


Floral Scents in Bee-Pollinated Buckwheat and Oilseed Rape under a Global Warming Scenario

Abstract
:Simple Summary
Abstract
1. Introduction
2. Material and Methods
2.1. Crop Plant Cultivation and Temperature Regime
2.2. Sampling and Analysis of Flower Scents
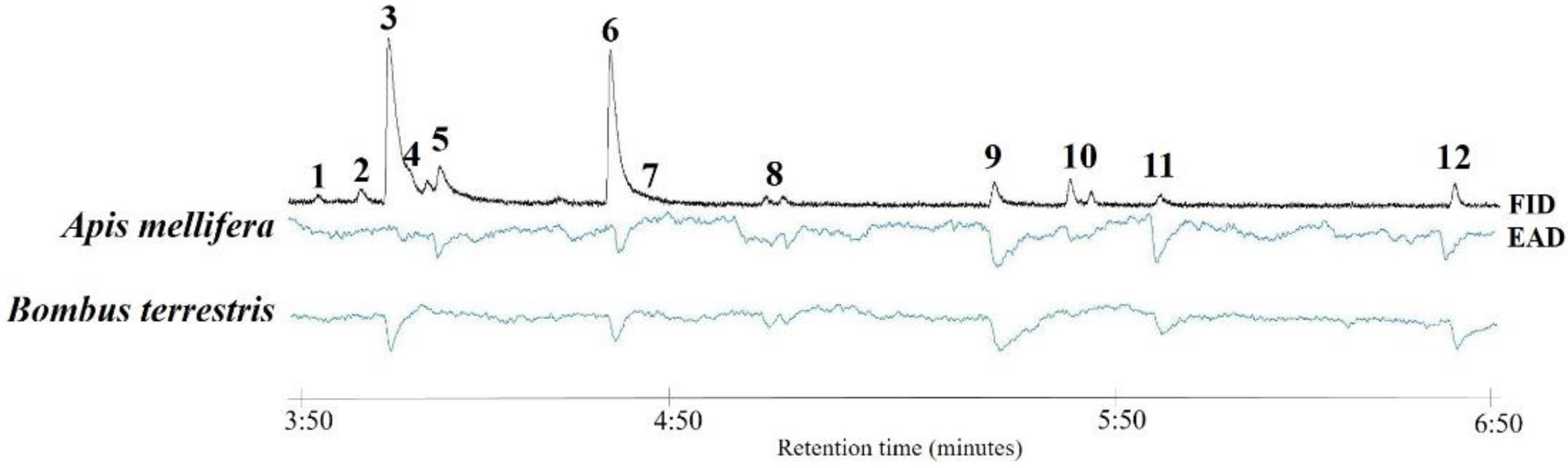
2.3. Electroantennographic Detection
2.4. Data Analyses
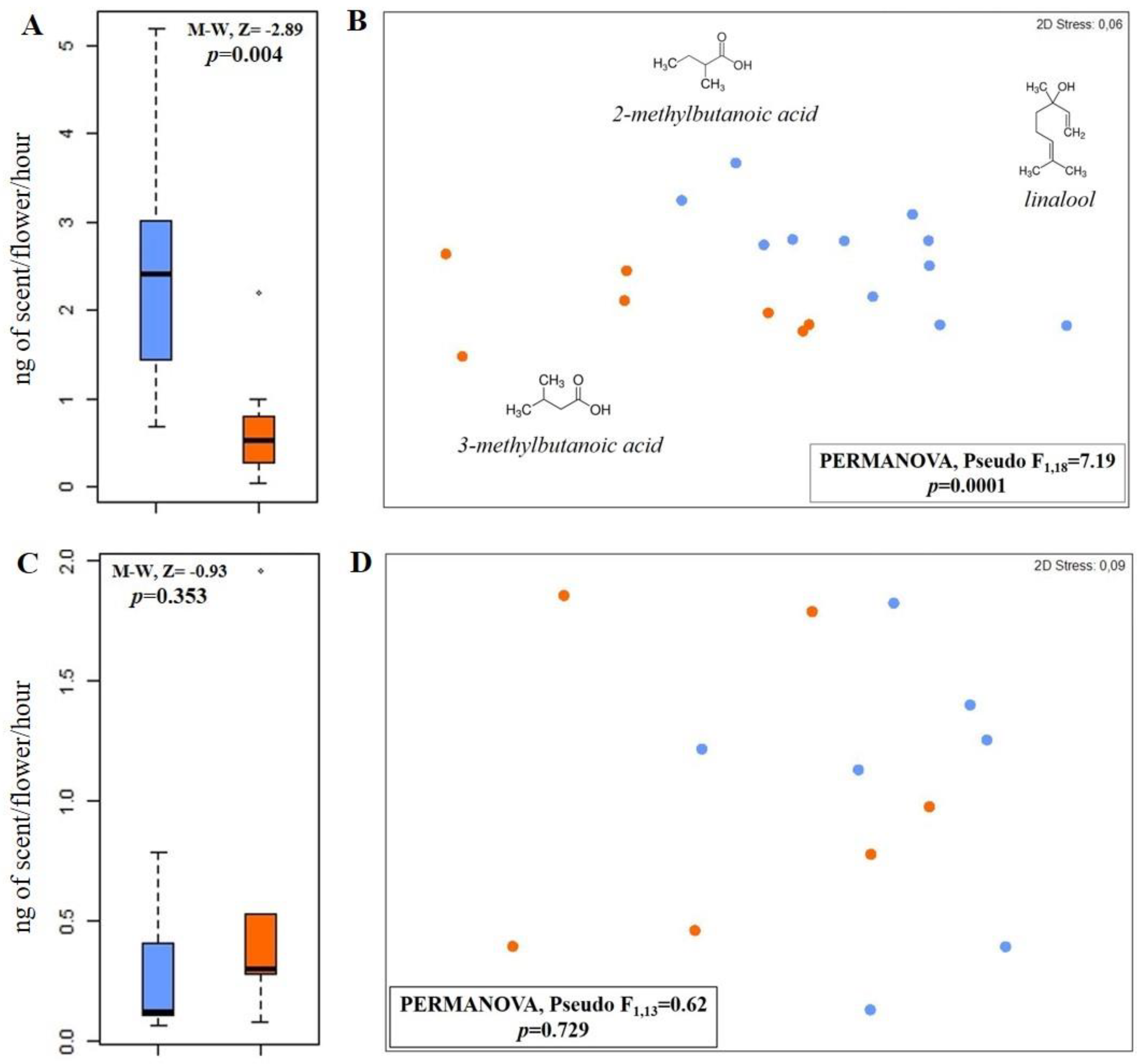
3. Results and Discussion
4. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- FAO-Food and Agriculture Organization. ‘Climate-Smart’ Agriculture, Policies, Practices and Finances for Food Security, Adaptation and Mitigation; FAO: Rome, Italy, 2010. [Google Scholar]
- United Nations. Open working group proposal for sustainable development goals. In Open Working Group of the General Assembly on Sustainable Development Goals; Full report A/68/970; United Nations: San Francisco, CA, USA, 2014. [Google Scholar]
- Ostberg, S.; Schewe, J.; Childers, K.; Frieler, K. Changes in crop yields and their variability at different levels of global warming. Earth Syst. Dyn. 2018, 9, 479–496. [Google Scholar] [CrossRef] [Green Version]
- Engel, M.S. Insect evolution. Curr. Biol. 2015, 25, R868–R872. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Eilers, E.J.; Kremen, C.; Greenleaf, S.S.; Garber, A.K.; Klein, A.M. Contribution of pollinator-mediated crops to nutrients in the human food supply. PLoS ONE 2011, 6, e21363. [Google Scholar] [CrossRef]
- Bailes, E.J.; Ollerton, J.; Pattrick, J.G.; Glover, B.J. How can an understanding of plant–pollinator interactions contribute to global food security? Curr. Opin. Plant Biol. 2015, 26, 72–79. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Potts, S.G.; Imperatriz-Fonseca, V.L.; Ngo, H.T.; Aizen, M.A.; Biesmeijer, J.C.; Breeze, T.D.; Dicks, L.V.; Garibaldi, L.A.; Hill, R.; Settele, J.; et al. Safeguarding pollinators and their values to human well-being. Nature 2016, 540, 220–229. [Google Scholar] [CrossRef] [Green Version]
