
Effects of Environmental Factors on the Spatial Distribution Pattern and Diversity of Insect Communities along Altitude Gradients in Guandi Mountain, China
Abstract
:Simple Summary
Abstract
1. Introduction
2. Materials and Methods
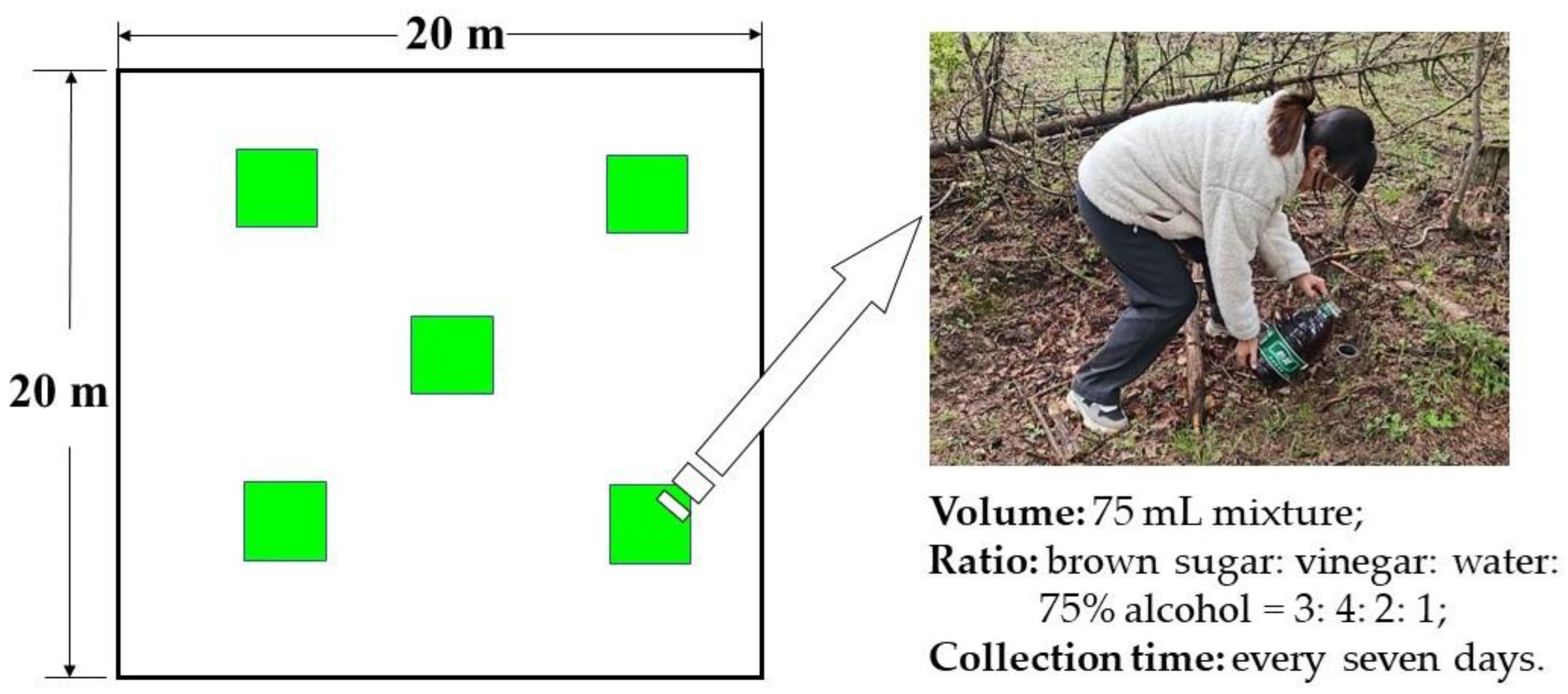
2.1. Study Sites
2.2. Insect Sampling and Specimen Identification
2.3. Environmental Data Collection
2.4. Data Analysis
3. Results
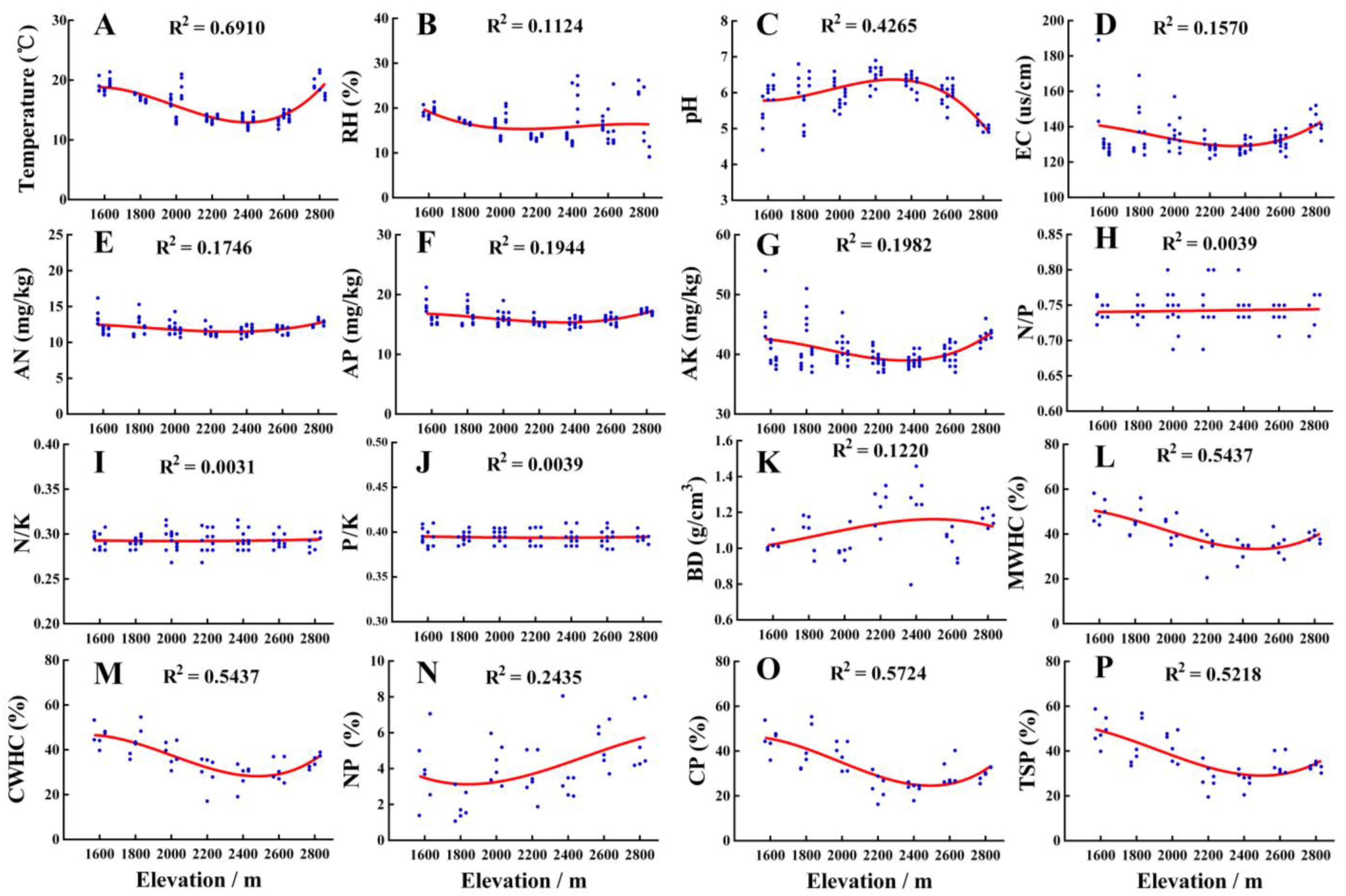
3.1. Variations of Environmental Variables along the Altitudinal Gradient
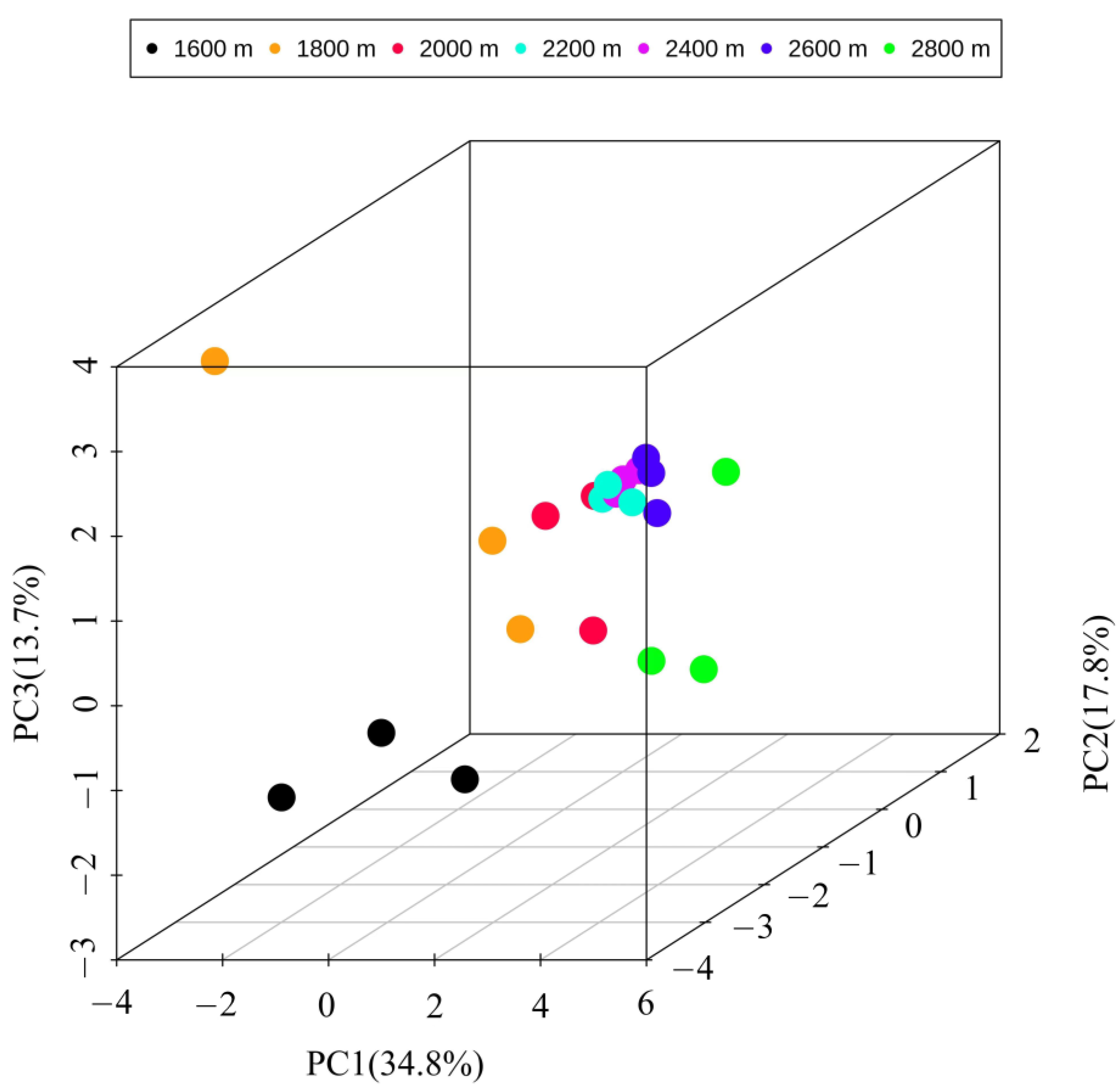
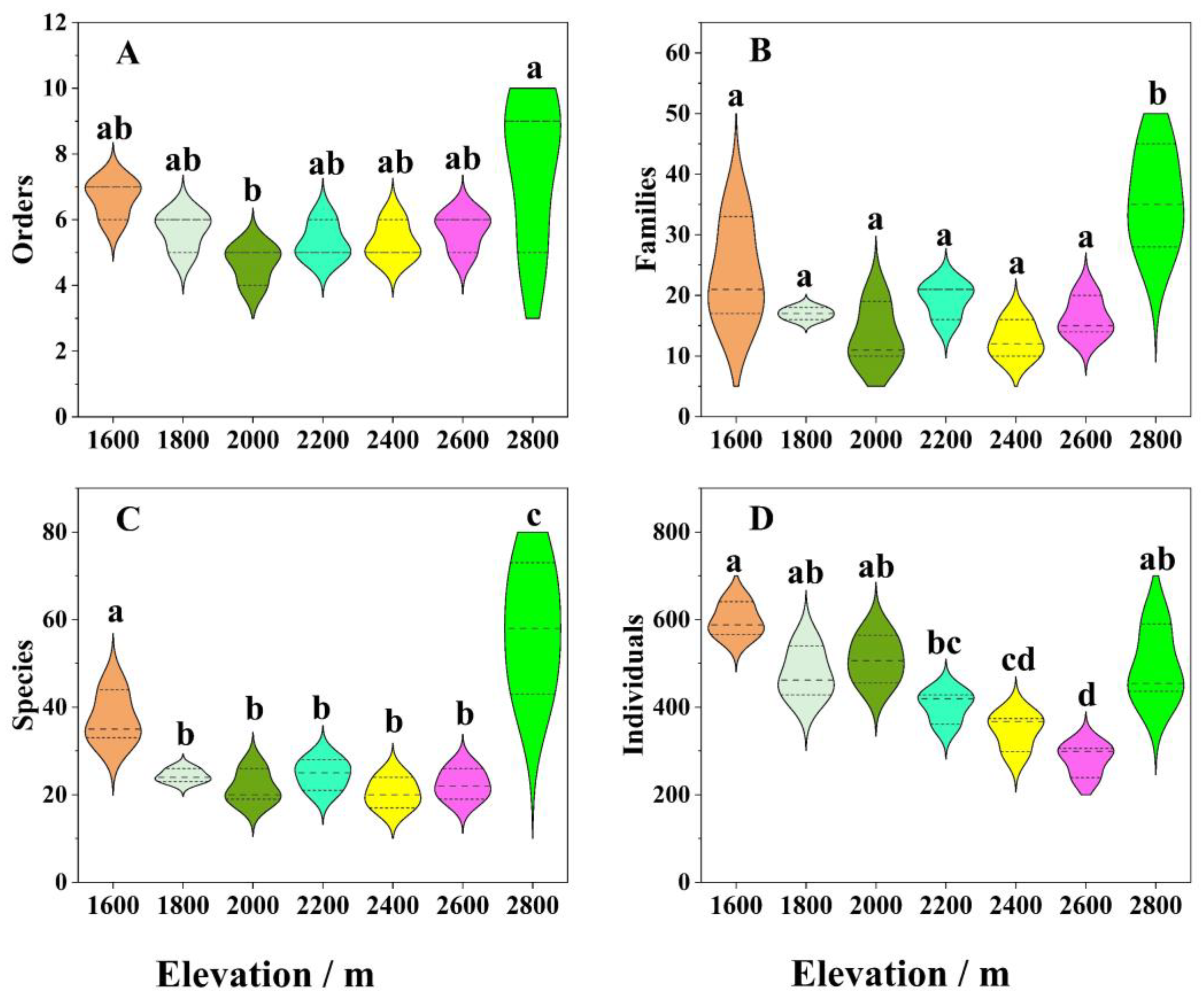
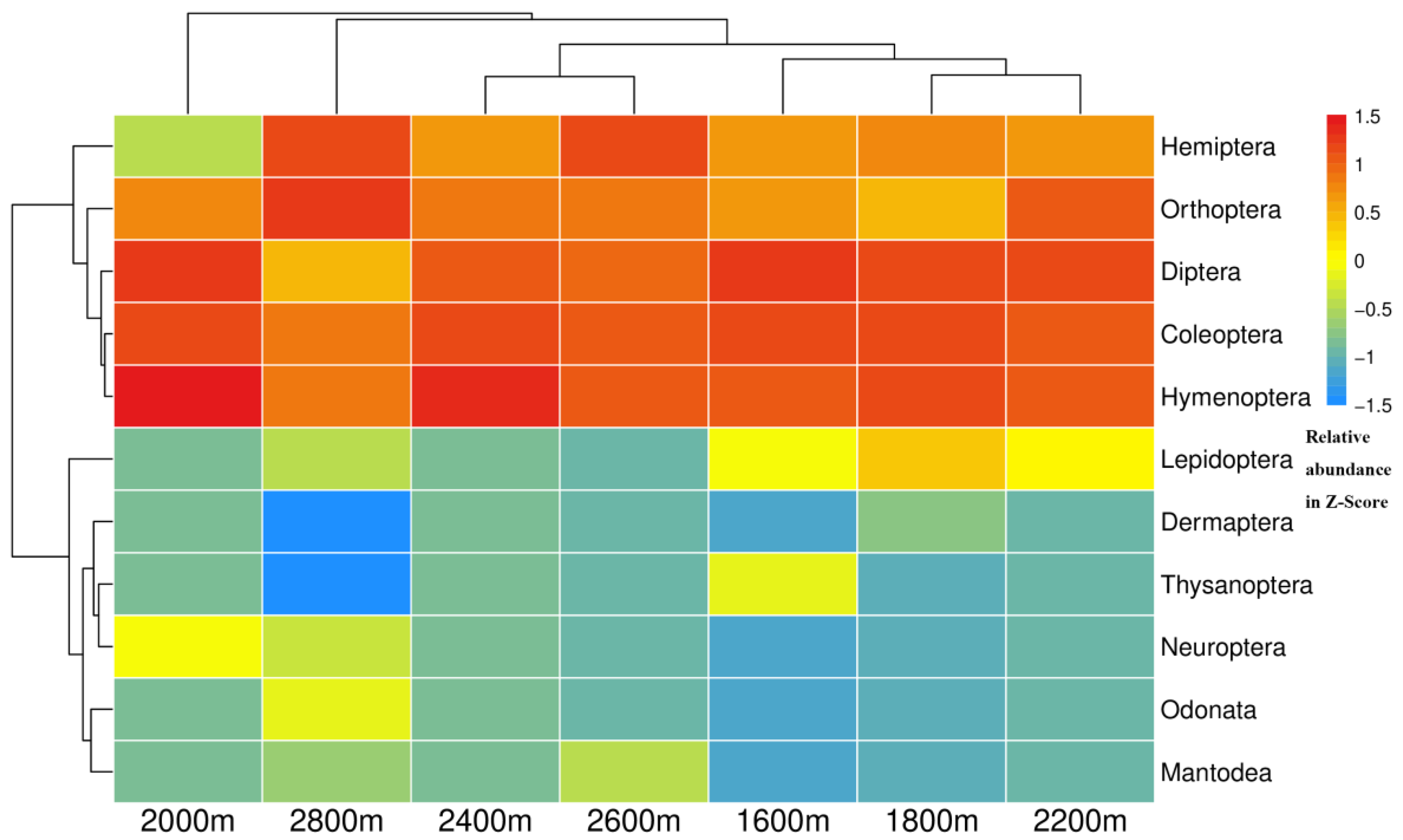
3.2. Variations in Insect Community Composition along the Altitudinal Gradient
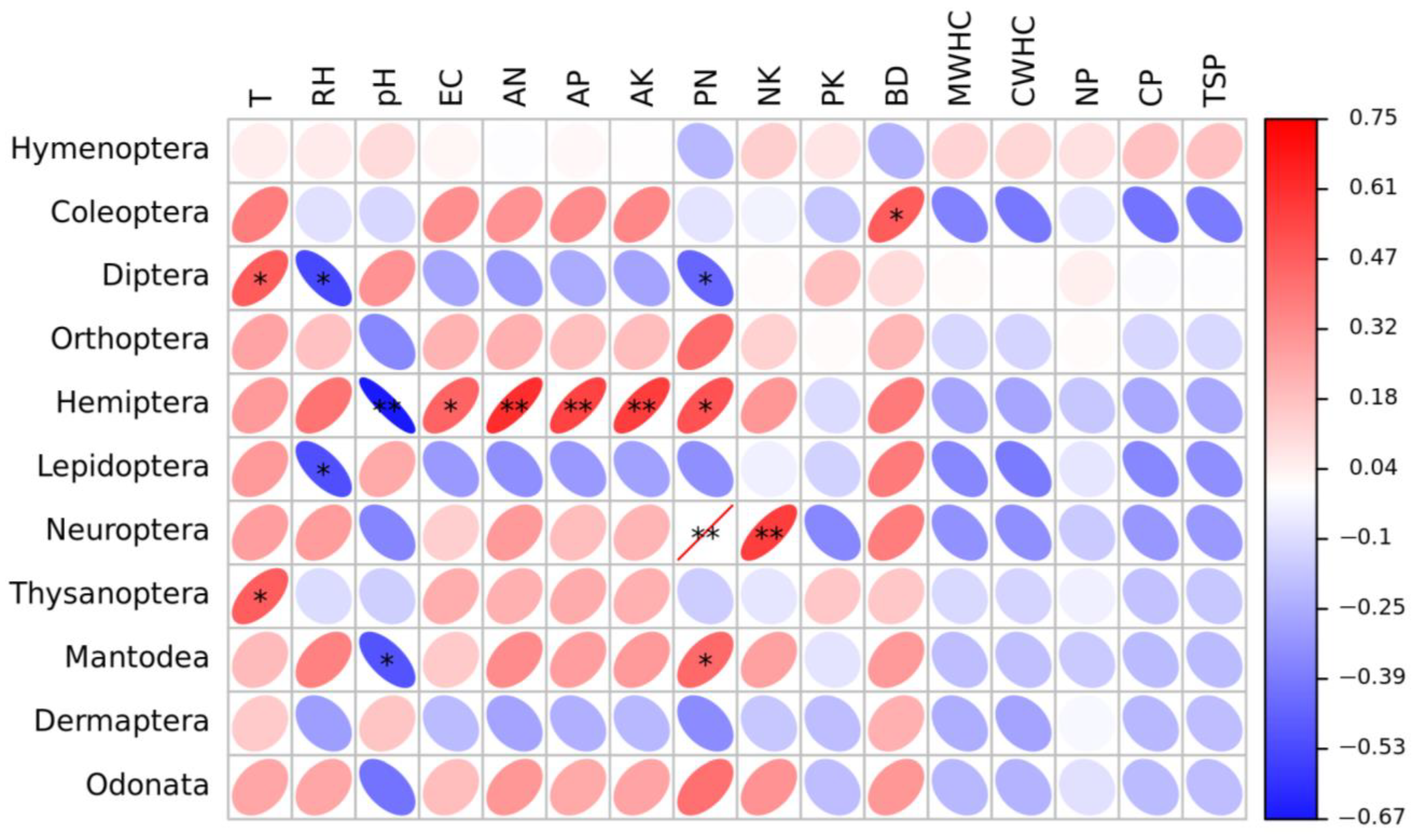
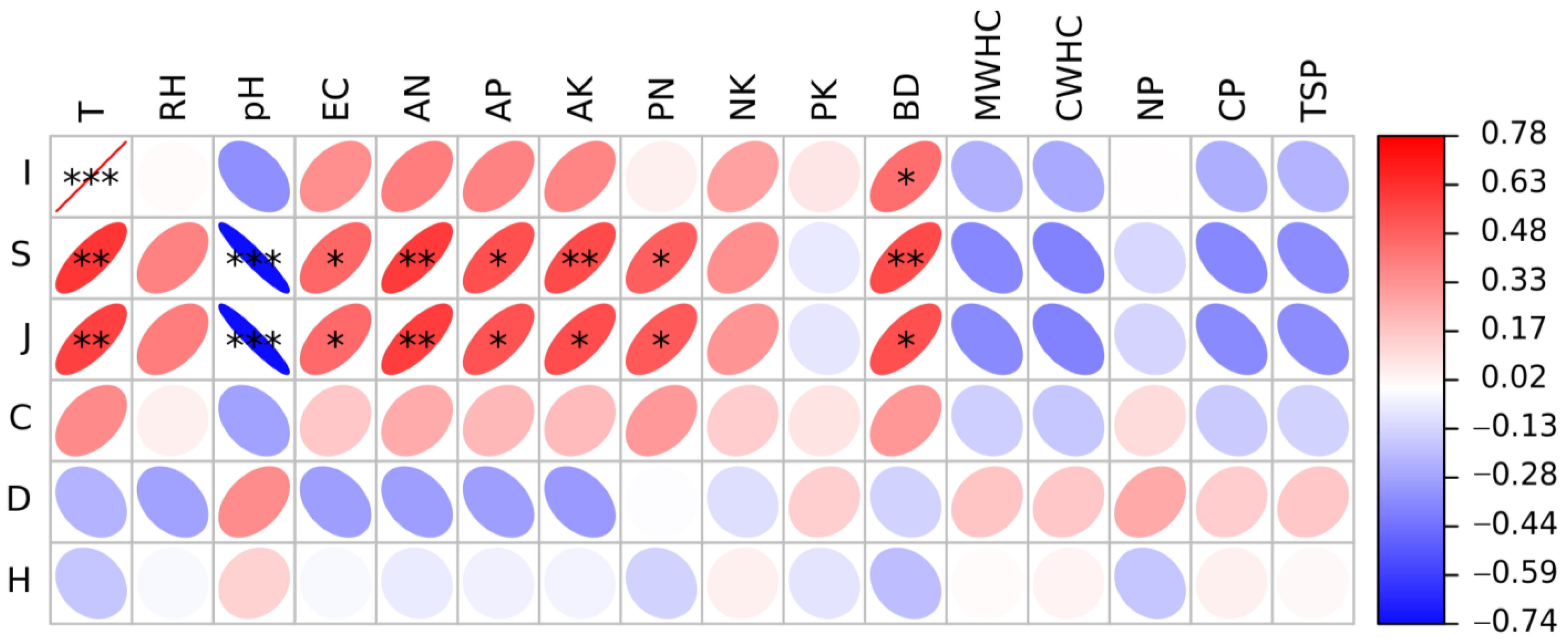
3.3. Correlation of Environmental Variables with Insect Community and Diversity
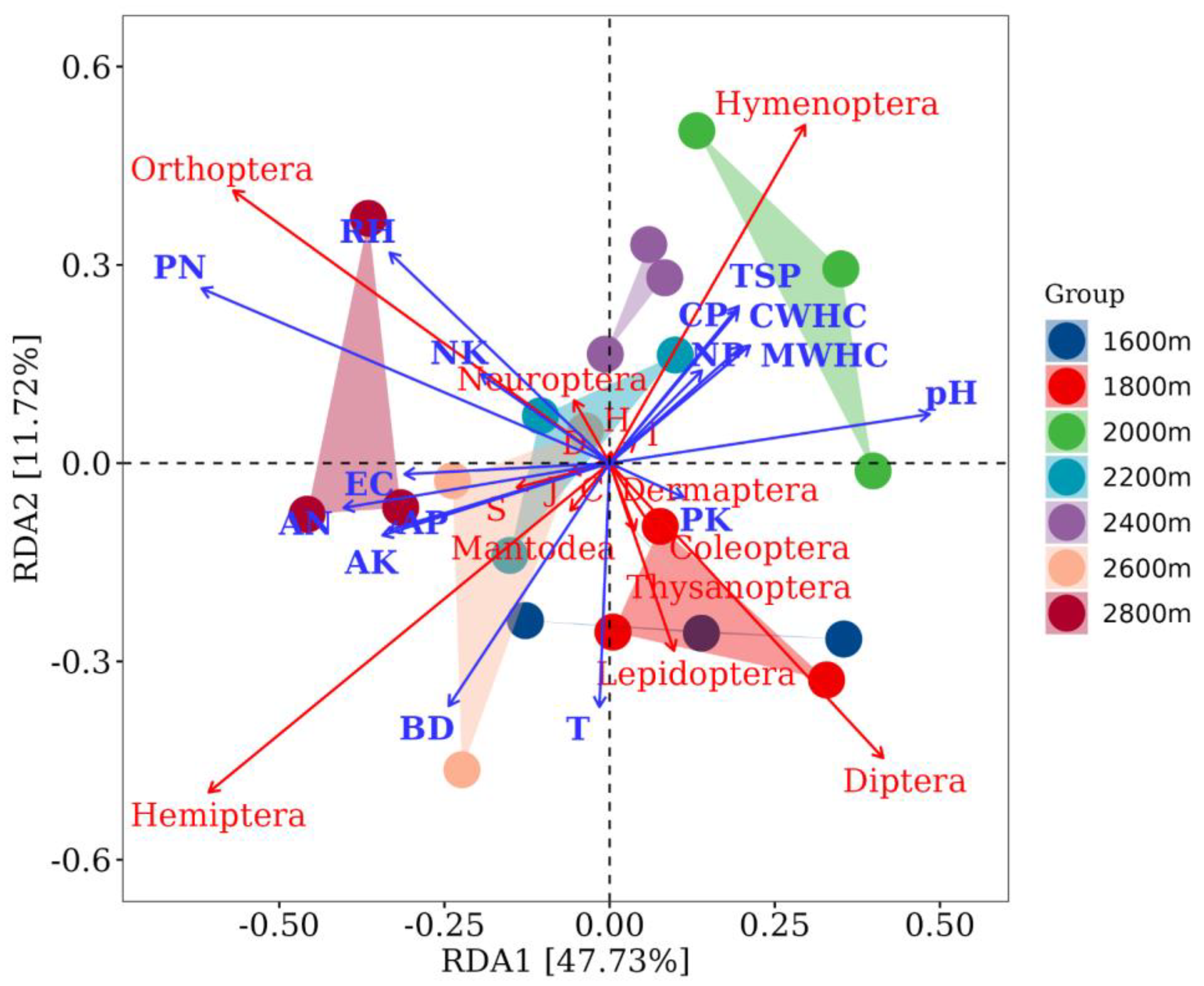
3.4. Effects of Environmental Variables on the Spatial Distribution and Diversity of Insect Communities along Altitudinal Gradients
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Biaggini, M.; Consorti, R.; Dapporto, L. The taxonomic level order as a possible tool for rapid assessment of Arthropod diversity in agricultural landscapes. Agric. Ecosyst. Environ. 2007, 122, 183–191. [Google Scholar] [CrossRef]
