
Petals Reduce Attachment of Insect Pollinators: A Case Study of the Plant Dahlia pinnata and the Fly Eristalis tenax


{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:Simple Summary
Abstract
1. Introduction
2. Materials and Methods
2.1. Plants and Insects
2.2. Microscopy
2.3. Experiments
3. Results
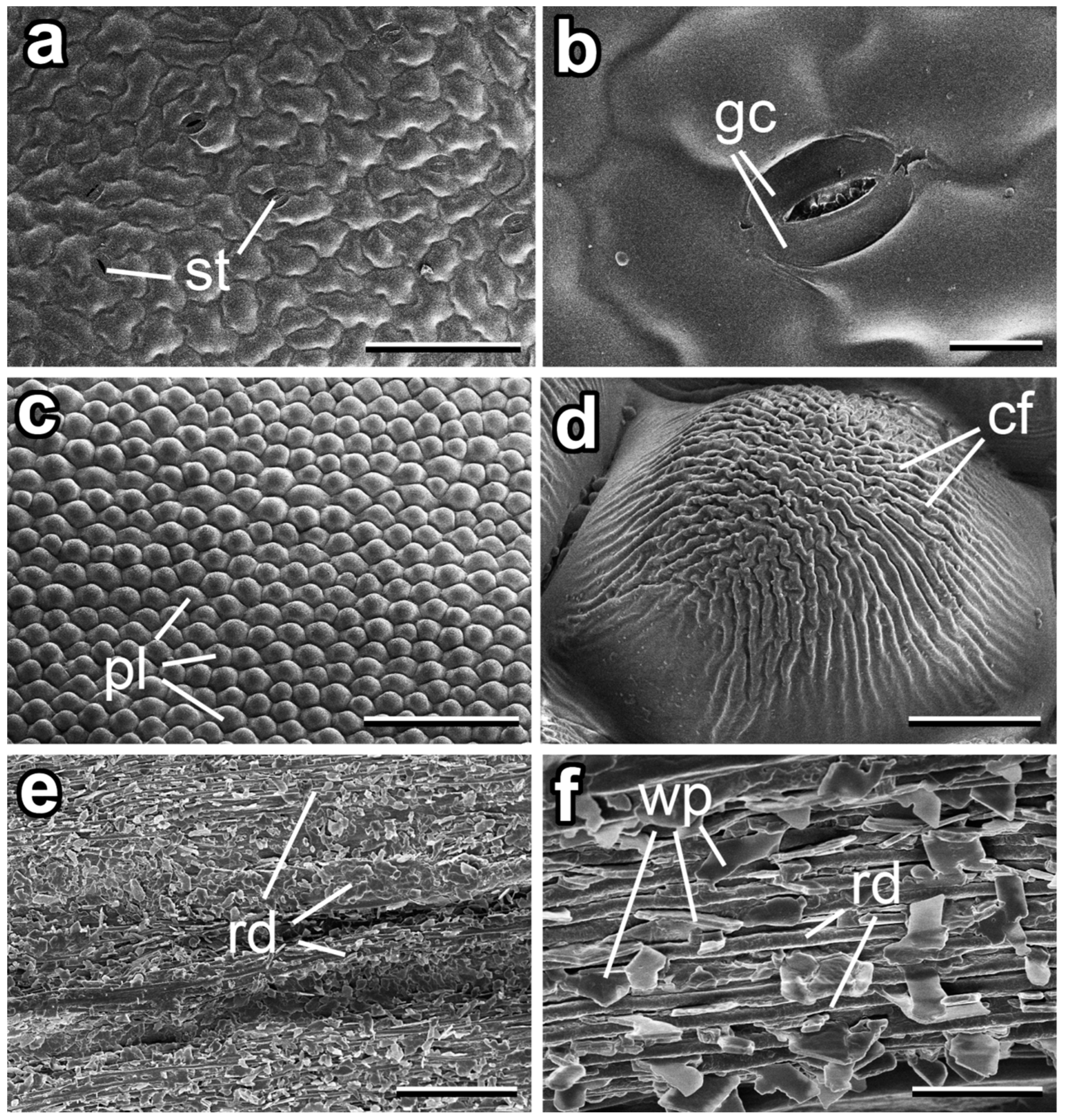
3.1. Micromorphology of Dahlia pinnata Surfaces
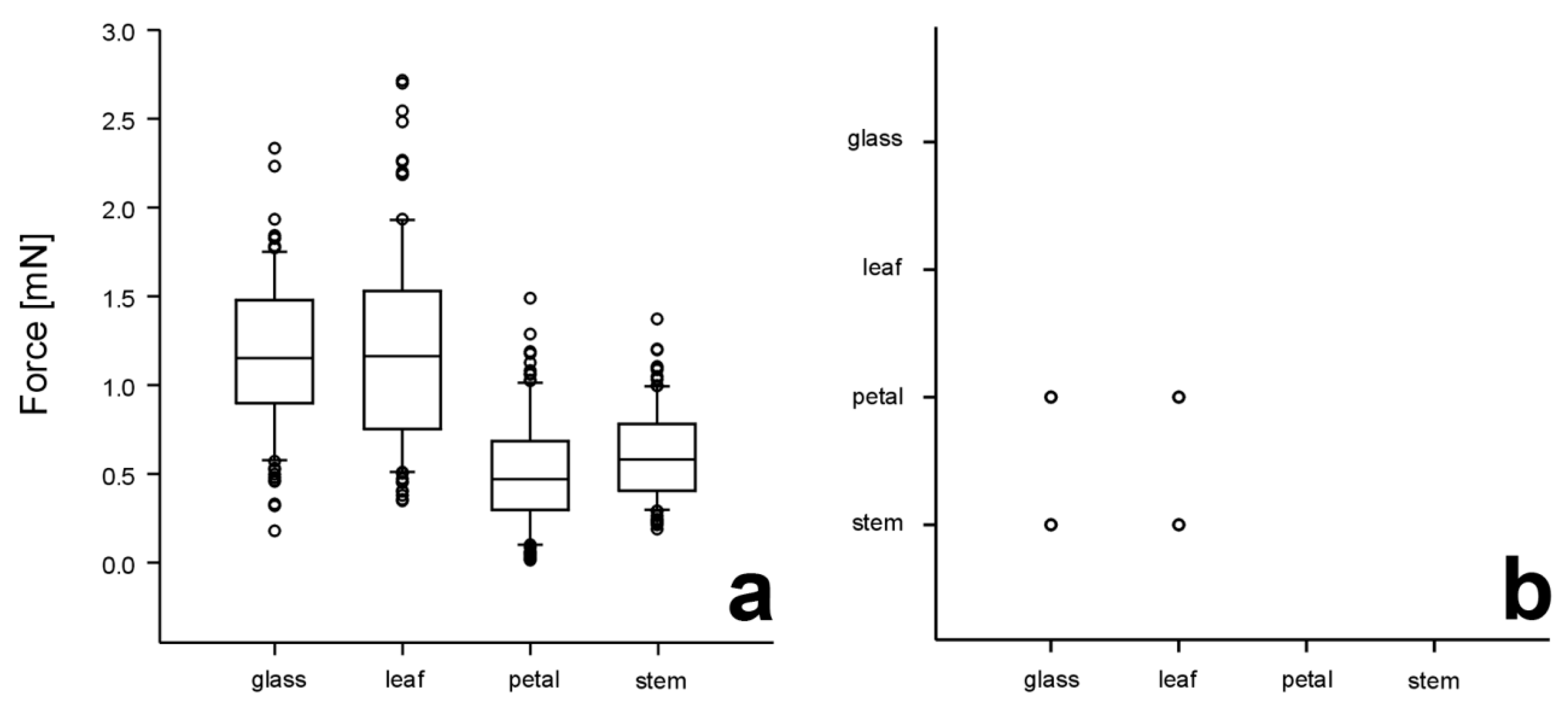
3.2. Traction Forces on Different Test Substrates
4. Discussion
Author Contributions
Funding
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Southwood, S. Plant Surfaces and Insects—An Overview. In Insects and the Plant Surface; Juniper, B., Southwood, R., Eds.; Edward Arnold Publishers: London, UK, 1986; pp. 1–22. [Google Scholar]
- Jeffree, C.F. The Cuticle, Epicuticular Waxes and Trichomes of Plants, with Reference to Their Structure, Function and Evolution. In Insects and the Plant Surface; Juniper, B., Southwood, R., Eds.; Edward Arnold Publishers: London, UK, 1986; pp. 23–63. [Google Scholar]
- Gorb, E.V.; Gorb, S.N. Anti-Adhesive Surfaces in Plants and Their Biomimetic Potential. In Materials Design Inspired by Nature: Function Through Inner Architecture; Fratzl, P., Dunlop, J.W.C., Weinkamer, R., Eds.; RSC Publishing: Cambridge, UK, 2013; pp. 282–309. [Google Scholar]
- Levin, D.A. The role of trichomes in plant defense. Q. Rev. Biol. 1973, 48, 3–15. [Google Scholar] [CrossRef]
- Johnson, B. The injurious effects of the hooked epidermal hairs of French beans (Phaseolus vulgaris L.) on Aphis craccivora Koch. Bull. Entomol. Res. 1953, 44, 779–788. [Google Scholar] [CrossRef]
- Rebora, M.; Salerno, G.; Piersanti, S.; Gorb, E.; Gorb, S.N. Entrapment of Bradysia paupera (Diptera: Sciaridae) by Phaseolus vulgaris (Fabaceae) plant leaf. Arthropod. Plant. Interact. 2020, 14, 499–509. [Google Scholar] [CrossRef]
- Hare, J.; Elle, E. Variable impact of diverse insect herbivores on dimorphic Datura wrightii. Ecology 2002, 83, 2711–2720. [Google Scholar] [CrossRef]
- Andres, M.R.; Connor, E.F. The community-wide and guild-specific effects of pubescence on the folivorous insects of manzanitas Arctostaphylos spp. Ecol. Entomol. 2003, 28, 383–396. [Google Scholar] [CrossRef] [Green Version]