- Klein, A.; Vaissiere, B.E.; Cane, J.H.; Steffan-Dewenter, I.; Cunningham, S.A.; Kremen, C.; Tscharntke, T. Importance of pollinators in changing landscapes for world crops. Proc. R. Soc. B Biol. Sci. 2007, 274, 303–313. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Giannini, T.C.; Cordeiro, G.D.; Freitas, B.M.; Saraiva, A.M.; Imperatriz-Fonseca, V.L. The dependence of crops for pollinators and the economic value of pollination in Brazil. J. Econ. Entomol. 2015, 108, 849–857. [Google Scholar] [CrossRef]
- Dötterl, S.; Vereecken, N. The chemical ecology and evolution of bee-flower interactions: A review and perspectives. Can. J. Zool. 2010, 88, 668–697. [Google Scholar] [CrossRef]
- Potts, S.G.; Biesmeijer, J.C.; Kremen, C.; Neumann, P.; Schweiger, O.; Kunin, W.E. Global pollinator declines: Trends, impacts and drivers. Trends Ecol. Evol. 2010, 25, 345–353. [Google Scholar] [CrossRef] [PubMed]
- El-Sayed, A.M.; Sporle, A.; Colhoun, K.; Furlong, J.; White, R.; Suckling, D.M. Scents in orchards: Floral volatiles of four stone fruit crops and their attractiveness to pollinators. Chemoecology 2018, 28, 39–49. [Google Scholar] [CrossRef]
- Rachersberger, M.; Cordeiro, G.D.; Schäffler, I.; Dötterl, S. Honeybee pollinators use visual and floral scent cues to find apple (Malus domestica) flowers. J. Agric. Food Chem. 2019, 67, 13221–13227. [Google Scholar] [CrossRef]
- Walters, J.; Zavalnitskaya, J.; Isaacs, R.; Szendrei, Z. Heat of the moment: Extreme heat poses a risk to bee-plant interactions and crop yields. Curr. Opin. Insect Sci. 2022, 52, 100927. [Google Scholar] [CrossRef]
- Dobson, H.E.M. Relationship between floral fragrance composition and type of pollinator. In Biology of Floral Scent; Dudareva, N., Pichersky, E., Eds.; CRC Press: Boca Raton, FL, USA, 2006; pp. 147–198. [Google Scholar]
- Knudsen, J.T.; Tollsten, L. Trends in floral scent chemistry in pollination syndromes—Floral scent composition in moth-pollinated taxa. Bot. J. Linn. Soc. 1993, 113, 263–284. [Google Scholar] [CrossRef]
- Dötterl, S.; Jürgens, A. Spatial fragrance patterns in flowers of Silene latifolia: Lilac compounds as olfactory nectar guides? Plant Syst. Evol. 2005, 255, 99–109. [Google Scholar] [CrossRef]