- Maguire, D.Y.; James, P.M.A.; Buddle, C.M.; Bennett, E.M. Landscape connectivity and insect herbivory: A framework for understanding tradeoffs among ecosystem services. Glob. Ecol. Conserv. 2015, 4, 73–84. [Google Scholar] [CrossRef] [Green Version]
- Shao, X.; Zhang, Q.; Yang, X. Spatial patterns of insect herbivory within a forest landscape: The role of soil type and forest stratum. For. Ecosyst. 2021, 8, 1–14. [Google Scholar] [CrossRef]
- Nahmani, J.; Lavelle, P.; Rossi, J.P. Does changing the taxonomical resolution alter the value of soil macroinvertebrates as bioindicators of metal pollution? Soil Biol. Biochem. 2006, 38, 385–396. [Google Scholar] [CrossRef]
- Tom, R.; Bishop, M.P.; Robertson, B.J.; Van, R.; Catherine, L.P. Elevation–diversity patterns through space and time: Ant communities of the Maloti-Drakensberg Mountains of southern Africa. J. Biogeogr. 2014, 41, 2256–2268. [Google Scholar]
- Rasmann, S.; Pellissier, L.; Defossez, E.; Jactel, H.; Kunstler, G.; Bailey, J.K. Climate-driven change in plant-insect interactions along elevation gradients. Funct. Ecol. 2014, 28, 46–54. [Google Scholar] [CrossRef]
- Gao, R.; Shi, J.; Huang, R.; Wang, Z.; Luo, Y. Effects of pine wilt disease invasion on soil properties and Masson pine forest communities in the Three Gorges reservoir region, China. Ecol. Evol. 2015, 5, 1702–1716. [Google Scholar] [CrossRef]