- Gibson, R.W. Glandular hairs providing resistance to aphids in certain wild potato species. Ann. Appl. Bio. 1971, 68, 113–119. [Google Scholar] [CrossRef]
- Dufey, S.S. Plant Glandular Trichomes: Their Partial Role in Defense Against Insects. In Insects and the Plant Surface; Juniper, B., Southwood, R., Eds.; Edward Arnold Publishers: London, UK, 1986; pp. 151–172. [Google Scholar]
- Voigt, D.; Gorb, E.; Gorb, S. Plant surface–bug interactions: Dicyphus errans stalking along trichomes. Arthropod Plant Interact. 2007, 1, 221–243. [Google Scholar] [CrossRef]
- Prüm, B.; Seidel, R.; Bohn, H.F.; Speck, T. Plant surfaces with cuticular folds are slippery for beetles. J. R. Soc. Interface 2011, 9, 127–135. [Google Scholar] [CrossRef] [Green Version]
- Prüm, B.; Bohn, H.F.; Seidel, R.; Rubach, S.; Speck, T. Plant surfaces with cuticular folds and their replicas: Influence of microstructuring and surface chemistry on the attachment of a leaf beetle. Acta Biomater. 2013, 9, 6360–6368. [Google Scholar] [CrossRef]
- Surapaneni, V.A.; Aust, T.; Speck, T.; Thielen, M. Polarity in cuticular ridge development and insect attachment on leaf surfaces of Schismatoglottis calyptrata (Araceae). Beilstein J. Nanotechnol. 2021, 12, 1326–1338. [Google Scholar] [CrossRef]
- Gorb, E.V.; Gorb, S.N. Anti-adhesive effects of plant wax coverage on insect attachment. J. Exp. Bot. 2017, 68, 5323–5337. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Gorb, E.V.; Gorb, S.N. Attachment ability of the beetle Chrysolina fastuosa on various plant surfaces. Entomol. Exp. Appl. 2002, 105, 13–28. [Google Scholar] [CrossRef]
- Gorb, E.V.; Purtov, J.; Gorb, S.N. Adhesion force measurements on the two wax layers of the waxy zone in Nepenthes alata pitchers. Sci. Rep. 2014, 4, 5154. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Juniper, B.E.; Robins, R.J.; Joel, D.M. The Carnivorous Plants; Academic Press: London, UK, 1989; 353p. [Google Scholar]
- Gaume, L.; Gorb, S.; Rowe, N. Function of epidermal surfaces in the trapping efficiency of Nepenthes alata pitchers. New Phytol. 2002, 156, 479–489. [Google Scholar] [CrossRef]