- Wright, G.A.; Choudhary, A.F.; Bentley, M.A. Reward quality influences the development of learned olfactory biases in honeybees. Proc. R. Soc. B Biol. Sci. 2009, 276, 2597–2604. [Google Scholar] [CrossRef] [Green Version]
- Knudsen, J.T.; Eriksson, R.; Gershenzon, J.; Ståhl, B. Diversity and distribution of floral scent. Bot. Rev. 2006, 72, 1. [Google Scholar] [CrossRef]
- Klatt, B.K.; Burmeister, C.; Westphal, C.; Tscharntke, T.; von Fragstein, M. Flower volatiles, crop varieties and bee responses. PLoS ONE 2013, 8, e72724. [Google Scholar] [CrossRef]
- Cordeiro, G.D.; Pinheiro, M.; Dötterl, S.; Alves-dos-Santos, I. Pollination of Campomanesia phaea (Myrtaceae) by night-active bees: A new nocturnal pollination system mediated by floral scent. Plant Biol. 2017, 19, 132–139. [Google Scholar] [CrossRef]
- Cordeiro, G.D.; Santos, I.G.F.; Silva, C.I.; Schlindwein, C.; Alves-dos-Santos, I.; Dötterl, S. Nocturnal floral scent profiles of Myrtaceae fruit crops. Phytochemistry 2019, 162, 193–198. [Google Scholar] [CrossRef] [PubMed]
- Krug, C.; Cordeiro, G.D.; Schäffler, I.; Silva, C.I.; Oliveira, R.; Schlindwein, C.; Dötterl, S.; Alves-dos-Santos, I. Nocturnal bee pollinators are attracted to Guarana flowers by their scents. Front. Plant Sci. 2018, 9, 1072. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lukas, K.; Harig, T.; Schulz, S.; Hadersdorfer, J.; Dötterl, S. Flowers of European pear release common and uncommon volatiles that can be detected by honey bee pollinators. Chemoecology 2019, 29, 211–223. [Google Scholar] [CrossRef] [Green Version]
- Jakobsen, H.B.; Olsen, C.E. Influence of climatic factors on emission of flower volatiles in situ. Planta 1994, 192, 365–371. [Google Scholar] [CrossRef]
- Hu, Z.; Zhang, H.; Leng, P.; Zhao, J.; Wang, W.; Wang, S. The emission of floral scent from Lilium ‘siberia’ in response to light intensity and temperature. Acta Physiol. Plant 2013, 35, 1691–1700. [Google Scholar] [CrossRef]
- Farré-Armengol, G.; Filella, I.; Llusià, J.; Niinemets, Ü.; Peñuelas, J. Changes in floral bouquets from compound-specific responses to increasing temperatures. Glob. Chang. Biol. 2014, 20, 3660–3669. [Google Scholar] [CrossRef] [Green Version]
