1. Introduction
Invasive insects pose a serious threat to forest health, ecosystem stability, and even public health in areas where they establish [
1,
2,
3,
4]. Field observations of an exotic insect’s life cycle can be important to understanding their population dynamics in the new environment and when control efforts could be maximised [
5,
6]. This would be particularly important for understanding species establishing in both the northern and southern hemispheres, due to the inverse of calendar season. Phenological studies might include documenting insect activity patterns and emergence of developmental stages (e.g., [
7,
8]) and, in the case of herbivorous insects, matching these to fluctuations in host quality [
9,
10]. For phloem feeding insects, mapping the timing of feeding life stages and feeding activity can be used to predict when impacts are greatest [
11,
12,
13]. In Australia, insect forest pests are being detected at an accelerating rate [
14,
15], many of which are phloem-feeding insects.
In 2014, the giant pine scale,
Marchalina hellenica Gennadius, (Hemiptera, Marchalinidae) was first detected in Victoria, Australia, feeding on the novel host
Pinus radiata, or California Monterey pine [
16].
Marchalina hellenica is endemic to the eastern Mediterranean basin [
17,
18,
19]; its establishment on
P. radiata (which originates from North America) poses a significant threat to Australian softwood plantations.
Pinus radiata represents the vast majority of Australia’s more than 1 million hectares of softwood plantation [
20]. Despite the widespread use of
P. radiata as a plantation species and an ornamental, in public gardens and farm windrows, across Australia, the documented distribution of
M. hellenica is currently restricted to southeast Melbourne [
16].
Little is known about how the life cycle of
M. hellenica in its new Australian habitat compares to its endemic range. The insect’s three immature nymphal instars acquire nutrients for development from the sap of pine trees [
21]. The nymphal instars feed on phloem by inserting syringe-like stylets into twigs, branches, trunks, and exposed roots [
22,
23,
24]. As with many phloem-feeding insects, excess carbohydrates and undesirable compounds such as insecticides [
25] that are ingested must be excreted as honeydew [
26,
27].
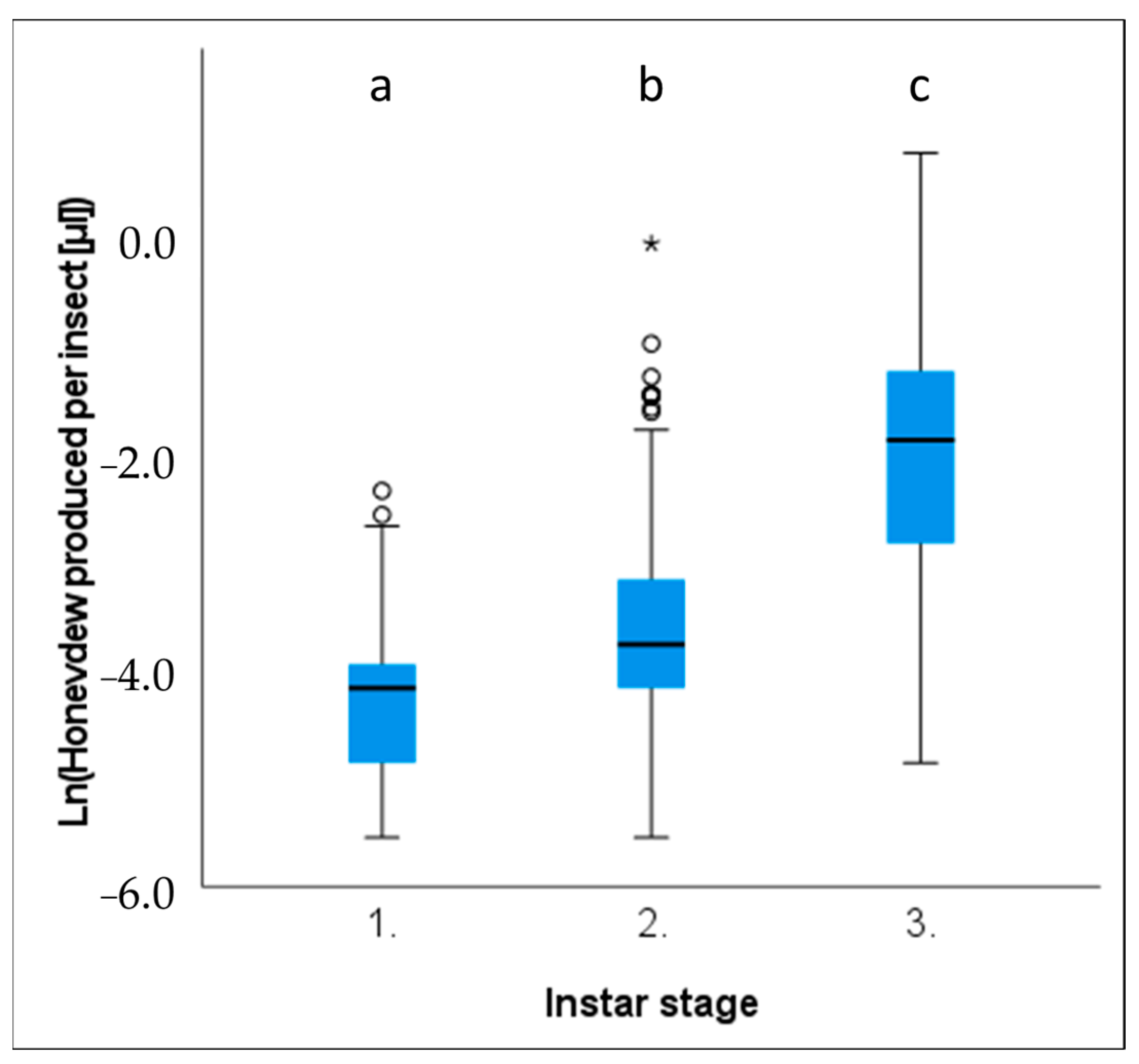
Marchalina hellenica does not feed on host trees all year round, and only the nymphal instar stages possess a functioning stylet and produce honeydew [
28].