- Gorb, E.; Haas, K.; Henrich, A.; Enders, S.; Barbakadze, N.; Gorb, S. Composite structure of the crystalline epicuticular wax layer of the slippery zone in the pitchers of the carnivorous plant Nepenthes alata and its effect on insect attachment. J. Exp. Biol. 2005, 208, 4651–4662. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Scholz, I.; Bückins, M.; Dolge, L.; Erlinghagen, T.; Weth, A.; Hischen, F.; Mayer, J.; Hoffmann, S.; Riederer, M.; Riedel, M.; et al. Slippery surfaces of pitcher plants: Nepenthes wax crystals minimize insect attachment via microscopic surface roughness. J. Exp. Biol. 2010, 213, 1115–1125. [Google Scholar] [CrossRef] [Green Version]
- Daumer, K. Blumenfarben, wie sie die Bienen sehen. Z. Vergl. Physiol. 1958, 41, 49–110. [Google Scholar] [CrossRef]
- Kugler, H. Blütenokologie; Gustav Fischer Verlag: Stuttgart, Germany, 1970; 345p. [Google Scholar]
- Willmer, P. Pollination and Floral Ecology; Princeton University Press: Princeton, NJ, USA, 2011; 832p. [Google Scholar]
- Oelschlägel, B.; Gorb, S.; Wanke, S.; Neinhuis, C. Structure and biomechanics of trapping flower trichomes and their role in the pollination biology of Aristolochia plants (Aristolochiaceae). New Phytol. 2009, 184, 988–1002. [Google Scholar] [CrossRef]
- Müller, L. Zur biologischen Anatomie der Blüte von Ceropegia woodii Schlechter. Biol. Gen. 1926, 2, 799–814. [Google Scholar]
- Vogel, S. Die Bestäubung der Kesselfallen-Blüten von Ceropegia. Beiträge Zur Biol. Der Pflanz. 1961, 36, 159–237. [Google Scholar]
- Vogel, S.; Martens, J. A survey of the lethal kettle traps of Arisaema (Araceae), with records of pollinating fungus gnats from Nepal. Bot. J. Linnean Soc. 2000, 133, 61–100. [Google Scholar] [CrossRef]
- Poppinga, S.; Koch, K.; Bohn, H.F.; Barthlott, W. Comparative and functional morphology of hierarchically structured anti-adhesive surfaces in carnivorous plants and kettle trap flowers. Funct. Plant Biol. 2010, 37, 952–961. [Google Scholar] [CrossRef]
- Barthlott, W.; Ehler, N. Raster-Elektronenmikroskopie der Epidermis-Oberflächen von Spermatophyten (Tropische und Subtropische Pflanzenwelt); Franz Steiner Verlag: Stuttgart, Germany, 1977; 105p. [Google Scholar]
- Christensen, K.; Hansen, H. SEM-studies of epidermal patterns of petals in the angiosperms. Opera Bot. 1998, 135, 1–91. [Google Scholar]