- Borghi, M.; Perez de Souza, L.; Yoshida, T.; Fernie, A.R. Flowers and climate change: A metabolic perspective. New Phytol. 2019, 224, 1425–1441. [Google Scholar] [CrossRef] [Green Version]
- Barman, M.; Mitra, A. Floral maturation and changing air temperatures influence scent volatiles biosynthesis and emission in Jasminum auriculatum Vahl. Environ. Exp. Bot. 2021, 181, 104296. [Google Scholar] [CrossRef]
- Cordeiro, G.D.; Dötterl, S. Global warming impairs the chemical attractiveness to pollinators. Paris-Lodron University of Salzburg, Salzburg, Austria. 2022; submitted. [Google Scholar]
- Majetic, C.J.; Raguso, R.A.; Ashman, T.L. The sweet smell of success: Floral scent affects pollinator attraction and seed fitness in Hesperis matronalis. Funct. Ecol. 2009, 23, 480–487. [Google Scholar] [CrossRef]
- Arpaia, S.; De Cristofaro, A.; Guerrieri, E.; Bossi, S.; Cellini, F.; Di Leo, G.M.; Germinara, G.S.; Iodice, L.; Maffei, M.E.; Petrozza, A.; et al. Foraging activity of bumblebees (Bombus terrestris L.) on Bt-expressing eggplants. Arthropod Plant Interact. 2011, 5, 255–261. [Google Scholar] [CrossRef]
- Morse, A.; Kevan, P.; Shipp, L.; Khosla, S.; McGarvey, B. The impact of greenhouse tomato (Solanales: Solanaceae) floral volatiles on bumble bee (Hymenoptera: Apidae) pollination. Environ. Entomol. 2012, 41, 855–864. [Google Scholar] [CrossRef]
- Beyaert, I.; Hilker, M. Plant odour plumes as mediators of plant-insect interactions. Biol. Rev. 2014, 89, 68–81. [Google Scholar] [CrossRef] [PubMed]
- IPCC. Summary for Policymakers. In Climate Change 2021: The Physical Science Basis. Contribution of Working Group I to the Sixth Assessment Report of the Intergovernmental Panel on Climate Change; Masson-Delmotte, V., Zhai, P., Pirani, A., Connors, S.L., Péan, C., Berger, S., Caud, N., Chen, Y., Goldfarb, L., Gomis, M.I., et al., Eds.; Cambridge University Press: Cambridge, UK, 2021. [Google Scholar]
- Björkman, T. Role of honey bees (Hymenoptera: Apidae) in the pollination of buckwheat in Eastern North America. J. Econ. Entomol. 1995, 88, 1739–1745. [Google Scholar] [CrossRef] [Green Version]
