
Drivers of Insect Community Change along the Margins of Mountain Streams in Serra da Estrela Natural Park (Portugal)
 , ,
, ,
Abstract
:Simple Summary
Abstract
1. Introduction
2. Materials and Methods
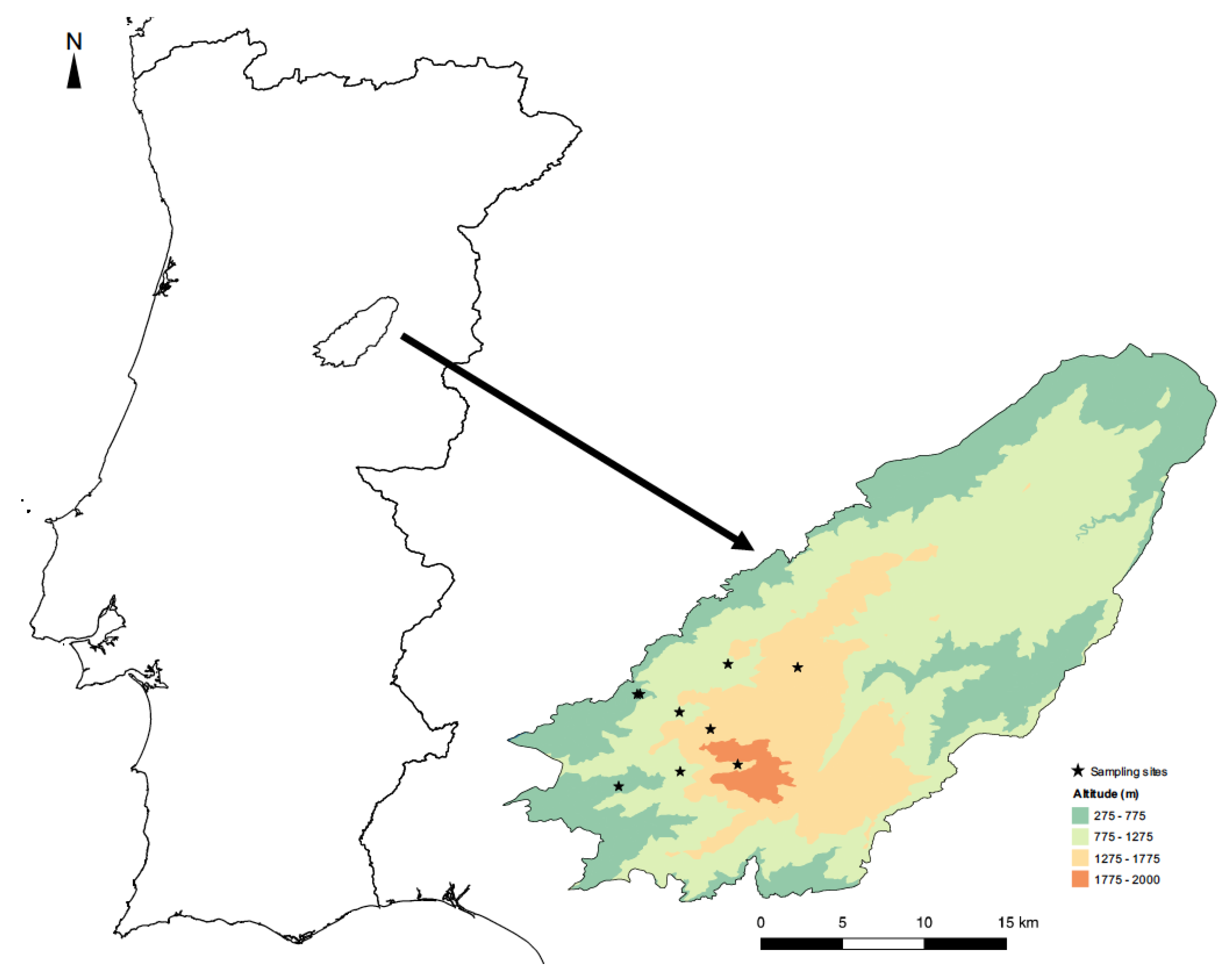
2.1. Study Area
2.2. Data Collection: Butterfly and Odonate Sampling
2.3. Data Analysis
3. Results
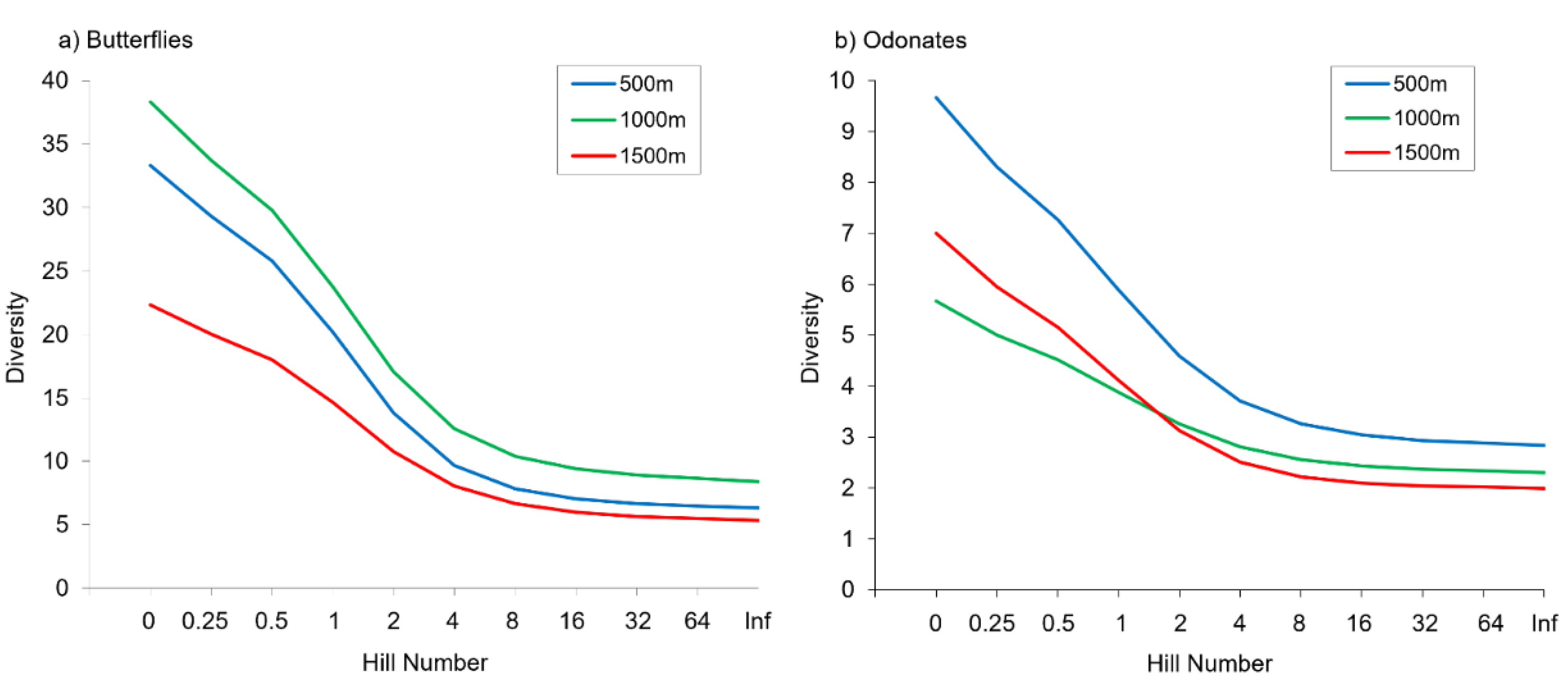
3.1. Alpha Diversity Patterns
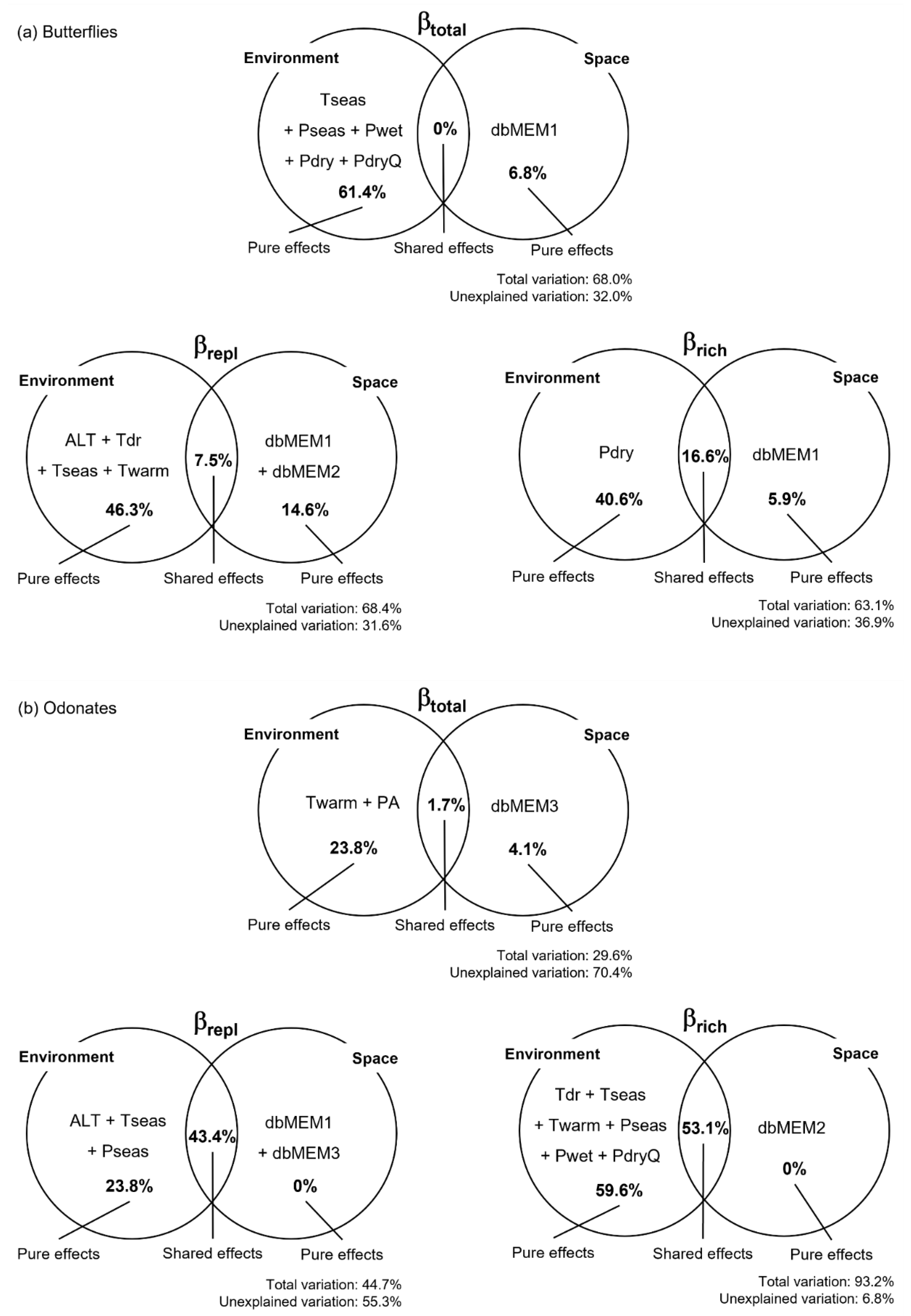
3.2. Beta Diversity Patterns and Drivers
4. Discussion
4.1. Alpha Diversity Patterns
4.2. Beta Diversity Patterns
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Rahbek, C. The role of spatial scale and the perception of large-scale species-richness patterns. Ecol. Lett. 2005, 8, 224–239. [Google Scholar] [CrossRef]
- Tittensor, D.P.; Walpole, M.; Hill, S.L.L.; Boyce, D.G.; Britten, G.L.; Burgess, N.D.; Butchart, S.H.M.; Leadley, P.W.; Regan, E.C.; Alkemade, R.; et al. A mid-term analysis of progress toward international biodiversity targets. Science 2014, 346, 241–244. [Google Scholar] [CrossRef] [PubMed]
- Diaz, H.F.; Grosjean, M.; Graumlich, L. Climate variability and change in high elevation regions: Past, present and future. Clim. Change 2003, 59, 1–4. [Google Scholar] [CrossRef]
- Hodkinson, I.D.; Jackson, J.K. Terrestrial and aquatic invertebrates as bioindicators for environmental monitoring, with particular reference to mountain ecosystems. Environ. Manage. 2005, 35, 649–666. [Google Scholar] [CrossRef]
- Aguiar, C.; Rodrigues, O.; Azevedo, J.; Domingos, T. Montanha. In Ecossistemas e Bem-Estar Humano: Avaliação para Portugal do Millennium Ecosystem Assessment; Pereira, H.M., Domingos, T., Vicente, L., Proença, V., Eds.; Escolar Editora: Lisboa, Portugal, 2009; pp. 295–339. [Google Scholar]
- Nogués-Bravo, D.; Araújo, M.B.; Romdal, T.; Rahbek, C. Scale effects and human impact on the elevational species richness gradients. Nature 2008, 453, 216–219. [Google Scholar] [CrossRef] [Green Version]
- McCoy, E.D. The distribution of insects along elevational gradients. Oikos 1990, 58, 313–322. [Google Scholar] [CrossRef]