- Whitney, H.M.; Chittka, L.; Glover, B.J. Conical epidermal cells allow bees to grip flowers and increase foraging efficiency. Curr. Biol. 2009, 19, 948–953. [Google Scholar] [CrossRef] [Green Version]
- Whitney, H.M.; Bennett, K.M.; Dorling, M.; Sandbach, L.; Prince, D.; Chittka, L.; Glover, B.J. Why do so many petals have conical epidermal cells. Ann. Bot. 2011, 108, 609–616. [Google Scholar] [CrossRef]
- Prüm, B.; Seidel, R.; Bohn, H.F.; Speck, T. Impact of cell shape in hierarchically structured plant surfaces on the attachment of male Colorado potato beetles (Leptinotarsa decemlineata). Beilstein J. Nanotechnol. 2012, 3, 57–64. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Papiorek, S.; Junker, R.R.; Lunau, K. Gloss, colour and grip: Multifunctional epidermal cell shapes in bee- and bird-pollinated flowers. PLoS ONE 2014, 9, e112013. [Google Scholar] [CrossRef] [Green Version]
- Knoll, F. Insekten und Blumen: Experimentelle Arbeiten zur Vertiefung unserer Kenntnisse über die Wechselbeziehungen zwischen Pflanzen und Tieren. Abbandl. D Zool Botan. Gesellsch. Wien 1921, 12, 1–646. [Google Scholar] [CrossRef] [Green Version]
- Bräuer, P.; Neinhuis, C.; Voigt, D. Attachment of honeybees and greenbottle flies to petal surfaces. Arthropod Plant Interact 2017, 11, 171–192. [Google Scholar] [CrossRef]
- Jäger, E.J.; Ebel, F.; Hanelt, P.; Müller, G.K. Rothmaler Exkursionsflora von Deutschland. 5: Krautige Zier- und Nutzpflanzen; Spektrum: Heidelberg, Germany, 2008; 879p. [Google Scholar]
- Heal, J.R. Colour patterns of syrphidae: IV. Mimicry and variation in natural populations of Eristalis tenax. Heredity 1982, 49, 95–109. [Google Scholar] [CrossRef]
- Buckton, G.B. The Natural History of Eristalis Tenax or the Drone-Fly; Macmillan: London, UK; New York, NY, USA, 1895; 140p. [Google Scholar]
- Holloway, B.A. Pollen-feeding in hover-flies (Diptera: Syrphidae). N. Z. J. Zool. 1979, 3, 339–350. [Google Scholar] [CrossRef]
- Skevington, J.H.; Locke, M.M.; Young, A.D.; Moran, K.; Crins, W.J.; Marshall, S.A. Field Guide to the Flower Flies of Northeastern North America; Princeton University Press: Princeton, NJ, USA, 2019; 512p. [Google Scholar]