- Sasaki, H.; Wagatsuma, T. Bumblebees (Apidae: Hymenoptera) are the main pollinators of common buckwheat, Fagopyrum esculentum, in Hokkaido, Japan. Appl. Entomol. Zool. 2007, 42, 659–661. [Google Scholar] [CrossRef] [Green Version]
- Adegas, J.E.B.; Nogueira-Couto, R.H. Entomophilous pollination in rape (Brassica napus L. var. oleifera) in Brazil. Apidologie 1992, 23, 203–209. [Google Scholar] [CrossRef] [Green Version]
- Woodcock, B.A.; Edwards, M.; Redheada, J.; Meek, W.R.; Nuttall, P.; Falk, S.; Nowakowski, M.; Pywell, R.F. Crop flower visitation by honeybees, bumblebees and solitary bees: Behavioural differences and diversity responses to landscape. Agric. Ecosyst. Environ. 2013, 171, 1–8. [Google Scholar] [CrossRef] [Green Version]
- Ouvrard, P.; Quinet, M.; Jacquemart, A.L. Breeding system and pollination biology of Belgian oilseed rape cultivars (Brassica napus. Crop Sci. 2017, 57, 1455–1463. [Google Scholar] [CrossRef]
- Burkle, L.A.; Runyon, J.B. Drought and leaf herbivory influence floral volatiles and pollinator attraction. Glob. Chang. Biol. 2016, 22, 1644–1654. [Google Scholar] [CrossRef]
- Chandler, J.; Corbesier, L.; Spielmann, P.; Dettendorfer, J.; Stahl, D.; Apel, K.; Melzer, S. Modulating flowering time and prevention of pod shatter in oilseed rape. Mol. Breed. 2005, 15, 87–94. [Google Scholar] [CrossRef]
- Hara, T.; Ohsawa, R. Accurate evaluation of photoperiodic sensitivity and genetic diversity in common Buckwheat under a controlled environment. Plant Prod. Sci. 2013, 16, 247–254. [Google Scholar] [CrossRef]
- Björkman, T.; Bellinder, R.; Hahn, R.; Shail, J.W., Jr. Buckwheat Cover Crop Handbook; Cornell University Cooperative Extension: Geneva, NY, USA, 2008. [Google Scholar]
- Thomas, P. Canola Grower’s Manual; Canola Council of Canada: Winnipeg, MB, Canada, 2003. [Google Scholar]
- PlavcovÁ, E.; KyselÝ, J. Evaluation of daily temperatures in Central Europe and their links to large-scale circulation in an ensemble of regional climate models. Tellus A Dyn. 2011, 63, 763–781. [Google Scholar] [CrossRef] [Green Version]
- Worldclim. New 1-km Spatial Resolution Climate Surfaces for Global Land Areas. 2017. Available online: http://worldclim.org/version2 (accessed on 6 April 2019).