Marchalina hellenica likely originates from Mount Carmel in Northern Israel, the origin of its primary host,
P. halepensis [
29]. Honeydew produced by
M. hellenica is collected by honeybees across the Eastern Mediterranean and accounts for 60% (~15,000 tonnes) of annual honey produced in Greece [
30,
31]. While this is important for apiculture in Greece, it also highlights the phloem feeding capability of this insect. Its impact as an introduced pest to Australian forestry could be severe, particularly if feeding stages are prolonged and/or populations flourish in the absence of important native predators, such as the predatory fly,
Neoleucopis kartliana Tanasijtshuk (Diptera: Chamaemyiidae) [
32]. In addition to exuding honeydew,
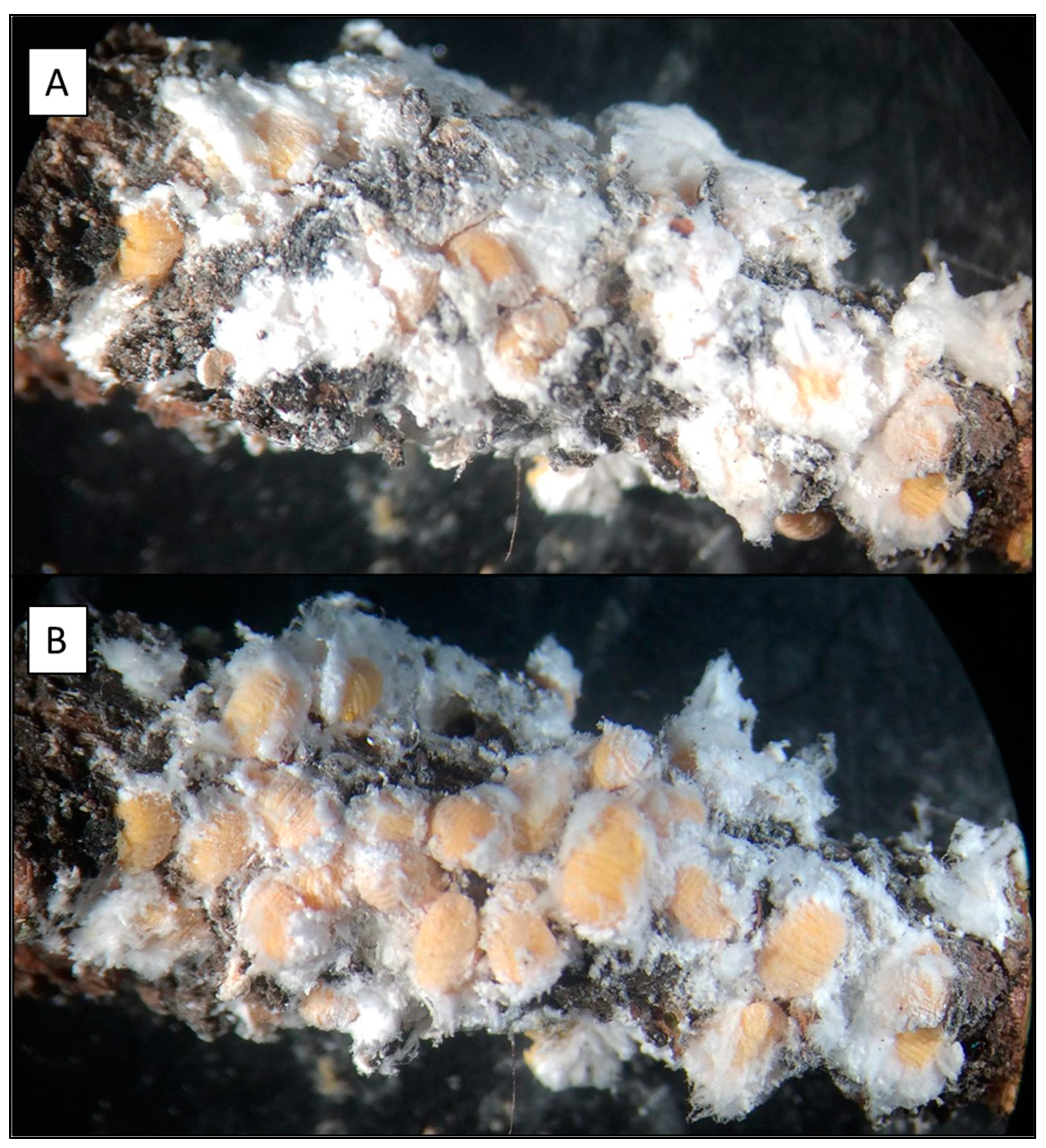
M. hellenica secretes cotton-like wax filaments [
21] termed flocculent, which may provide a hydrophobic layer and microclimate to protect scales from desiccation and flooding of feeding sites [
33,
34,
35].
Insect density and availability of honeydew are key indicators of the resources expropriated by
M. hellenica. Explaining variation in these indicators is important for understanding the insect’s activity patterns. As an ectothermic phloem feeder,
M. hellenica activity is likely related to climate via effects on host nutritional quality [
36,
37]. As an insect sheltering under bark and flocculent, climate conditions directly experienced by
M. hellenica may therefore provide inferior explanations for variation in insect activity than the climate experienced by host plants [
9,
38]. Understanding how
M. hellenica responds to the climate may help predict the potential spread of this pest in Australia or the impact as the climate in the Mediterranean [
39,
40] and Pacific region [
41,
42] trend towards more unpredictable conditions through climate change.