- Gorb, S.; Gorb, E.; Kastner, V. Scale effects on the attachment pads and friction forces in syrphid flies (Diptera, Syrphydae). J. Exp. Biol. 2001, 204, 1421–1431. [Google Scholar] [CrossRef]
- Barthlott, W.; Neinhuis, C.; Cutler, D.; Ditsch, F.; Meusel, I.; Theisen, I.; Wilhelmi, H. Classification and terminology of plant epicuticular waxes. Bot. J. Linn. Soc. 1998, 126, 237–260. [Google Scholar] [CrossRef]
- Gorb, S.N.; Popov, V.L. Probabilistic fasteners with parabolic elements: Biological system artificial model and theoretical considerations. Phil. Trans. R. Soc. Lond. A 2002, 360, 211–225. [Google Scholar] [CrossRef]
- Gorb, E.; Hosoda, N.; Miksch, C.; Gorb, S. Slippery pores: Anti-adhesive effect of nanoporous substrates on the beetle attachment system. J. R. Soc. Interface 2010, 7, 1571–1579. [Google Scholar] [CrossRef] [Green Version]
- Gorb, E.; Gorb, S. How a lack of choice can force ants to climb up waxy plant stems. Arthropod Plant Interact 2011, 5, 297–306. [Google Scholar] [CrossRef]
- Stork, N.E. Experimental analysis of adhesion of Chrysolina polita (Chrysomelidae, Coleoptera) on a variety of surfaces. J. Exp. Biol. 1980, 88, 91–107. [Google Scholar] [CrossRef]
- Edwards, P.B. Do waxes of juvenile Eucalyptus leaves provide protection from grazing insects. Aust. J. Ecol. 1982, 7, 347–352. [Google Scholar] [CrossRef]
- Federle, W.; Maschwitz, U.; Fiala, B.; Riederer, M.; Hölldobler, B. Slippery ant-plants and skilful climbers: Selection and protection of specific ant partners by epicuticular wax blooms in Macaranga (Euphorbiaceae). Oecologia 1997, 112, 217–224. [Google Scholar] [CrossRef]
- Eigenbrode, S.D.; White, C.; Rohde, M.; Simon, C.J. Behavior and effectiveness of adult Hyppodamia convergens (Coleoptera: Coccinellidae) as a predator of Acyrthosiphon pisum on a glossy-wax mutant of Pisum sativum. Environ. Entomol. 1998, 91, 902–909. [Google Scholar] [CrossRef] [Green Version]
- Gorb, E.; Gorb, S. Effects of surface topography and chemistry of Rumex obtusifolius leaves on the attachment of the beetle Gastrophysa viridula. Entomol. Exp. Appl. 2009, 130, 222–228. [Google Scholar] [CrossRef]