- Sanders, N.J. Elevational gradients in ant species richness: Area, geometry, and Rapoport’s rule. Ecography 2002, 25, 25–32. [Google Scholar] [CrossRef]
- Choi, S.; An, J. Altitudinal distribution of moths (Lepidoptera) in Mt. Jirisan National Park, South Korea. Eur. J. Entomol. 2010, 107, 229–245. [Google Scholar] [CrossRef] [Green Version]
- Werenkraut, V.; Ruggiero, A. The richness and abundance of epigaeic mountain beetles in north-western Patagonia, Argentina: Assessment of patterns and environmental correlates. J. Biogeogr. 2014, 41, 561–573. [Google Scholar] [CrossRef]
- Wang, J.; Soininen, J.; Zhang, Y.; Wang, B.; Yang, X.; Shen, J. Patterns of elevational beta diversity in micro-and macroorganisms. Global Ecol. Biogeogr. 2012, 21, 743–750. [Google Scholar] [CrossRef]
- Bishop, T.R.; Robertson, M.P.; van Rensburg, B.J.; Parr, C.L. Contrasting species and functional beta diversity in montane ant assemblages. J. Biogeogr. 2015, 42, 1776–1786. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zellweger, F.; Roth, T.; Bugmann, H.; Bollmann, K. Beta diversity of plants, birds and butterflies is closely associated with climate and habitat structure. Global Ecol. Biogeogr. 2017, 26, 898–906. [Google Scholar] [CrossRef]
- Perillo, L.N.; Neves, F.S.; Antonini, Y.; Martins, R.P. Compositional changes in bee and wasp communities along Neotropicalmountain altitudinal gradient. PLoS ONE 2017, 12, e0182054. [Google Scholar] [CrossRef] [Green Version]
- Beirão, M.V.; Neves, F.S.; Fernandes, G.W. Climate and plant structure determine the spatiotemporal butterfly distribution on a tropical mountain. Biotropica 2021, 53, 191–200. [Google Scholar] [CrossRef]
- Fontana, V.; Guariento, E.; Hilpold, A.; Niedrist, G.; Steinwandter, M.; Spitale, D.; Nascimbene, J.; Tappeiner, U. Species richness and beta diversity patterns of multiple taxa along an elevational gradient in pastured grasslands in the European Alps. Sci. Rep. 2020, 10, 12516. [Google Scholar] [CrossRef] [PubMed]
- Nunes, C.A.; Braga, R.F.; Figueira, J.E.C.; Neves, F.D.S.; Fernandes, G.W. Dung beetles along a tropical altitudinal gradient: Environmental filtering on taxonomic and functional diversity. PLoS ONE 2016, 11, e0157442. [Google Scholar] [CrossRef] [Green Version]