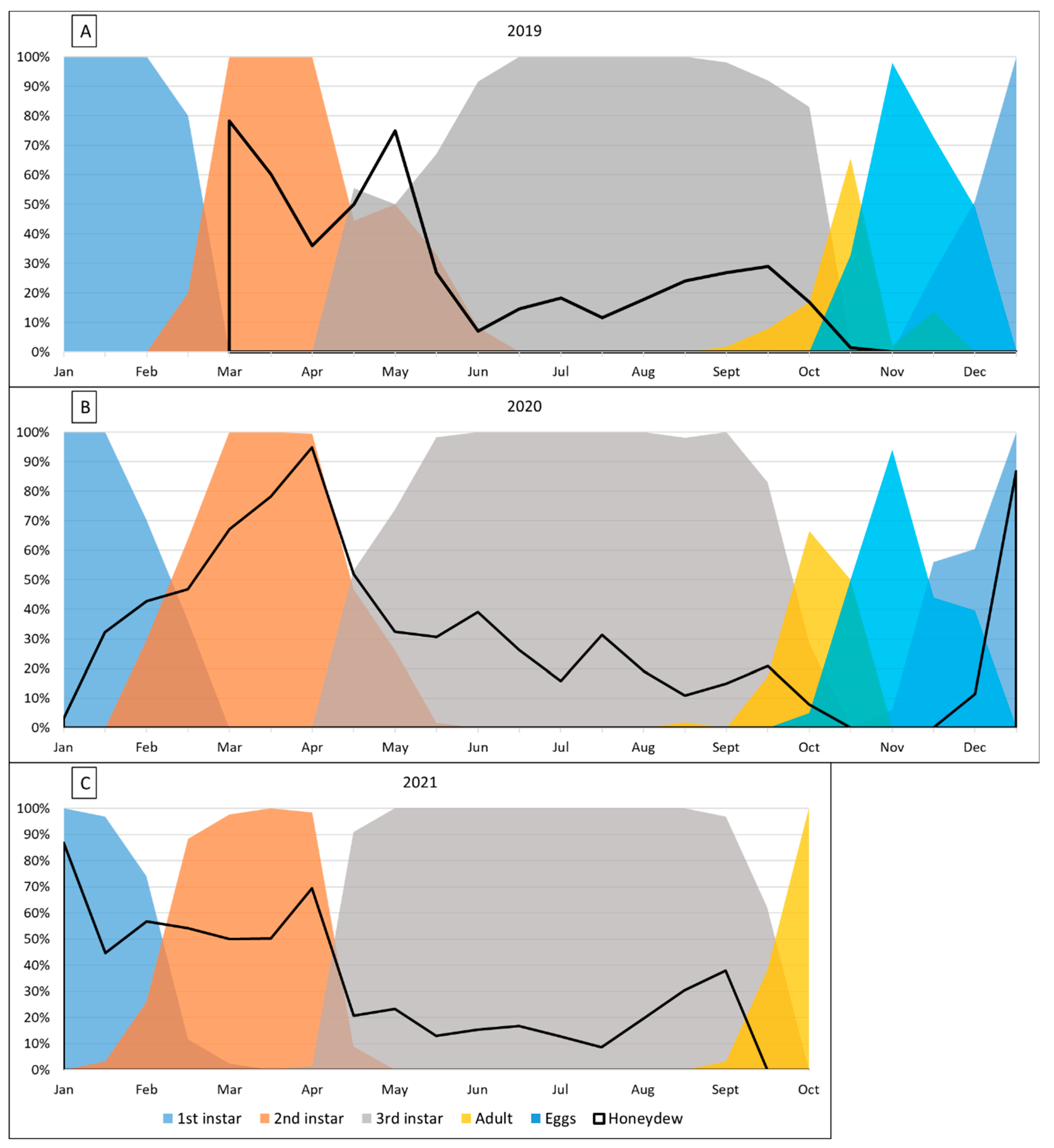
In this study, we used field-sampling to document the seasonal phenology of M. hellenica in southeast Melbourne, Victoria, across two and a half insect generations. As these represent the only known M. hellenica populations in the southern hemisphere or outside Europe, we explored whether major life cycle milestones (e.g., nymph emergence or moults) in Melbourne aligned with equivalent seasonal periods to East-Mediterranean conspecifics (i.e., 6 calendar-month difference). We also investigated whether the timing of instar stages and honeydew production (as the proportion of honeydew producing insects, HPI) followed the same seasonal pattern as Mediterranean conspecifics. As a univoltine (one generation per year) and semelparous (mortality following oviposition) insect, density of M. hellenica was predicted to reach the generational minimum during the adult stage and maximum at nymph emergence. Finally, we explored the prediction that variation in M. hellenica activity would be best explained by the environmental conditions experienced by the host tree, rather than the conditions directly experienced by M. hellenica.
4. Discussion
Broad similarities were identified between Mediterranean and Australian
M. hellenica populations in the timing of life stages in local seasons [
31,
53]. As expected, major developmental milestones typically exhibited a six-month delay between Mediterranean and Australian populations. In Australia, the occurrence of life stages between years of study was broadly similar, although the developmental stages showed progressive changes each year; for example, instar stages were detected earlier with successive generations. By contrast, in Greece [
50,
58], the timing of the occurrence of different instars varied between generations; however, initial detection of each life stage did not occur earlier or later with successive generations. Phenology and development can be delayed or accelerated by changes in climate, a trend observed in other scale insects [
10], other Hemiptera [
12], and other insect orders [
7,
53]. Given the relatively short time it has been present, Australian
M. hellenica may still be adjusting to the effects of climate drivers on their activity. The current study was limited by the fragmented distribution of exotic
P. radiata in Australia. Relating the seasonal phenology of
M. hellenica in other climates to that in Australia would provide valuable attestation of the relationships identified here.