- Dötterl, S.; Wolfe, L.M.; Jürgens, A. Qualitative and quantitative analyses of flower scent in Silene latifolia. Phytochemistry 2005, 66, 203–213. [Google Scholar] [CrossRef]
- Mitchell, T.C.; Dötterl, S.; Schaefer, H. Hawkmoth pollination and elaborate petals in Cucurbitaceae: The case of the Caribbean endemic Linnaeosicyos amara. Flora 2015, 216, 50–56. [Google Scholar] [CrossRef]
- Adams, R.P. Identification of Essential Oil Components by Gas Chromatography/Mass Spectrometry; Allured Publishing Corporation: Carol Stream, IL, USA, 2007; Volume 456, 811p. [Google Scholar]
- Dötterl, S.; Füssel, U.; Jürgens, A.; Aas, G. 1,4-Dimethoxybenzene, a floral scent compound in willows that attracts an oligolectic bee. J. Chem. Ecol. 2005, 31, 2993–2998. [Google Scholar] [CrossRef] [PubMed]
- Heiduk, A.; Brake, I.; von Tschirnhaus, M.; Göhl, M.; Jürgens, A.; Johnson, S.D.; Meve, U.; Dötterl, S. Ceropegia sandersonii mimics attacked honey bees to attract kleptoparasitic flies for pollination. Curr. Biol. 2016, 26, 2787–2793. [Google Scholar] [CrossRef]
- Hammer, Ø.; Harper, D.A.T.; Ryan, P.D. Past: Paleontological tatistics software package for education and data analysis. Palaeontol. Electron. 2001, 4, 9. [Google Scholar]
- Clarke, K.R.; Gorley, R.N. Primer v6: User Manual/Tutorial; Primer-E Ltd.: Plymouth, UK, 2006; pp. 1–91. [Google Scholar]
- Foti, M.C.; Rostás, M.; Peri, E.; Park, K.C.; Slimani, T.; Wratten, S.D.; Colazza, S. Chemical ecology meets conservation biological control: Identifying plant volatiles as predictors of floral resource suitability for an egg parasitoid of stink bugs. J. Pest Sci. 2017, 90, 299–310. [Google Scholar] [CrossRef]
- Henning, J.A.; Teuber, L.R. Combined gas chromatography–electroantennogram characterization of alfalfa floral volatiles recognized by honey bees (Hymenoptera: Apidae). J. Econ. Entomol. 1992, 85, 226–232. [Google Scholar] [CrossRef]
- Knauer, A.C.; Schiestl, F.P. Bees use honest floral signals as indicators of reward when visiting flowers. Ecol. Lett. 2014, 18, 135–143. [Google Scholar] [CrossRef]
- Twidle, A.M.; Barker, D.; Seal, A.G.; Fedrizzi, B.; Suckling, D.M. Identification of floral volatiles and pollinator responses in kiwifruit cultivars, Actinidia chinensis var. chinensis. J. Chem. Ecol. 2018, 44, 406–415. [Google Scholar] [CrossRef]
- Liu, J.; Zhang, J.; Shen, J.; Zhao, H.; Ma, W.; Jiang, Y. Differences in EAG response and behavioral choices between honey bee and bumble bee to tomato flower volatiles. Insects 2022, 13, 987. [Google Scholar] [CrossRef]
- Evans, K.A.; Allen-Williams, L.J. Electroantennogram responses of the cabbage seed weevil, Ceutorhynchus assimilis, to oilseed rape, Brassica napus ssp. oleifera, volatiles. J. Chem. Ecol. 1992, 18, 1641–1659. [Google Scholar] [CrossRef]