- Nunes, C.A.; Quintino, A.V.; Constantino, R.; Negreiros, D.; Reis Junior, R.; Fernandes, G.W. Patterns of taxonomic and functional diversity of termites along a tropical elevational gradient. Biotropica 2017, 49, 186–194. [Google Scholar] [CrossRef]
- Castro, F.S.D.; Da Silva, P.G.; Solar, R.; Fernandes, G.W.; Neves, F.D.S. Environmental drivers of taxonomic and functional diversity of ant communities in a tropical mountain. Insect Conserv. Divers. 2020, 13, 393–403. [Google Scholar] [CrossRef]
- Schowalter, T.D. Insect Ecology: An Ecosystem Approach, 4th ed.; Academic Press: London, UK, 2016. [Google Scholar]
- Noriega, J.A.; Hortal, J.; Azcárate, F.M.; Berg, M.P.; Bonada, N.; Briones, M.J.I.; Del Toro, I.; Goulson, D.; Ibanez, S.; Landis, D.A.; et al. Research trends in ecosystem services provided by insects. Basic Appl. Ecol. 2018, 26, 8–23. [Google Scholar] [CrossRef] [Green Version]
- Hallmann, C.A.; Sorg, M.; Jongejans, E.; Siepel, H.; Hofland, N.; Schwan, H.; Hörren, T.; Goulson, D.; de Kroon, H. More than 75 percent decline over 27 years in total flying insect biomass in protected areas. PLoS ONE 2017, 12, e0185809. [Google Scholar] [CrossRef] [Green Version]
- Simmons, B.I.; Balmford, A.; Bladon, A.J.; Christie, A.P.; De Palma, A.; Dicks, L.V.; Gallego-Zamorano, J.; Johnston, A.; Martin, P.A.; Purvis, A.; et al. Worldwide insect declines: An important message, but interpret with caution. Ecol. Evol. 2019, 9, 3678–3680. [Google Scholar] [CrossRef] [Green Version]
- Harvey, J.A.; Heinen, R.; Klein, A.-M.; Armbrecht, I.; Basset, Y.; Baxter-Gilbert, J.H.; Bezemer, M.; Böhm, M.; Bommarco, R.; Borges, P.A.V.; et al. International scientists formulate a roadmap for insect conservation and recovery. Nat. Ecol. Evol. 2020, 4, 174–176. [Google Scholar] [CrossRef]
- Wagner, D.L. Insect declines in the Anthropocene. Annu. Rev. Entomol. 2020, 65, 457–480. [Google Scholar] [CrossRef] [Green Version]
- Miličić, M.; Popov, S.; Branco, V.V.; Cardoso, P. Insect threats and conservation through the lens of global experts. Conserv. Lett. 2021, e12814. [Google Scholar] [CrossRef]
