rd locus | by locus | 22 tRNAs | 2 rRNAs |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Li, X.; Cai, X.; Ding, S.; Wang, L.; Li, W.; Liu, X.; Zhang, C.; Yang, D. Phylogeny and Evolutionary Timescale of Muscidae (Diptera: Calyptratae) Inferred from Mitochondrial Genomes. Insects 2023, 14, 286. https://doi.org/10.3390/insects14030286
Li X, Cai X, Ding S, Wang L, Li W, Liu X, Zhang C, Yang D. Phylogeny and Evolutionary Timescale of Muscidae (Diptera: Calyptratae) Inferred from Mitochondrial Genomes. Insects. 2023; 14(3):286. https://doi.org/10.3390/insects14030286
Chicago/Turabian StyleLi, Xin, Xiaodong Cai, Shuangmei Ding, Liang Wang, Wenliang Li, Xiaoyan Liu, Chuntian Zhang, and Ding Yang. 2023. "Phylogeny and Evolutionary Timescale of Muscidae (Diptera: Calyptratae) Inferred from Mitochondrial Genomes" Insects 14, no. 3: 286. https://doi.org/10.3390/insects14030286