- Robertson, G.W.; Griffiths, D.W.; Smith, W.M.; Butcher, R.D. The application of thermal desorption-gas chromatography-mass spectrometry to the analyses of flower volatiles from five varieties of oilseed rape (Brassica napus spp. oleifera). Phytochem. Anal. 1993, 4, 152–157. [Google Scholar] [CrossRef]
- Jakobsen, H.B.; Friis, P.; Nielsen, J.K.; Olsen, C.E. Emission of volatiles from flowers and leaves of Brassica napus in situ. Phytochemistry 1994, 37, 695–699. [Google Scholar] [CrossRef]
- Blight, M.M.; Pickett, J.A.; Wadhams, L.J.; Woodcock, C.M. Antennal perception of oilseed rape Brassica napus (Brassicaceae) volatiles by the cabbage seed weevil, Ceutorhynchus assimilis (Coleoptera: Curculionidae). J. Chem. Ecol. 1995, 21, 1649–1664. [Google Scholar] [CrossRef]
- Kobayashi, K.; Arai, M.; Tanaka, A.; Matsuyama, S.; Honda, H. Variation in floral scent compounds recognized by honeybees in Brassicaceae crop species. Breed. Sci. 2012, 62, 293–302. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Blight, M.M.; Métayer, M.L.; Delègue, M.H.P.; Pickett, J.A.; Marion-Poll, F.; Wadhams, L.J. Identification of floral volatiles involved in recognition of oilseed rape flowers, Brassica napus by honeybees, Apis mellifera. J. Chem. Ecol. 1997, 23, 1715–1727. [Google Scholar] [CrossRef]
- Pham-Delègue, M.H.; Blight, M.M.; Kerguelen, V.; Le Métayer, M.; Marion-Poll, F.; Sandoz, J.C.; Wadhams, L.J. Discrimination of oilseed rape volatiles by the honeybee: Combined chemical and biological approaches. Entomol. Exp. Appl. 1997, 83, 87–92. [Google Scholar] [CrossRef] [Green Version]
- Bartlet, E.; Blight, M.M.; Lane, P.; Williams, I.H. The responses of the cabbage seed weevil Ceutorhynchus assimilis to volatile compounds from oilseed rape in a linear track olfactometer. Entomol. Exp. Appl. 1997, 85, 257–262. [Google Scholar] [CrossRef]
- Saunier, A.; Blande, J.D. The effect of elevated ozone on floral chemistry of Brassicaceae species. Environ. Pollut. 2019, 255, 113257. [Google Scholar] [CrossRef] [PubMed]
- Younas, M.; Ali, M.; Matloob, A.; Gul, H.T.; Saeed, S. Effect of drought stress on the foraging behavior of insect pollinators and the reproductive success of canola (Brassica napus L.). Emir. J. Food Agric. 2022, 34, 170–175. [Google Scholar] [CrossRef]
- Sagae, M.; Oyama-Okubo, N.; Ando, T.; Marchesi, E.; Nakayama, M. Effect of temperature on the floral scent emission and endogenous volatile profile of Petunia axillaris. Biosci. Biotechnol. Biochem. 2008, 72, 110–115. [Google Scholar] [CrossRef] [Green Version]
- Rering, C.C.; Franco, J.G.; Yeater, K.M.; Mallinger, R.E. Drought stress alters floral volatiles and reduces floral rewards, pollinator activity, and seed set in a global plant. Ecosphere 2020, 11, e03254. [Google Scholar] [CrossRef]
- Blois, J.L.; Zarnetske, P.L.; Fitzpatrick, M.C.; Finnegan, S. Climate change and the past, present, and future of biotic interactions. Science 2013, 341, 499–504. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Giannini, T.C.; Costa, W.F.; Cordeiro, G.D.; Imperatriz-Fonseca, V.L.; Saraiva, A.M.; Biesmeijer, J.; Garibaldi, L.A. Projected climate change threatens pollinators and crop production in Brazil. PLoS ONE 2017, 12, e0182274. [Google Scholar] [CrossRef] [PubMed] [Green Version]