- Hodkinson, I.D. Terrestrial insects along elevation gradients: Species and community responses to altitude. Biol. Rev. 2005, 80, 489–513. [Google Scholar] [CrossRef] [Green Version]
- Moreira, X.; Abdala-Roberts, L.; Rasmann, S. Plant diversity effects on insect herbivores and their natural enemies: Current thinking, recent findings, and future directions. Curr. Opin. Insect Sci. 2016, 14, 1–7. [Google Scholar] [CrossRef] [Green Version]
- Cuevas-Reyes, P.; Quesada, M.; Hanson, P.; Dirzo, R.; Oyama, K. Diversity of gall-inducing insects in a Mexican tropical dry forest: The importance of plant species richness, life-forms, host plant age and plant density. J. Ecol. 2004, 92, 707–716. [Google Scholar] [CrossRef]
- Klingauf, F. Interrelations between pests and climatic factors. In Food-Climate Interactions; Bach, W., Pankrath, J., Schneider, S.H., Eds.; Springer: Dordrecht, The Netherlands, 1981; pp. 285–301. [Google Scholar]
- Heinen, R.; Biere, A.; Harvey, J.A.; Bezemer, T. Effects of Soil Organisms on Aboveground Plant-Insect Interactions in the Field: Patterns, Mechanisms and the Role of Methodology. Front. Ecol. Evol. 2018, 6, 106. [Google Scholar] [CrossRef] [Green Version]
- Jaworski, T.; Hilszczański, J. The effect of temperature and humidity changes on insects development and their impact on forest ecosystems in the context of expected climate change. For. Res. Pap. 2013, 74, 345–355. [Google Scholar]