- Múrria, C.; Sáinz-Bariáin, M.; Vogler, A.P.; Viza, A.; González, M.; Zamora-Muñoz, C. Vulnerability to climate change for two endemic high-elevation, low-dispersive Annitella species (Trichoptera) in Sierra Nevada, the southernmost high mountain in Europe. Insect Conserv. Divers. 2020, 13, 283–295. [Google Scholar] [CrossRef]
- Fourcade, Y.; Åström, S.; Öckinger, E. Decline of parasitic and habitat-specialist species drives taxonomic, phylogenetic and functional homogenization of sub-alpine bumblebee communities. Oecologia 2021, 196, 905–917. [Google Scholar] [CrossRef]
- Wilson, R.J.; Gutiérrez, D.; Gutiérrez, J.; Monserrat, V.J. An elevational shift in butterfly species richness and composition accompanying recent climate change. Global Change Biol. 2007, 13, 1873–1887. [Google Scholar] [CrossRef]
- Scalercio, S.; Bonacci, T.; Mazzei, A.; Pizzolotto, R.; Brandmayr, P. Better up, worse down: Bidirectional consequences of three decades of climate change on a relict population of Erebia cassioides. J. Insect Conserv. 2014, 18, 643–650. [Google Scholar] [CrossRef]
- Zografou, K.; Kati, V.; Grill, A.; Wilson, R.J.; Tzirkalli, E.; Pamperis, L.N.; Halley, J.M. Signals of climate change in butterfly communities in a Mediterranean protected area. PLoS ONE 2014, 9, e87245. [Google Scholar] [CrossRef] [Green Version]
- Simaika, J.P.; Samways, M.J. Predicted range shifts of dragonflies over a wide elevation gradient in the southern hemisphere. Freshw. Sci. 2015, 34, 1133–1143. [Google Scholar] [CrossRef] [Green Version]
- Talavera, G.; Espadaler, X.; Vila, R. Discovered just before extinction? The first endemic ant from the Balearic Islands (Lasius balearicus sp. nov.) is endangered by climate change. J. Biogeogr. 2015, 42, 589–601. [Google Scholar] [CrossRef]
- Oertli, B. The use of dragonflies in the assessment and monitoring of aquatic habitats. In Dragonflies and Damselflies: Model Organisms for Ecological and Evolutionary Research; Córdoba-Aguilar, A., Ed.; Oxford University Press: New York, NY, USA, 2008; pp. 79–96. [Google Scholar]