{kind=link}
{kind=link}
| Buckwheat | Oilseed Rape | ||||
|---|---|---|---|---|---|
| Total Absolute Amount of Scent | Optimum | Warmer | Optimum | Warmer | |
| Mean ± SE | Mean ± SE | Mean ± SE | Mean ± SE | ||
| 2.38 ± 0.39 | 0.71 ± 0.28 | 0.29 ± 0.11 | 0.57 ± 0.28 | ||
| Compounds | KRI | ||||
| Aliphatics | |||||
| 2-methylpropanoic acid * | 742 | 0.8 (±0.2) | 1.1 (±1.1) | ||
| butanoic acid * | 769 | 5.1 (±0.9) | 2.6 (±1.3) | ||
| pentanoic acid * | 872 | 2.2 (±0.6) | |||
| hexyl isobutyrate * | 1146 | 0.1 (±0.1) | |||
| Aromatics | |||||
| p-benzoquinone * | 920 | 6.8 (±1.1) | 3.4 (±2.1) | ||
| p-cresol * | 1073 | 7.8 (±3.7) | 3.2 (±0.2) | ||
| 2-methoxyphenol * | 1095 | 0.1 (±0.1) | |||
| 1,4-dimethoxybenzene * | 1168 | 5.4 (±5.4) | 6.1 (±0.5) | ||
| p-anisaldehyde * | 1265 | 4.3 (±1.3) | 1.3 (±1.3) | 40.8 (±13.0) | 26.2 (±9.1) |
| p-hydroquinone * | 1267 | 0.2 (±0.1) | |||
| C5-branched chain compounds | |||||
| 3-methylbutanoic acid * | 833 | 28.0 (±2.8) | 45.4 (±2.5) | ||
| 2-methylbutanoic acid * | 850 | 18.0 (±1.9) | 27.3 (±4.9) | ||
| Monoterpenes | |||||
| sabinene * | 980 | 1.4 (±0.8) | 10.6 (±7.3) | ||
| δ-3-carene * | 1016 | 14.4 (±8.7) | 11.2 (±4.8) | ||
| β-phellandrene * | 1037 | 0.1 (±0.1) | |||
| (Z)-β-ocimene * | 1038 | 3.4 (±3.4) | |||
| (E)-β-ocimene * | 1050 | 3.5 (±1.3) | 3.9 (±3.9) | 22.3 (±7.1) | 13.0 (±8.4) |
| (Z)-linalool oxide furanoid * | 1078 | 0.6 (±0.2) | 0.5 (±0.5) | ||
| (E)-linalool oxide furanoid * | 1094 | 0.1 (±0.1) | |||
| linalool * | 1102 | 10.3 (±2.8) | 17.1 (±7.9) | 37.4 (±18.1) | |
| allo-ocimene * | 1130 | 1.6 (±1.6) | |||
| 1,3,8-p-menthatriene | 1136 | 0.3 (±0.2) | |||
| neo-allo-ocimene * | 1143 | 1.0 (±1.0) | |||
| phenylacetonitrile * | 1145 | 1.1 (±0.4) | |||
| terpinene-4-ol * | 1187 | 1.5 (±0.7) | |||
| Nitrogen-containing compounds | |||||
| 3-pyridinecarboxaldehyde * | 1002 | 0.1 (±0.1) | |||
| 2-methylpyrazine * | 1078 | 0.4 (±0.2) | |||
| Indole * | 1305 | 2.9 (±0.8) | |||
| Irregular terpenes | |||||
| (E)-4,8-dimethyl-1,3,7-nonatriene | 1119 | 4.4 (±1.1) | 0.4 (±0.4) | ||
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Cordeiro, G.D.; Dötterl, S. Floral Scents in Bee-Pollinated Buckwheat and Oilseed Rape under a Global Warming Scenario. Insects 2023, 14, 242. https://doi.org/10.3390/insects14030242
Cordeiro GD, Dötterl S. Floral Scents in Bee-Pollinated Buckwheat and Oilseed Rape under a Global Warming Scenario. Insects. 2023; 14(3):242. https://doi.org/10.3390/insects14030242
Chicago/Turabian StyleCordeiro, Guaraci Duran, and Stefan Dötterl. 2023. "Floral Scents in Bee-Pollinated Buckwheat and Oilseed Rape under a Global Warming Scenario" Insects 14, no. 3: 242. https://doi.org/10.3390/insects14030242