- Guo, Q.; Fei, S.; Potter, K.M.; Liebhold, A.M.; Wen, J. Tree diversity regulates forest pest invasion. Proc. Natl. Acad. Sci. USA 2019, 116, 7382–7386. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Jactel, H.; Koricheva, J.; Castagneyrol, B. Responses of forest insect pests to climate change: Not so simple. Curr. Opin. Insect Sci. 2019, 35, 103–108. [Google Scholar] [CrossRef] [PubMed]
- Lehmann, P.; Ammunét, T.; Barton, M.; Battisti, A.; Eigenbrode, S.D.; Jepsen, J.U.; Kalinkat, G.; Neuvonen, S.; Niemelä, P.; Terblanche, J.S.; et al. Complex responses of global insect pests to climate warming. Front. Ecol. Environ. 2020, 18, 141–150. [Google Scholar] [CrossRef] [Green Version]
- Vergara, G.D.; Williams, L.G.; Casanoves, F. Leaf functional traits vary within and across tree species in tropical cloud forest on rock outcrop versus volcanic soil. J. Veg. Sci. 2019, 31, 129–138. [Google Scholar] [CrossRef]
- Cisneros, J.J.; Godfrey, L.D. Midseason pest status of the cotton aphid (Homoptera: Aphididae) in California cotton: Is nitrogen a key factor? Popul. Ecol. 2001, 30, 501–510. [Google Scholar] [CrossRef] [Green Version]
- Stiling, P.; Moon, D.C. Quality or quantity: The direct and indirect effects of host plants on herbivores and their natural enemies. Oecologia 2004, 142, 413–420. [Google Scholar] [CrossRef]