- Gorb, E.V.; Gorb, S.N. 2006. Do Plant Waxes Make Insect Attachment Structures Dirty? Experimental Evidence for the Contamination Hypothesis. In Ecology and Biomechanics: A Mechanical Approach to the Ecology of Animals and Plants; Herrel, A., Speck, T., Rowe, N.P., Eds.; CRC Press: Boca Raton, FL, USA, 2006; pp. 147–162. [Google Scholar]
- Gorb, E.V.; Lemke, W.; Gorb, S.N. Porous substrate affects a subsequent attachment ability of the beetle Harmonia axyridis (Coleoptera, Coccinellidae). J. R. Soc. Interface 2019, 16, 20180696. [Google Scholar] [CrossRef] [Green Version]
- Kerner von Marilaun, A. Flowers and Their Unbidden Guests; C. K. Paul & Co.: London, UK, 1878. [Google Scholar]
- Harley, R.M. Evolution and Distribution of Eriope (Labiatae) and its Relatives in Brazil. In Proceedings of a Workshop on Neotropical Distributions; Academia Brasileira de Ciencias: Rio de Janeiro, Brazil, 1988; pp. 71–120. [Google Scholar]
- Harley, R. The Greasy Pole Syndrome. In Ant-Plant Interactions; Huxley, C.R., Cutler, D.E., Eds.; Oxford University Press: Oxford, UK, 1991; pp. 430–433. [Google Scholar]
- Gorb, S.N.; Gorb, E.V. Frequency of plant visits by the generalist ant Lasius niger depends on the surface microstructure of plant stems. Arthropod Plant Interact 2019, 13, 311–320. [Google Scholar] [CrossRef]
- Gorb, S.N.; Gorb, E.V. Combined effect of different flower stem features on the visiting frequency of the generalist ant Lasius niger: An experimental study. Insects 2021, 12, 1026. [Google Scholar] [CrossRef]
- Feng, L.; Zhang, Y.; Xi, J.; Zhu, Y.; Wang, N.; Xia, F.; Jiang, L. Petal effect: A superhydrophobic state with high adhesive force. Langmuir 2008, 24, 4114–4119. [Google Scholar] [CrossRef]
- Bhushan, B.; Nosonovsky, M. The rose petal effect and the modes of superhydrophobicity. Philos. Trans. Royal. Soc. A 2010, 368, 4713–4728. [Google Scholar] [CrossRef]
- Jin, M.H.; Feng, X.L.; Feng, L.; Sun, T.L.; Zhai, J.; Li, T.J.; Jiang, L. Superhydrophobic aligned polystyrene nanotube films with high adhesive force. Adv. Mater. 2005, 17, 1977–1981. [Google Scholar] [CrossRef]
- Bormashenko, E.; Stein, T.; Pogreb, R.; Aurbach, D. “Petal Effect” on surfaces based on lycopodium: High-stick surfaces demonstrating high apparent contact angles. J. Phys. Chem. C 2009, 113, 5568–5572. [Google Scholar] [CrossRef]