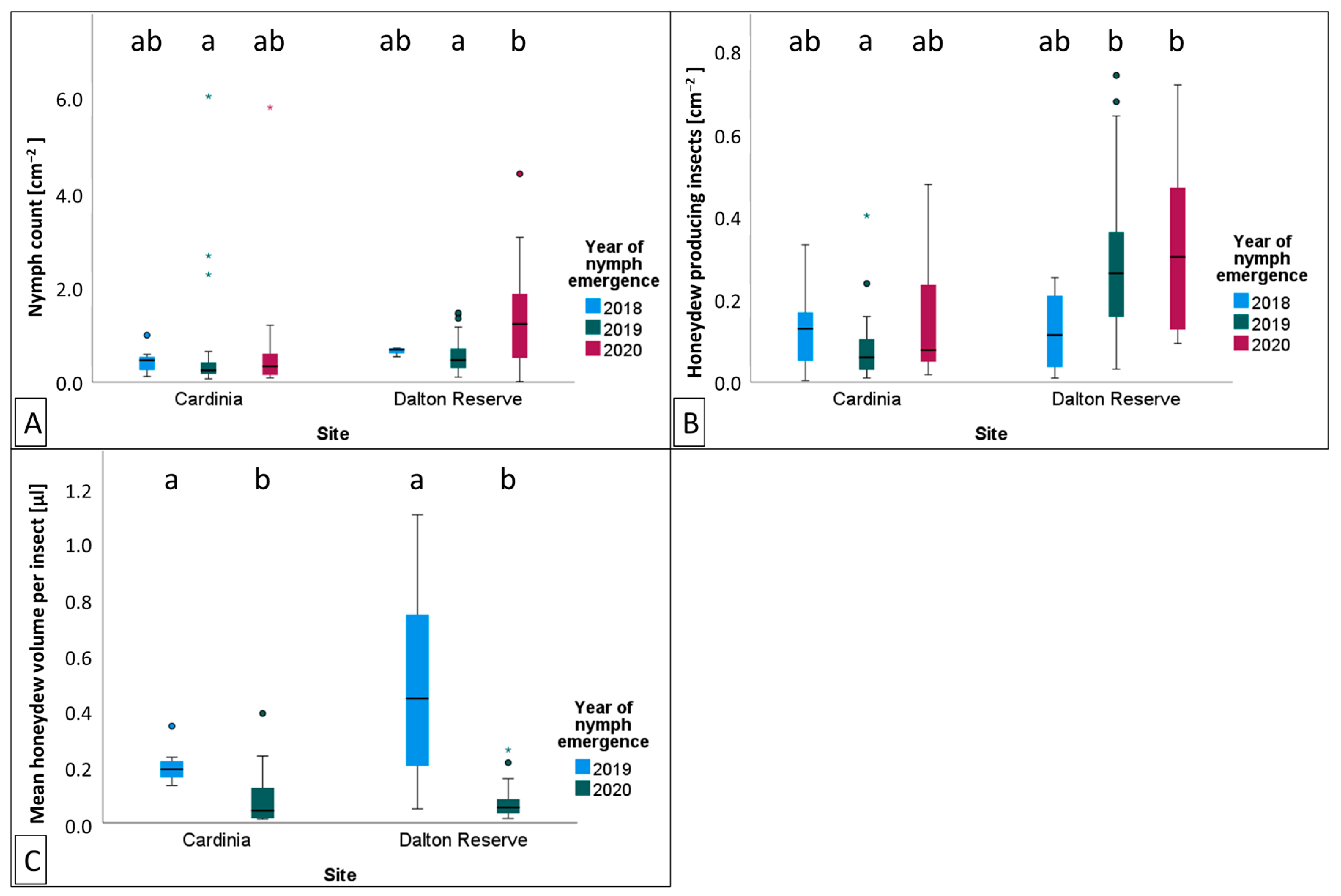
The densities of
M. hellenica on Australian
P. radiata were typically higher than Mediterranean conspecifics feeding on
Pinus halepensis during seasonally equivalent months [
51]; however, the proportion of honeydew producing insects did not reach proportions as high as those previously documented in Greece [
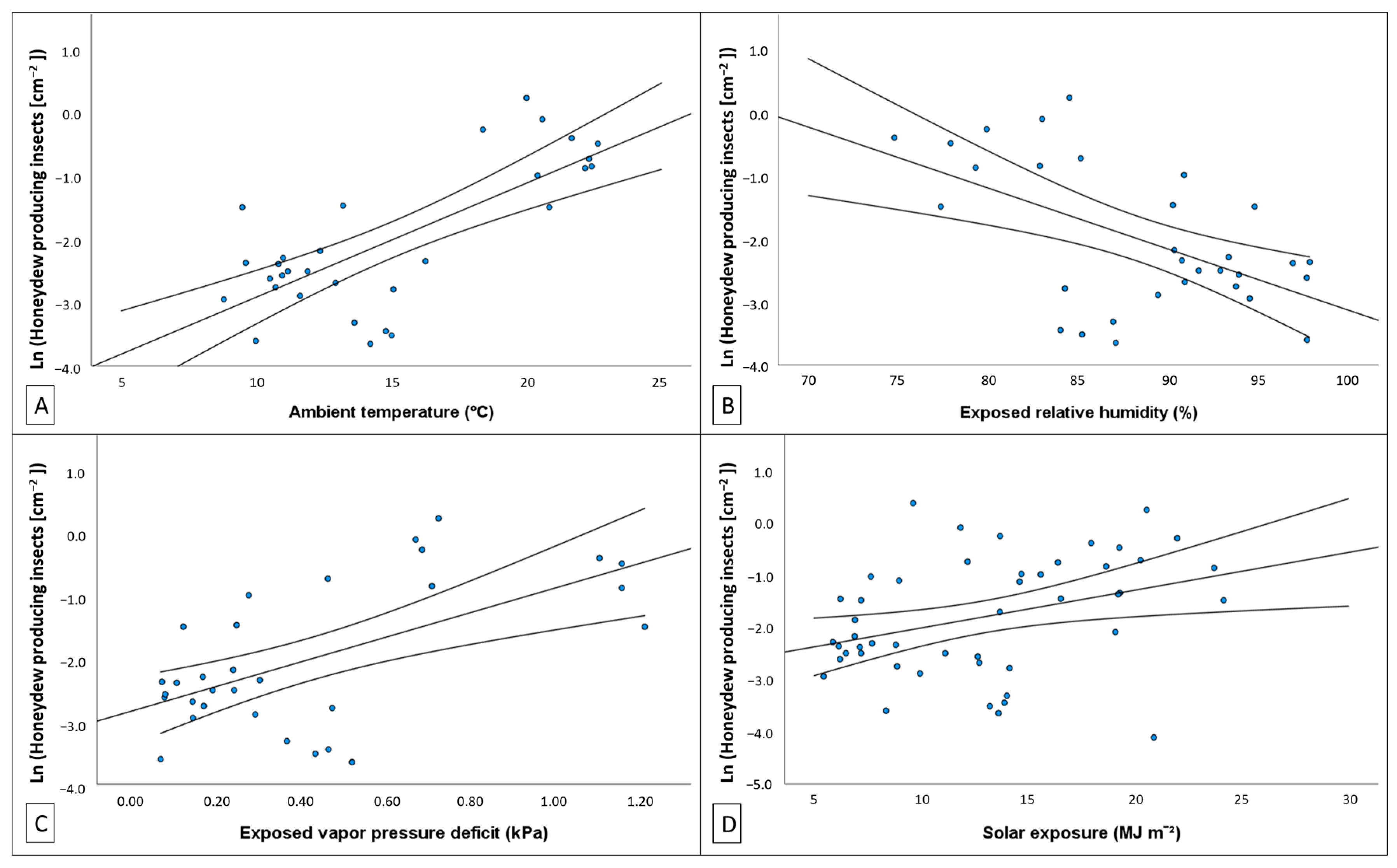
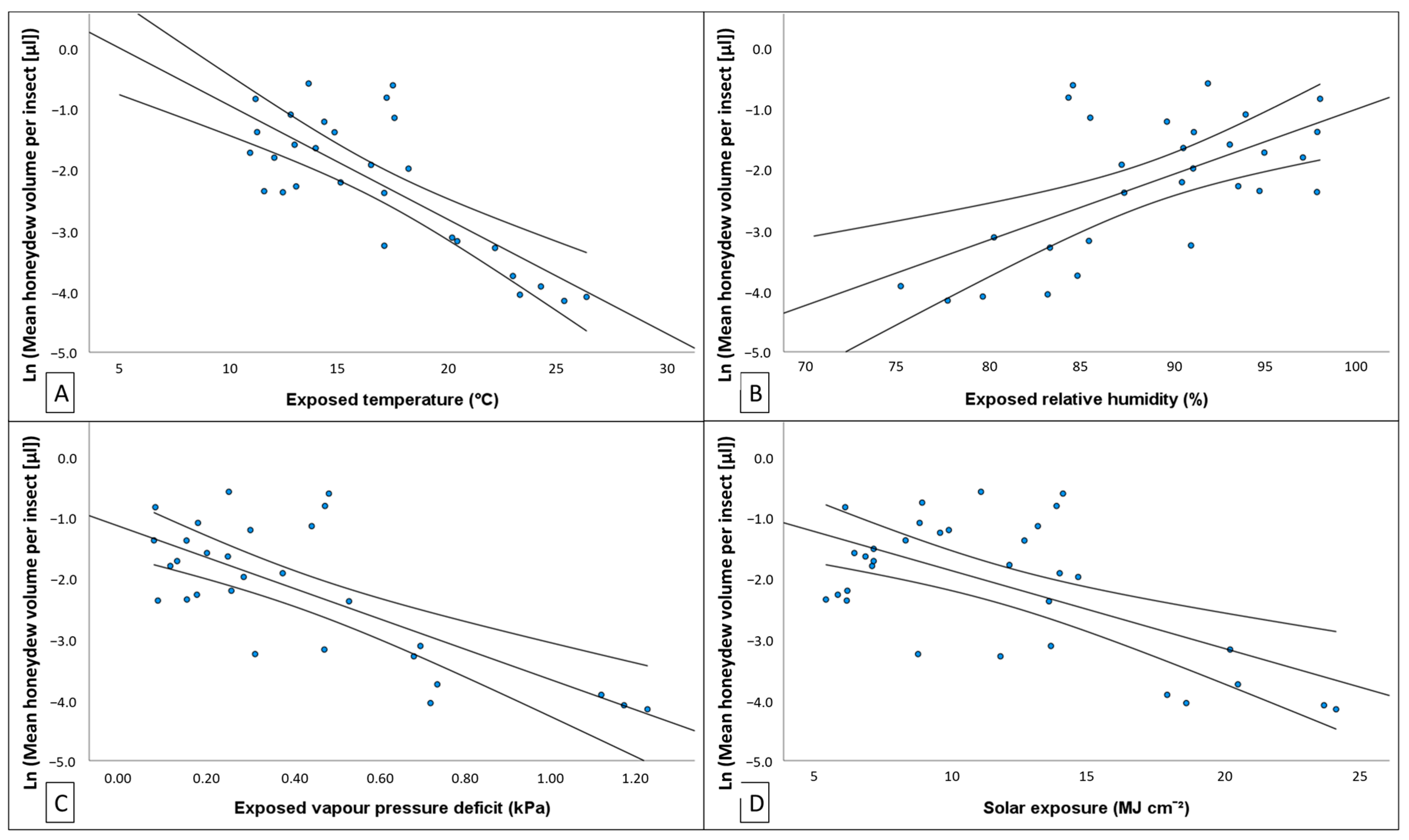
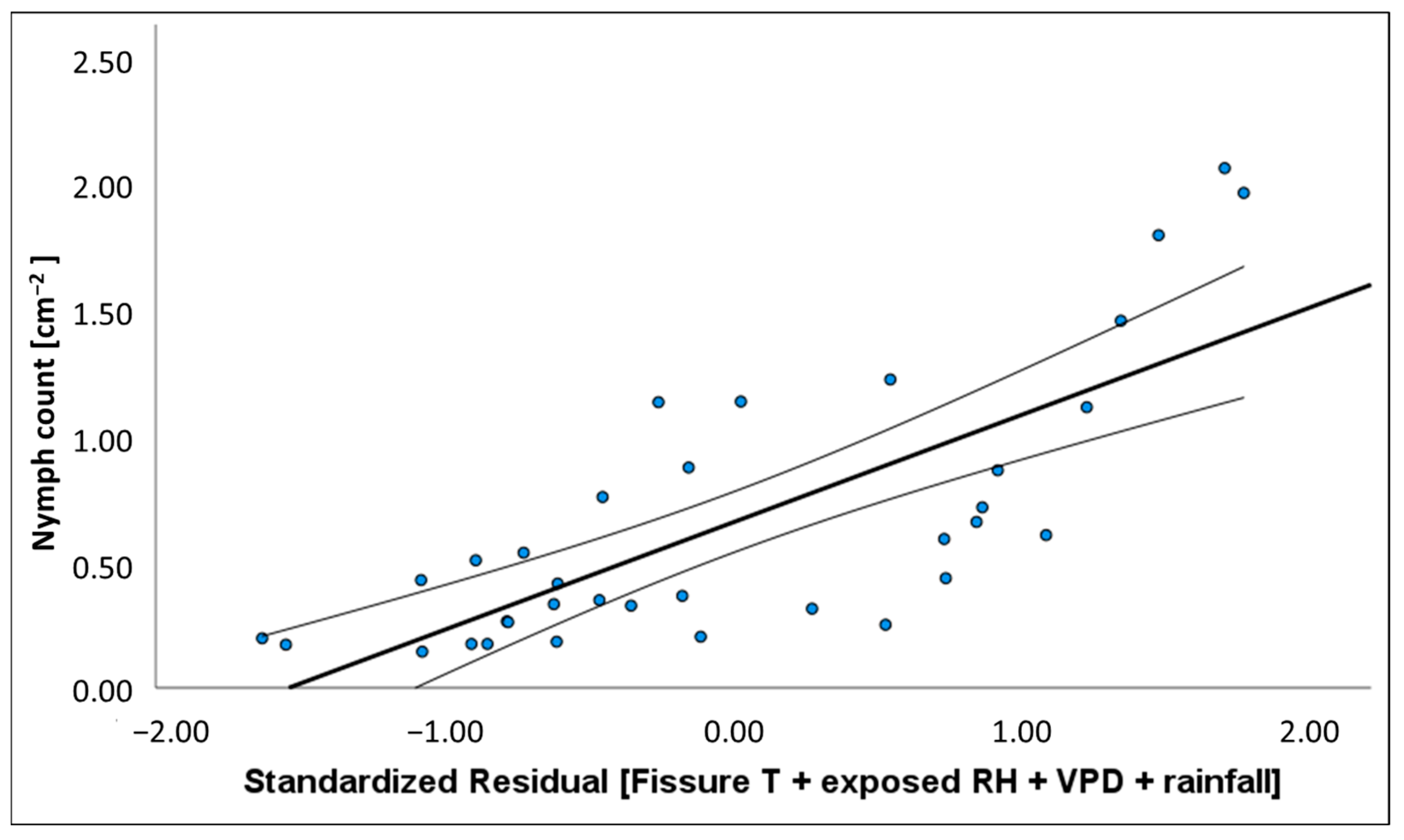