- Huberty, A.F.; Denno, R.F. Consequences of nitrogen and phosphorus limitation for the performance of two planthoppers with divergent life history strategies. Oecologia 2006, 149, 444–455. [Google Scholar] [CrossRef]
- Körner, C. The use of ‘altitude’ in ecological research. Trends Ecol. Evol. 2007, 22, 569–574. [Google Scholar] [CrossRef]
- Peters, M.; Hemp, A.; Appelhans, T.; Behler, C.; Classen, A.; Detsch, F.; Ensslin, A.; Ferger, S.W.; Frederiksen, S.B.; Gebert, F. Predictors of elevational biodiversity gradients change from single taxa to the multi-taxa community level. Nat. Commun. 2016, 7, 13736. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Stiegel, S.; Entling, M.H.; Mantilla, C.J. Reading the leaves’ palm: Leaf traits and herbivory along the microclimatic gradient of forest layers. PLoS ONE 2017, 12, e0169741. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Mantoni, C.; Tsafack, N.; Palusci, E. Diversity Patterns of Dung Beetles along a Mediterranean Elevational Gradient. Insects 2021, 12, 781. [Google Scholar] [CrossRef]
- Keil, P.; Konvicka, M. Local species richness of Central European hoverflies (Diptera: Syrphidae): A lesson taught by local faunal lists. Divers. Distrib. 2005, 11, 417–426. [Google Scholar] [CrossRef]