- Gerlach, J.; Samways, M.; Pryke, J. Terrestrial invertebrates as bioindicators: An overview of available taxonomic groups. J. Insect Conserv. 2013, 17, 831–850. [Google Scholar] [CrossRef]
- Kitahara, M.; Yumoto, M.; Kobayashi, T. Relationship of butterfly diversity with nectar plant species richness in and around the Aokigahara primary woodland of Mount Fuji, central Japan. Biodiv. Conserv. 2008, 17, 2713–2734. [Google Scholar] [CrossRef]
- Thomas, J.A. Monitoring change in the abundance and distribution of insects using butterflies and other indicator groups. Phil. Trans. R. Soc. B 2005, 360, 339–357. [Google Scholar] [CrossRef] [PubMed]
- Ott, J. (Ed.) Monitoring Climatic Change with Dragonflies; Biorisk 5; Pensoft: Sophia, Bulgaria, 2010. [Google Scholar]
- Goertzen, D.; Suhling, F. Urbanization versus other land use: Diverging effects on dragonfly communities in Germany. Divers. Distrib. 2019, 25, 38–47. [Google Scholar] [CrossRef] [Green Version]
- Rocha-Ortega, M.; Rodríguez, P.; Córdoba-Aguilar, A. Can dragonfly and damselfly communities be used as bioindicators of land use intensification? Ecol. Indic. 2019, 107, 105553. [Google Scholar] [CrossRef]
- Cleary, D.F.R.; Mooers, A.Ø.; Eichhorn, K.A.O.; van Tol, J.; de Jong, R.; Menken, S.B.J. Diversity and community composition of butterflies and odonates in an ENSO-induced fire affected habitat mosaic: A case study from East Kalimantan, Indonesia. Oikos 2004, 105, 426–446. [Google Scholar] [CrossRef]
- McPeek, M.A. Ecological factors limiting the distributions and abundances of Odonata. In Dragonflies and Damselflies: Model Organisms for Ecological and Evolutionary Research; Córdoba-Aguilar, A., Ed.; Oxford University Press: New York, NY, USA, 2008; pp. 51–62. [Google Scholar]
- Herrando, S.; Brotons, L.; Anton, M.; Páramo, F.; Villero, D.; Titeux, N.; Quesada, X.; Stefanescu, C. Assessing impacts of land abandonment on Mediterranean biodiversity using indicators based on bird and butterfly monitoring data. Environm. Conserv. 2016, 43, 69–78. [Google Scholar] [CrossRef] [Green Version]
- ICNF 2022. Available online: https://www.icnf.pt/ (accessed on 2 December 2022).