- Park, Y.M.; Gang, M.; Seo, Y.H.; Kim, B.H. Artificial petal surface based on hierarchical micro- and nanostructures. Thin Solid Films 2011, 520, 362–367. [Google Scholar] [CrossRef]
- Féat, A.; Federle, W.; Kamperman, M.; van der Gucht, J. Coatings preventing insect adhesion: An overview. Progr. Org. Coat. 2019, 134, 349–359. [Google Scholar] [CrossRef]
- Zhou, Y.; Robinson, A.; Steiner, U.; Federle, W. 2014 Insect adhesion on rough surfaces: Analysis of adhesive contact of smooth and hairy pads on transparent microstructured substrates. J. R. Soc. Interface 2014, 11, 20140499. [Google Scholar] [CrossRef] [Green Version]
- Gorb, E.; Böhm, S.; Jacky, N.; Maier, L.P.; Dening, K.; Pechook, S.; Pokroy, B.; Gorb, S. Insect attachment on crystalline bioinspired wax surfaces formed by alkanes of varying chain lengths. Beilstein J. Nanotechnol. 2014, 5, 1031–1041. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Eichler-Volf, A.; Xue, L.; Dornberg, G.; Chen, H.; Kovalev, A.; Enke, D.; Wang, Y.; Gorb, E.V.; Gorb, S.N.; Steinhart, M. The influence of surface topography and surface chemistry on the anti-adhesive performance of nanoporous monoliths. ACS Appl. Mater. Interfaces 2016, 8, 22593–22604. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Eichler-Volf, A.; Kovalev, A.; Wedeking, T.; Gorb, E.V.; Xue, L.; You, C.; Piehler, J.; Gorb, S.N.; Steinhart, M. Bioinspired monolithic polymer microsphere arrays as generically anti-adhesive surfaces. Bioinspir. Biomim. 2016, 11, 025002. [Google Scholar] [CrossRef] [PubMed] [Green Version]




Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Gorb, E.V.; Gorb, S.N. Petals Reduce Attachment of Insect Pollinators: A Case Study of the Plant Dahlia pinnata and the Fly Eristalis tenax. Insects 2023, 14, 285. https://doi.org/10.3390/insects14030285
Gorb EV, Gorb SN. Petals Reduce Attachment of Insect Pollinators: A Case Study of the Plant Dahlia pinnata and the Fly Eristalis tenax. Insects. 2023; 14(3):285. https://doi.org/10.3390/insects14030285
Chicago/Turabian StyleGorb, Elena V., and Stanislav N. Gorb. 2023. "Petals Reduce Attachment of Insect Pollinators: A Case Study of the Plant Dahlia pinnata and the Fly Eristalis tenax" Insects 14, no. 3: 285. https://doi.org/10.3390/insects14030285