- Laiolo, P.; Pato, J.; Obeso, J.R. Ecological and evolutionary drivers of the elevational gradient of diversity. Ecol. Lett. 2018, 21, 1022–1032. [Google Scholar] [CrossRef] [Green Version]
- Laossi, K.R.; Barot, S.; Carvalho, D.; Lavelle, P.; Martins, M.; Mitja, D.; Rendeiro, A.C.; Roussin, G.; Sarazin, M. Effects of plant diversity on plant biomass production and soil macro fauna in Amazonian pastures. Pedobiologia 2008, 51, 397–407. [Google Scholar] [CrossRef]
- Yin, X.; Qiu, L.; Jiang, Y.; Wang, Y. Diversity and Spatial-Temporal Distribution of Soil Macrofauna Communities Along Elevation in the Changbai Mountain, China. Environ. Entomol. 2017, 46, 454–459. [Google Scholar] [CrossRef]
- Jiang, Y.F.; Yin, X.Q.; Wang, F.B. Composition and Spatial Distribution of Soil Mesofauna Along an Elevation Gradient on the North Slope of the Changbai Mountains, China. Pedosphere 2015, 25, 811–824. [Google Scholar] [CrossRef]
- Fattorini, S.; Mantoni, C.; Di Biase, L.; Strona, G.; Pace, L.; Biondi, M. Elevational patterns of generic diversity in the tenebrionid beetles (Coleoptera Tenebrionidae) of Latium (Central Italy). Diversity 2020, 12, 47. [Google Scholar] [CrossRef] [Green Version]
- The National Meteorological Information Center (NMIC) of China Meteorological Administration (CMA). Available online: http://www.data.cma.cn (accessed on 15 February 2023).