- Maravalhas, E. As Borboletas de Portugal; Vento Norte: Porto, Portugal, 2003. [Google Scholar]
- Maravalhas, E.; Soares, A. The dragonflies of Portugal; Booky Publisher: Porto, Portugal, 2013. [Google Scholar]
- Serrano, A.R.M.; Soares, A.; Silva, A.; Fonseca, A.; Bívar de Sousa, A.; Figueiredo, A.; Rego, C.; Aguiar, C.A.S.; Monteiro, E.; Figueiredo, H.; et al. Biodiversidade, endemismos e espécies protegidas associadas às lagoas e cursos de água da Serra da Estrela: Valorização de um século de aproveitamento hidroeléctrico. Ecologi@ 2014, 7, 92–94. [Google Scholar]
- Habitats Directive. Council of the European Communities—Council Directive 92⁄43⁄EEC of 21 May 1992 on the conservation of natural habitats and of wild fauna and flora. Off. J. Eur. Communities 1992, 35, 7–50. [Google Scholar]
- Pollard, E.; Yates, T.J. Monitoring Butterflies for Ecology and Conservation; Chapman & Hall: London, UK, 1993. [Google Scholar]
- Hill, M.O. Diversity and evenness: A unifying notation and its consequences. Ecology 1973, 54, 427–432. [Google Scholar] [CrossRef] [Green Version]
- Chao, A.; Gotelli, N.J.; Hsieh, T.C.; Sander, E.L.; Ma, K.H.; Colwell, R.K.; Ellison, A.M. Rarefaction and extrapolation with Hill numbers: A framework for sampling and estimation in species diversity studies. Ecol. Monogr. 2014, 84, 45–67. [Google Scholar] [CrossRef] [Green Version]
- Carvalho, J.C.; Cardoso, P.; Gomes, P. Determining the relative roles of species replacement and species richness differences in generating beta-diversity patterns. Global Ecol. Biogeogr. 2012, 21, 760–771. [Google Scholar] [CrossRef]
- Cardoso, P.; Rigal, F.; Carvalho, J.C.; Fortelius, M.; Borges, P.A.V.; Podani, J.; Schmera, D. Partitioning taxon, phylogenetic and functional beta diversity into replacement and richness difference components. J. Biogeogr. 2014, 41, 749–761. [Google Scholar] [CrossRef] [Green Version]
- Borcard, D.; Legendre, P.; Drapeau, P. Partialling out the spatial component of ecological variation. Ecology 1992, 73, 1045–1055. [Google Scholar] [CrossRef] [Green Version]
- Fick, S.E.; Hijmans, R.J. Worldclim 2: New 1-km spatial resolution climate surfaces for global land areas. Int. J. Climat. 2017, 37, 4302–4315. [Google Scholar] [CrossRef]
- Borcard, D.; Gillet, F.; Legendre, P. Numerical Ecology with R; Springer: New York, NY, USA, 2018. [Google Scholar]
- Blanchet, F.G.; Legendre, P.; Borcard, D. Forward selection of explanatory variables. Ecology 2008, 89, 2623–2632. [Google Scholar] [CrossRef] [PubMed]
- Peres-Neto, P.R.; Legendre, P.; Dray, S.; Borcard, D. Variation partitioning of species data matrices: Estimation and comparison of fractions. Ecology 2006, 87, 2614–2625. [Google Scholar] [CrossRef]
- Legendre, P.; Borcard, D.; Peres-Neto, P.R. Analyzing beta diversity: Partitioning the spatial variation of community composition data. Ecol. Monogr. 2005, 75, 435–450. [Google Scholar] [CrossRef]
- Cardoso, P.; Rigal, F.; Carvalho, J.C. BAT: Biodiversity Assessment Tools. 2018, R Package Version 1.6.0. Available online: https://CRAN.R-project.org/package=BAT (accessed on 3 October 2022).
- Dinno, A. dunn.test: Dunn’s Test of Multiple Comparisons Using Rank Sums. R Package Version 1.3.5. 2017. Available online: https://CRAN.R-project.org/package=dunn.test (accessed on 3 October 2022).
- Oksanen, J.; Blanchet, F.G.; Friendly, M.; Kindt, R.; Legendre, P.; McGlinn, D.; Minchin, P.R.; O’Hara, R.B.; Simpson, G.L.; Solymos, P.; et al. Vegan: Community Ecology Package. R package version 2.5-2. 2018. Available online: https://CRAN.R-project.org/package=vegan (accessed on 3 October 2022).
- Dray, S.; Bauman, D.; Blanchet, G.; Borcard, D.; Clappe, S.; Guenard, G.; Jombart, T.; Larocque, G.; Legendre, P.; Madi, N.; et al. Adespatial: Multivariate Multiscale Spatial Analysis. 2021. R Package Version 0.3-14. Available online: https://CRAN.R-project.org/package=adespatial (accessed on 3 October 2022).
- R Core Team. R: A Language and Environment for Statistical Computing. R Foundation for Statistical Computing, Vienna, Austria. 2018. Available online: https://www.R-project.org/ (accessed on 3 October 2022).