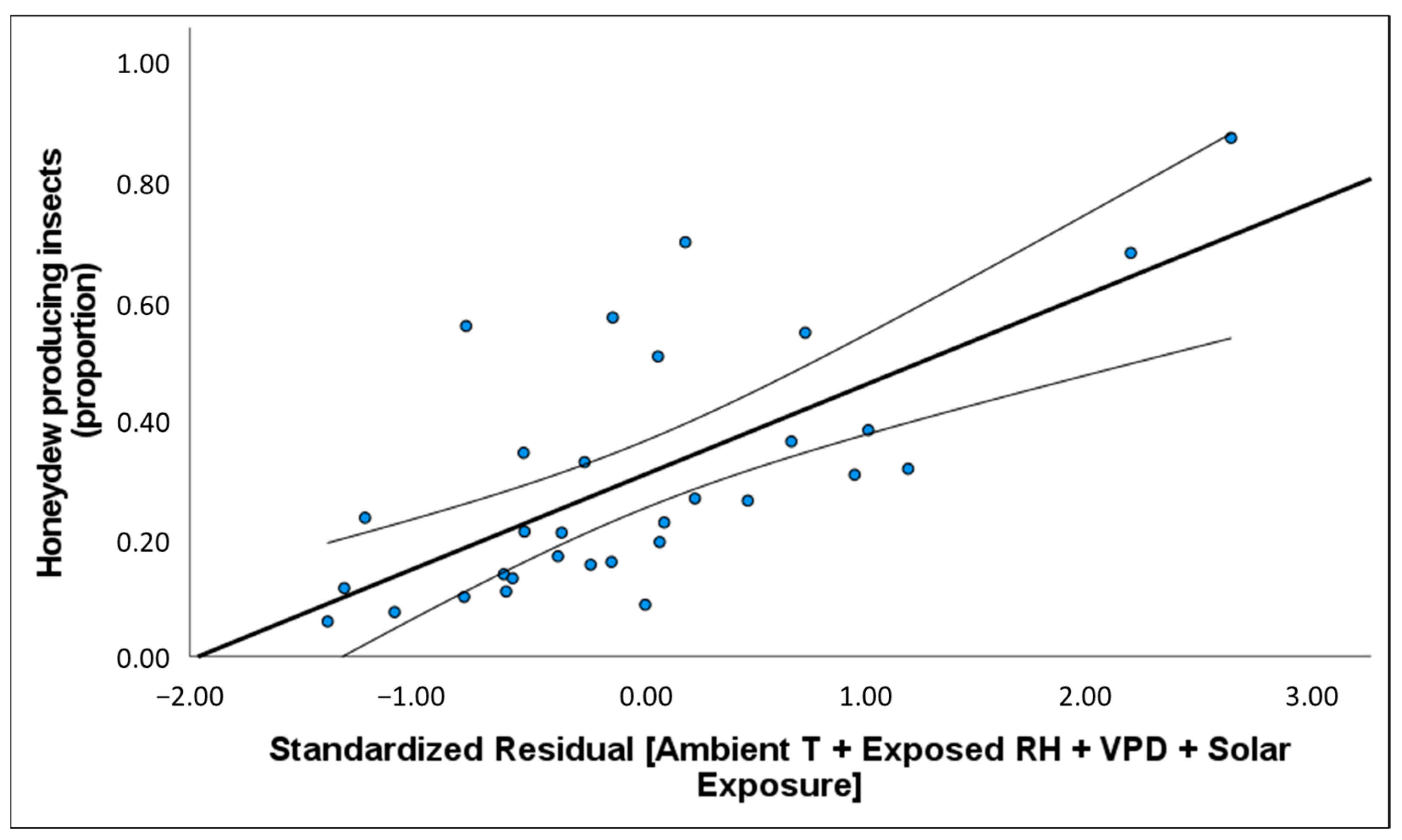
50]. We found evidence that variation in the density of honeydew producing insects and the quantity of honeydew produced could be explained by changes in temperature, humidity, vapour pressure in the atmosphere, and solar exposure. Depending on where climate was recorded (i.e., weather station, exposed tree trunk or bark fissure), the degree of association
M. hellenica demonstrated with climate variables varied greatly in this study. The insect’s flocculent and preference for crevices and non-sun facing aspects [
59] realises a microclimate distinct from broader forest conditions. Microhabitat climate was found to provide weaker predictions of insect activity, as it directly records the climate experienced by sheltered ectothermic
M. hellenica [
37], rather than the experience of the host. With regards to this study, the climate, as experienced by the host, most often provided the best predictive models.