- Wang, Z.; Zhao, L.; Liu, J.; Yang, Y.; Shi, J.; Wen, J.; Gao, R. Functional relationship between woody plants and insect communities in response to Bursaphelenchus xylophilus infestation in the Three Gorges Reservoir region. Ecol. Evol. 2021, 11, 8843–8855. [Google Scholar] [CrossRef]
- Zhai, H.; Yu, X.M.; Ma, Y.A.; Zhang, Y.; Wang, D. Sugar–Acetic Acid–Ethanol–Water Mixture as a Potent Attractant for Trapping the Oriental Fruit Moth (Lepidoptera: Tortricidae) in Peach–Apple Mixed-Planting Orchards. Plants 2019, 8, 401. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Yang, M.F.; Mang, Z.H.; Li, Z.Z. Fauna Sinica Insecta; Science Press: Beijing, China, 2017; Volume 67, p. 637. [Google Scholar]
- Fan, Z.D.; Deng, Y.H. Fauna Sinica Insecta; Science Press: Beijing, China, 2008; Volume 49, p. 1186. [Google Scholar]
- He, J.H.; Chen, X.X.; Ma, Y. Fauna Sinica Insecta; Science Press: Beijing, China, 2000; Volume 18, p. 757. [Google Scholar]
- Han, H.X.; Xue, D.Y. Fauna Sinica Insecta; Science Press: Beijing, China, 2011; Volume 54, p. 787. [Google Scholar]
- Ren, G.D.; Liu, H.Y. Fauna Sinica Insecta; Science Press: Beijing, China, 2016; Volume 63, p. 534. [Google Scholar]
- Li, M.L. Resource Entomology; China Forestry Publishing House: Beijing, China, 2005. (In Chinese) [Google Scholar]
- Chou, I. Monographia Rhopalocerorum Sinensium; Henan Scientific and Technological Publishing House: Zhengzhou, China, 2000. (In Chinese) [Google Scholar]
- Hebert, P.; Cywinska, A.; Ball, S.L.; Dewaard, J.R. Biological identification through DNA barcodes. Proc. Biol. Sci. 2003, 270, 313–321. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zhang, Y.S.; Liu, S.; Ma, J. Water-holding capacity of ground covers and soils in alpine and sub-alpine shrubs in western Sichuan, China. Acta Ecol. 2006, 26, 2775–2781. [Google Scholar] [CrossRef]
- Hill, M.O. Diversity and evenness: A unifying notation and its consequences. Ecology 1973, 54, 427–432. [Google Scholar] [CrossRef] [Green Version]
- Jost, L. Entropy and diversity. Oikos 2006, 113, 363–375. [Google Scholar] [CrossRef]
- Magurran, A.E. Measuring Ecological Diversity; Blackwell Publishing: Oxford, UK, 2004; p. 256. [Google Scholar]
- Tuomisto, H. A consistent terminology for quantifying species diversity? Yes, it does exist. Oecologia 2010, 164, 853–860. [Google Scholar] [CrossRef]
- Rasmann, S.; Agrawal, A.A. Latitudinal patterns in plant defense: Evolution of cardenolides, their toxicity and induction following herbivory. Ecol. Lett. 2011, 14, 476–483. [Google Scholar] [CrossRef]
- Sundqvist, M.K.; Sanders, N.J.; Wardle, D.A. Community and ecosystem responses to elevational gradients: Processes, mechanisms, and insights for global change. Annu. Rev. Ecol. Evol. Syst. 2013, 44, 261–280. [Google Scholar] [CrossRef] [Green Version]
- Jump, A.S.; Mátyás, C.; Peñuelas, J. The altitude for latitude disparity in the range retractions of woody species. Trends Ecol. Evol. 2009, 24, 694–701. [Google Scholar] [CrossRef] [Green Version]
- Chatelain, P.; Plant, A.; Soulier-Perkins, A. Diversity increases with elevation: Empidine dance flies (Diptera: Empididae) challenge a predominant pattern. Biotropica 2018, 50, 633–640. [Google Scholar] [CrossRef]
- Khairul, H.N.; Noor Nasuha, A.A.; Saiyid Jalaluddin, S.S. Diversity and abundance of dipteran species at two different elevations in Gunung Datuk, Negeri Sembilan, Malaysia. Serangga 2018, 23, 194–202. [Google Scholar]
- Plant, A.R.; Bickel, D.J.; Chatelain, P. Composition and organization of highly speciose Empidoidea (Diptera) communities in tropical montane forests of northern Thailand. Zootaxa 2019, 4590, 1–39. [Google Scholar] [CrossRef] [PubMed]
- Jobidon, R.; Cyr, G.; Thiffault, N. Plant species diversity and composition along an experimental gradient of northern hardwood abundance in Picea mariana plantations. For. Ecol. Manag. 2004, 198, 209–221. [Google Scholar] [CrossRef]
- Ricklefs, R.E.; Marquis, R.J. Species richness and niche space for temperate and tropical folivores. Oecologia 2012, 168, 213–220. [Google Scholar] [CrossRef]
- Neves, F.S.; Silva, J.O.; Espírito-Santo, M.M.; Fernandes, G.W. Insect herbivores and leaf damage along successional and vertical gradients in a tropical dry forest. Biotropica 2014, 46, 14–24. [Google Scholar] [CrossRef]
- Choi, S.W. Diversity and composition of larger moths in three different forest types of Southern Korea. Ecol. Res. 2008, 23, 503–509. [Google Scholar] [CrossRef]