- Boieiro, M.; Antunes, S.; Figueiredo, H.; Soares, A.; Lopes, A.; Monteiro, E.; Garcia-Pereira, P.; Rego, C.; Conde, J.; Borges, P.A.V.; et al. Standardised inventories of lepidopterans and odonates from Serra da Estrela Natural Park (Portugal)—Setting the scene for mountain biodiversity monitoring. Biodivers. Data J. 2023; submitted. [Google Scholar]
- McCain, C.M.; Grytnes, J.A. Elevational gradients in species richness. In Encyclopedia of Life Sciences; John Wiley & Sons Ltd.: Chichester, UK, 2010. [Google Scholar]
- Shen, C.; Liang, W.; Shi, Y.; Lin, X.; Zhang, H.; Wu, X.; Xie, G.; Chain, P.; Grogan, P.; Chu, H. Contrasting elevational diversity patterns between eukaryotic soil microbes and plants. Ecology 2014, 95, 3190–3202. [Google Scholar] [CrossRef] [Green Version]
- French, S.K.; McCauley, S.J. Canopy cover affects habitat selection by adult dragonflies. Hydrobiologia 2018, 818, 129–143. [Google Scholar] [CrossRef]
- Alves-Martins, F.; Calatayud, J.; Medina, N.G.; De Marco, P.; Juen, L.; Hortal, J. Drivers of regional and local diversity of Amazonian stream Odonata. Insect Conserv. Divers. 2019, 12, 251–261. [Google Scholar] [CrossRef]
- Nagy, H.B.; László, Z.; Szabó, F.; Szőcs, L.; Dévai, G.; Tóthmérész, B. Landscape-scale terrestrial factors are also vital in shaping Odonata assemblages of watercourses. Sci. Rep. 2019, 9, 18196. [Google Scholar] [CrossRef] [Green Version]
- Lawton, J.H.; MacGarvin, M.; Heads, P.A. Effects of altitude on the abundance and species richness of insect herbivores on bracken. J. Animal Ecol. 1987, 56, 147–160. [Google Scholar] [CrossRef]
- Molina-Martínez, A.; León-Cortés, J.L.; Regan, H.M. Climatic and geometric constraints as driving factors of butterfly species richness along a Neotropical elevational gradient. J. Insect Conserv. 2013, 17, 1169–1180. [Google Scholar] [CrossRef]
- Kaltsas, D.; Dede, K.; Giannaka, J.; Nasopoulou, T.; Kechagioglou, S.; Grigoriadou, E.; Raptis, D.; Damos, P.; Vasiliadis, I.; Christopoulos, V.; et al. Taxonomic and functional diversity of butterflies along an altitudinal gradient in two NATURA 2000 sites in Greece. Insect Conserv. Divers. 2018, 11, 464–478. [Google Scholar] [CrossRef]
- Hodkinson, I.D. Terrestrial insects along elevation gradients: Species and community responses to altitude. Biol. Rev. 2005, 80, 489–513. [Google Scholar] [CrossRef] [Green Version]
- Sanders, N.J.; Lessard, J.P.; Fitzpatrick, M.C.; Dunn, R.R. Temperature, but not productivity or geometry, predicts elevational diversity gradients in ants across spatial grains. Global Ecol. Biogeogr. 2007, 16, 640–649. [Google Scholar] [CrossRef]
- Musthafa, M.M.; Abdullah, F. Coleoptera of Genting Highland, Malaysia: Species richness and diversity changes along the elevations. Arxius de Miscellania Zoologica 2019, 17, 123–144. [Google Scholar] [CrossRef]
- Jansen, J. Guia Geobotânico da Serra da Estrela; Instituto da Conservação da Natureza: Lisboa, Portugal, 2002. [Google Scholar]
- Garcia-Barros, E.; Munguira, M.L.; Stefanescu, C.; Moreno, A.V. Lepidoptera: Papilionoidea; Fauna Iberica: Madrid, Spain, 2013. [Google Scholar]
- Stevens, G.C. The elevational gradient in altitudinal range: An extension of Rapoport’s latitudinal rule to altitude. Am. Nat. 1992, 140, 893–911. [Google Scholar] [CrossRef] [PubMed]
- Colwell, R.K.; Lees, D.C. The mid-domain effect: Geometric constraints on the geography of species richness. Trends Ecol. Evol. 2000, 15, 70–76. [Google Scholar] [CrossRef] [PubMed]
- Fontanilla, A.M.; Nakamura, A.; Xu, Z.; Cao, M.; Kitching, R.L.; Tang, Y.; Burwell, C.J. Taxonomic and functional ant diversity along tropical, subtropical, and subalpine elevational transects in Southwest China. Insects 2019, 10, 128. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Yen, J.D.L.; Fleishman, E.; Fogarty, F.; Dobkin, D.S. Relating beta diversity of birds and butterflies in the Great Basin to spatial resolution, environmental variables and trait-based groups. Global Ecol. Biogeogr. 2019, 28, 328–340. [Google Scholar] [CrossRef]