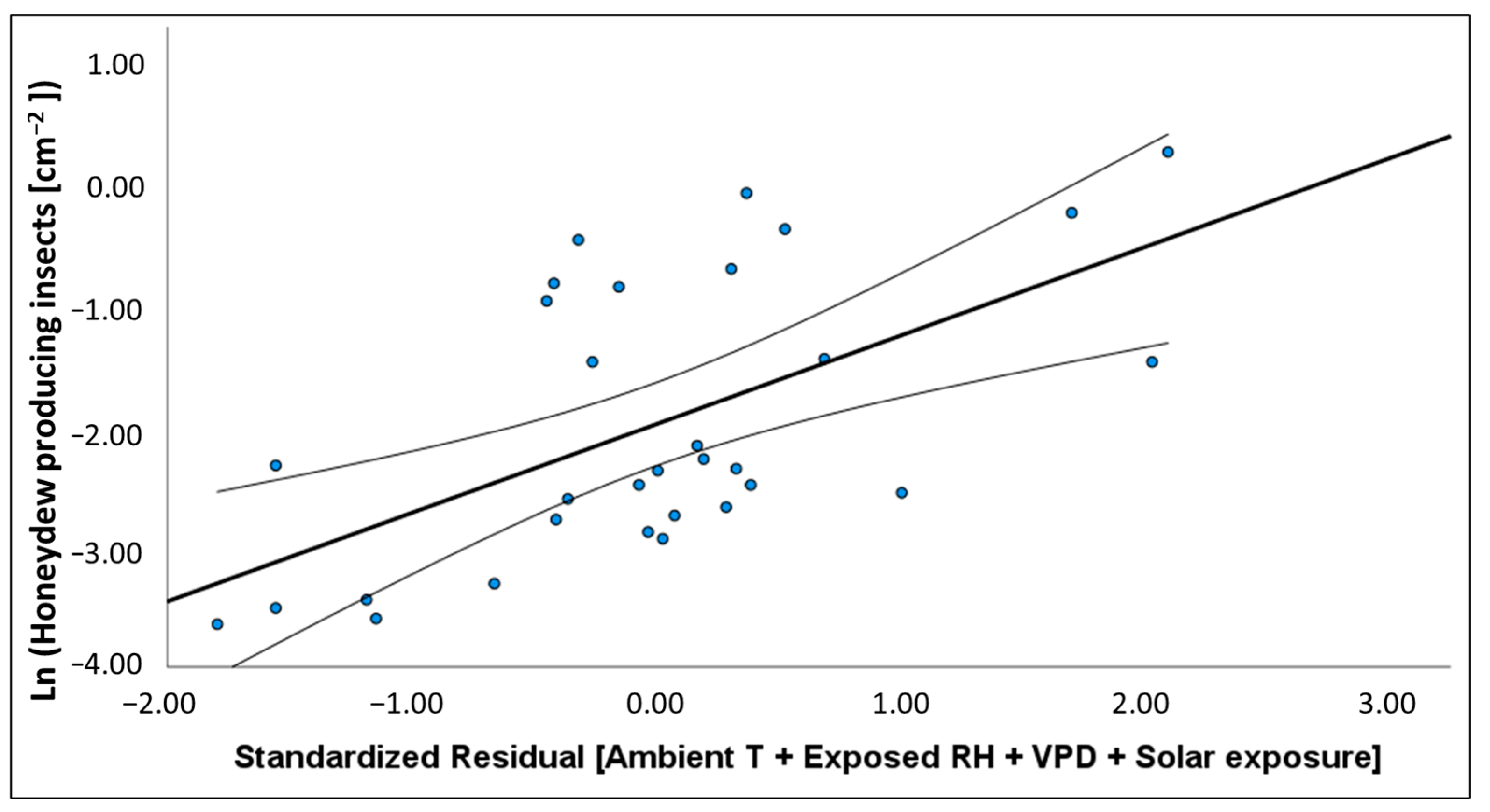
In Australia, variation in the density of
M. hellenica nymphs and proportion of honeydew producing insects (HPI [%]) were associated with warmer and drier conditions. HPI [%] in Australia was often higher over winter than reported in Greece [
50]; however, unlike honeydew availability in Greece, there were no instances where 100% of
M. hellenica sampled in Australia were excreting honeydew. Broadly, Australian HPI [%] followed similar seasonal patterns to Mediterranean observations, including an increase in honeydew availability following overwintering. Since the phenological study from Greece occurred from 2001 to 2003, it is unclear whether the difference in findings between the current study and previous observations are attributable to underlying geographic differences, inherent generational variation, or host species. The insect density recorded in the current study, from 2019–2020, was only separated by six months from insect densities recorded from Greek conspecifics in 2018–2019 [
51]. However, the density of Australian
M. hellenica was generally higher than the density of the concurrent Greek conspecifics. Insect density, HPI [%], and HPI [cm
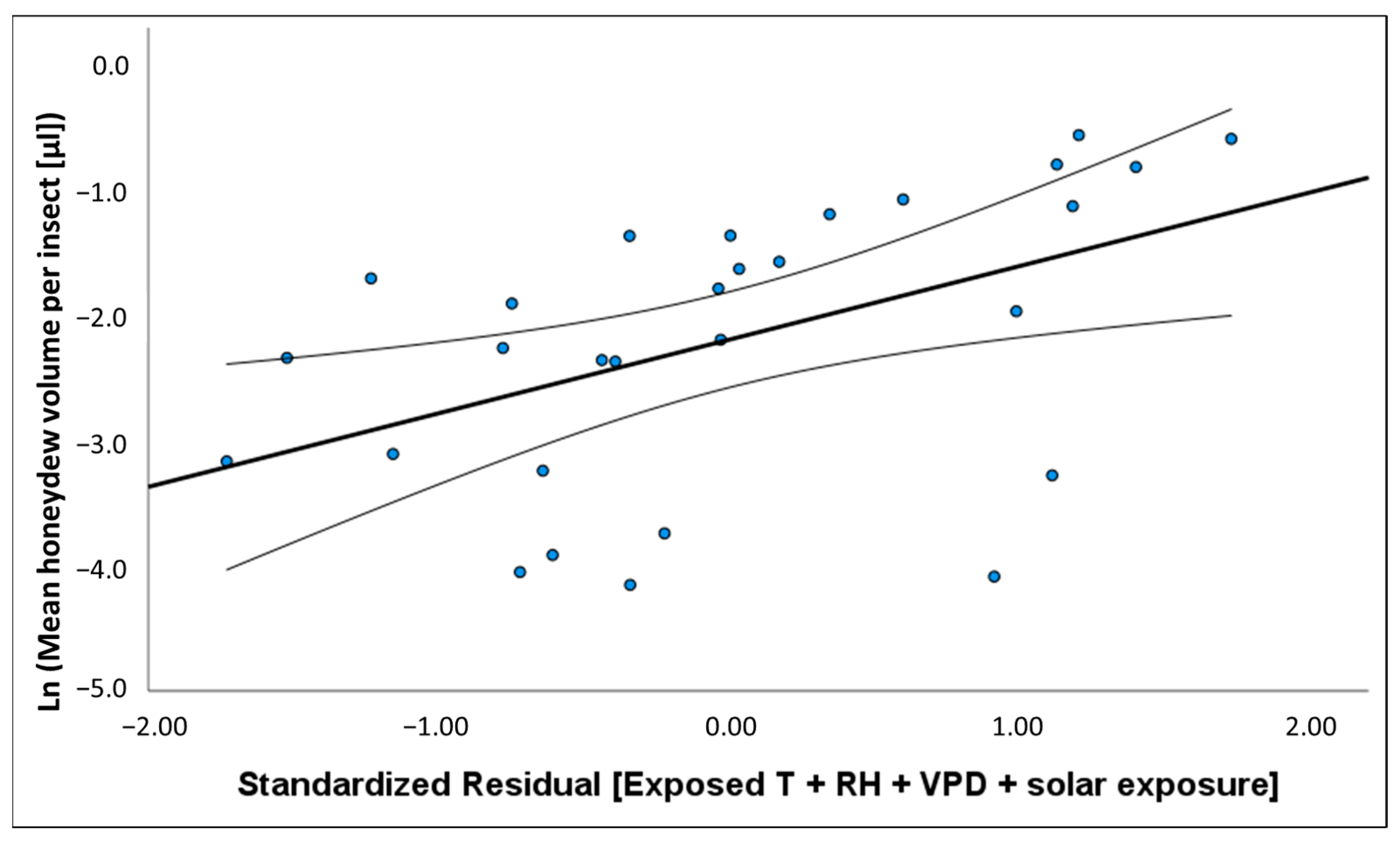
−2] showed a positive relationship with temperature and atmospheric vapour pressure deficit (VPD) and negative association with rainfall and RH. The volume of honeydew produced per insect had the opposite associations with these same climate variables. These relationships with environmental conditions may explain why