- Alalouni, U.; Brandl, R.; Auge, H. Does insect herbivory on oak depend on the diversity of tree stands? Basic Appl. Ecol. 2014, 15, 685–692. [Google Scholar] [CrossRef]
- Root, R.B. Organization of a plant-arthropod association in simple and diverse habitats: The fauna of collards (Brassica oleracea). Ecol. Monogr. 1973, 43, 95–124. [Google Scholar] [CrossRef]
- Novotny, V.; Drozd, P.; Miller, S.E.; Kulfan, M.; Janda, M.; Basset, Y.; Weiblen, G.D. Why are there so many species of herbivorous insects in tropical rainforests? Science 2006, 313, 1115–1118. [Google Scholar] [CrossRef] [Green Version]
- Lewinsohn, T.M.; Roslin, T. Four ways towards tropical herbivore megadiversity. Ecol. Lett. 2008, 11, 398–416. [Google Scholar] [CrossRef]
- Savopoulou-Soultani, M.; Papadopoulos, N.T.; Milonas, P.; Moyal, P. Abiotic factors and insect abundance. Psyche-J. Entomol. 2012, 12, 1–2. [Google Scholar] [CrossRef] [Green Version]
- Fine, P.V.A. Ecological and evolutionary drivers of geographic variation in species diversity. Annu. Rev. Ecol. Evol. Syst. 2015, 46, 369–392. [Google Scholar] [CrossRef] [Green Version]









{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Vegetation Community a | Elevation (m) | Slope (°) | Plant Coverage (%) | Number of Individuals (Tree ha−1) | Mean DBH b (cm) | Mean Tree Height (m) |
|---|---|---|---|---|---|---|
| QWF | 1600 | 26.16 ± 1.41 | 82.67 ± 2.52 | 1058.33 ± 101.04 | 9.36 ± 2.00 | 9.12 ± 2.55 |
| PTF | 1800 | 21.25 ± 2.02 | 81.67 ± 10.12 | 741.67 ± 128.29 | 17.44 ± 0.65 | 13.67 ± 2.43 |
| PBM | 2000 | 12.93 ± 5.20 | 73.50 ± 2.12 | 516.67 ± 14.43 | 17.15 ± 3.57 | 12.62 ± 1.55 |
| PWF | 2200 | 18.62 ± 6.25 | 82.33 ± 4.04 | 816.67 ± 80.36 | 16.98 ± 2.73 | 14.89 ± 1.80 |
| PLF | 2400 | 23.33 ± 4.16 | 77.67 ± 2.08 | 483.33 ± 38.19 | 30.82 ± 4.95 | 18.31 ± 3 |
| LPF | 2600 | 26.36 ± 6.53 | 76.33 ± 1.53 | 491.67 ± 87.80 | 25.38 ± 2.46 | 16.54 ± 2.78 |
| SAM | 2800 | 17.16 ± 3.28 | 92.67 ± 1.56 |
| Order | Family | Species | Individual | |||
|---|---|---|---|---|---|---|
| Number | Percentage/% | Number | Percentage/% | Number | Percentage/% | |
| Coleoptera | 23 | 28.75 | 73 | 33.03 | 2118 | 22.72 |
| Diptera | 18 | 22.50 | 35 | 15.84 | 2068 | 22.19 |
| Orthoptera | 11 | 13.75 | 35 | 15.84 | 1214 | 13.02 |
| Hemiptera | 11 | 13.75 | 33 | 14.93 | 1064 | 11.42 |
| Hymenoptera | 7 | 8.75 | 21 | 9.50 | 2656 | 28.49 |
| Lepidoptera | 4 | 5.00 | 18 | 8.14 | 109 | 1.17 |
| Neuroptera | 2 | 2.50 | 2 | 0.90 | 22 | 0.24 |
| Dermaptera | 1 | 1.25 | 1 | 0.45 | 16 | 0.17 |
| Odonata | 1 | 1.25 | 1 | 0.45 | 6 | 0.06 |
| Mantodea | 1 | 1.25 | 1 | 0.45 | 9 | 0.10 |
| Thysanoptera | 1 | 1.25 | 1 | 0.45 | 39 | 0.42 |
| Total | 80 | 100 | 221 | 100 | 9321 | 100 |
| Elevation (m) | Species Richness | Shannon–Wiener Index | Inverse Simpson’s Index | Berger–Parker Index |
|---|---|---|---|---|
| 1600 | 38.00 ± 7.00 a | 3.26 ± 0.21 a | 0.05 ± 0.01 a | 0.12 ± 0.04 a |
| 1800 | 24.33 ± 1.53 b | 2.66 ± 0.45 b | 0.11 ± 0.07 ab | 0.19 ± 0.09 ab |
| 2000 | 22.67 ± 3.06 b | 2.41 ± 0.10 b | 0.15 ± 0.03 b | 0.31 ± 0.08 b |
| 2200 | 25.00 ± 4.00 b | 2.76 ± 0.47 ab | 0.11 ± 0.08 ab | 0.25 ± 0.15 ab |
| 2400 | 22.00 ± 4.36 b | 2.51 ± 0.29 ab | 0.13 ± 0.06 ab | 0.28 ± 0.12 ab |
| 2600 | 22.33 ± 3.51 b | 2.77 ± 0.35 ab | 0.08 ± 0.04 ab | 0.16 ± 0.05 ab |
| 2800 | 60.67 ± 14.19 c | 3.12 ± 0.31 ab | 0.08 ± 0.03 ab | 0.18 ± 0.05 ab |
| p-value | 0.00 | 0.07 | 0.29 | 0.21 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Zhao, L.; Gao, R.; Liu, J.; Liu, L.; Li, R.; Men, L.; Zhang, Z. Effects of Environmental Factors on the Spatial Distribution Pattern and Diversity of Insect Communities along Altitude Gradients in Guandi Mountain, China. Insects 2023, 14, 224. https://doi.org/10.3390/insects14030224
Zhao L, Gao R, Liu J, Liu L, Li R, Men L, Zhang Z. Effects of Environmental Factors on the Spatial Distribution Pattern and Diversity of Insect Communities along Altitude Gradients in Guandi Mountain, China. Insects. 2023; 14(3):224. https://doi.org/10.3390/insects14030224
Chicago/Turabian StyleZhao, Lijuan, Ruihe Gao, Jiaqi Liu, Lei Liu, Rongjiao Li, Lina Men, and Zhiwei Zhang. 2023. "Effects of Environmental Factors on the Spatial Distribution Pattern and Diversity of Insect Communities along Altitude Gradients in Guandi Mountain, China" Insects 14, no. 3: 224. https://doi.org/10.3390/insects14030224