- Duan, M.; Liu, Y.; Yu, Z.; Baudry, J.; Li, L.; Wang, C.; Axmacher, C.J. Disentangling effects of abiotic factors and biotic interactions on cross-taxon congruence in species turnover patterns of plants, moths and beetles. Sci. Rep. 2016, 6, 23511. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Filazzola, A.; Matter, S.F.; Roland, J. Inclusion of trophic interactions increases the vulnerability of an alpine butterfly species to climate change. Glob Change Biol. 2020, 26, 2867–2877. [Google Scholar] [CrossRef] [PubMed]
- Jiang, X.; Chen, J.; Xie, J. Different roles of elevational and local environmental factors on abundance-based beta diversity of the soil Enchytraeidae on the Changbai Mountain. Ecol. Evol. 2019, 9, 2180–2188. [Google Scholar] [CrossRef] [Green Version]
- Carneiro, E.; Mielke, O.H.H.; Casagrande, M.M.; Fiedler, K. Community structure of skipper butterflies (Lepidoptera, Hesperiidae) along elevational gradients in Brazilian Atlantic Forest reflects vegetation type rather than altitude. PLoS ONE 2014, 9, e108207. [Google Scholar] [CrossRef] [Green Version]
- Lasmar, C.J.; Ribas, C.R.; Louzada, J.; Queiroz, A.C.; Feitosa, R.M.; Imata, M.M.; Alves, G.P.; Nascimento, G.B.; Neves, F.S.; Domingos, D.Q. Disentangling elevational and vegetational effects on ant diversity patterns. Acta Oecol. 2020, 102, 103489. [Google Scholar] [CrossRef]
- Dirnböck, T.; Essel, F.; Rabitsch, W. Disproportional risk for habitat loss of high-altitude endemic species under climate change. Glob. Chang. Biol. 2011, 17, 990–996. [Google Scholar] [CrossRef]
- Wilson, R.J.; Gutiérrez, D.; Gutiérrez, J.; Martínez, D.; Agudo, R.; Monserrat, V.J. Changes to the elevational limits and extent of species ranges associated with climate change. Ecol. Lett. 2005, 8, 1138–1146. [Google Scholar] [CrossRef] [PubMed]
- Cerrato, C.; Rocchia, E.; Brunetti, M.; Bionda, R.; Bassano, B.; Provenzale, A.; Bonelli, S.; Viterbi, R. Butterfly distribution along altitudinal gradients: Temporal changes over a short time period. Nat. Conserv. 2019, 34, 91–118. [Google Scholar] [CrossRef] [Green Version]
- Delpon, G.; Vogt-Schilb, H.; Munoz, F.; Richard, F.; Schatz, B. Diachronic variations in the distribution of butterflies and dragonflies linked to recent habitat changes in Western Europe. Insect Conserv. Divers. 2019, 12, 49–68. [Google Scholar] [CrossRef]
- Miranda, M.D.; Pereira, H.M.; Corley, M.F.V.; Merckx, T. Beta diversity patterns reveal positive effects of farmland abandonment on moth communities. Sci. Rep. 2019, 9, 1549. [Google Scholar] [CrossRef] [PubMed]
- Eskildsen, A.; Carvalheiro, L.G.; Kissling, W.D.; Biesmeijer, J.C.; Schweiger, O.; Høye, T.T. Ecological specialization matters: Long-term trends in butterfly species richness and assemblage composition depend on multiple functional traits. Divers. Distribut. 2015, 21, 792–802. [Google Scholar] [CrossRef]




{kind=link}
{kind=link}
{kind=link}
| Butterflies | Odonates | |
|---|---|---|
| Species Richness | 16–48 (31.3 ± 8.9) | 3–14 (7.4 ± 3.2) |
| Abundance | 88–409 (186.7 ± 96.0) | 17–164 (88.4 ± 48.3) |
| EShannon | 10.3–33.0 (19.5 ± 6.1) | 2.1–8.4 (4.6 ± 1.7) |
| ISimpson | 7.2–26.2 (13.9 ± 15.2) | 1.8–6.1 (3.7 ± 1.3) |
| Berger–Parker | 0.07–0.27 (0.17 ± 0.06) | 0.30–0.71 (0.45 ± 0.13) |
| Evenness | 0.48–0.61 (0.53 ± 0.04) | 0.27–0.74 (0.43 ± 0.16) |
| Estimated Species Richness | 16.0–53.6 (36.2 ± 10.9) | 3.0–15.5 (7.8 ± 3.6) |
| Completeness | 0.73–1.00 (0.88 ± 0.08) | 0.88–1.00 (0.97 ± 0.05) |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Ceia-Hasse, A.; Boieiro, M.; Soares, A.; Antunes, S.; Figueiredo, H.; Rego, C.; Borges, P.A.V.; Conde, J.; Serrano, A.R.M. Drivers of Insect Community Change along the Margins of Mountain Streams in Serra da Estrela Natural Park (Portugal). Insects 2023, 14, 243. https://doi.org/10.3390/insects14030243
Ceia-Hasse A, Boieiro M, Soares A, Antunes S, Figueiredo H, Rego C, Borges PAV, Conde J, Serrano ARM. Drivers of Insect Community Change along the Margins of Mountain Streams in Serra da Estrela Natural Park (Portugal). Insects. 2023; 14(3):243. https://doi.org/10.3390/insects14030243
Chicago/Turabian StyleCeia-Hasse, Ana, Mário Boieiro, Albano Soares, Sandra Antunes, Hugo Figueiredo, Carla Rego, Paulo A.V. Borges, José Conde, and Artur R.M. Serrano. 2023. "Drivers of Insect Community Change along the Margins of Mountain Streams in Serra da Estrela Natural Park (Portugal)" Insects 14, no. 3: 243. https://doi.org/10.3390/insects14030243