M. hellenica in Australia achieved greater densities than Mediterranean populations.
Marchalina hellenica in Australia may occupy a more solar exposed, warmer, and drier climate than Mediterranean conspecifics, which are conducive to
Pinus growth [
60,
61] and, by association, tree quality as a host for herbivores [
62,
63]. Unlike previous studies showing microclimate better reflects insect ontogeny, compared to macroclimate records [
37], climate conditions within the microhabitat of settled
M. hellenica nymphs in this study were usually not the best indicator of changes in insect activity. This may be due to strong climate effects on the host’s activity [
64,
65], and therefore, the availability of resources for
M. hellenica. The influence of host quality on phloem-feeder activity may therefore be amplified because settled
M. hellenica are sheltered from direct effects of forest climate condition and depend on climate-driven phloem enrichment [
66]. Indeed, phloem-feeder honeydew production and density can fluctuate in association with daily cycles of phloem enrichment [
13] and chemical defensive responses [
51].
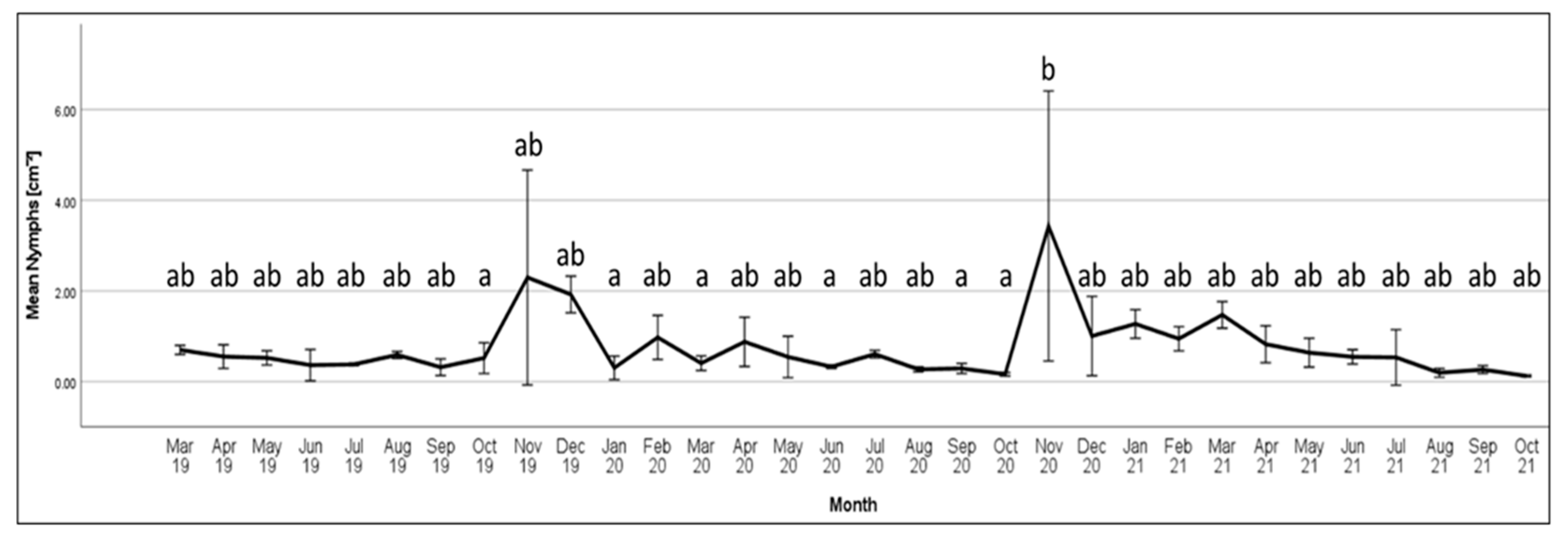
The highest recorded density for Australian
M. hellenica in this study was over 5
M. hellenica per cm
2, compared to the highest density recorded in Greece of approximately 1.4
M. hellenica per cm
2 [
51]. Both of these maximum density records coincided with first instar emergence, and at other times, Greek
M. hellenica densities were close to zero for most months, and almost always less than 0.25
M. hellenica per cm
2. By contrast, Australian density presented here was 0.5 per cm
2 or higher across most months. One explanation for elevated
M. hellenica density in Australia compared to Greece may be that the insect has escaped its natural enemies. The notable enemy is the predatory fly,
Neoleucopis kartliana, a monophagous predator and prospective biocontrol agent [
53,
67].
Neoleucopis kartliana is recorded as being the most effective and widely distributed predator of
M. hellenica in Turkey [
32]. With
N. kartliana absent,
M. hellenica populations in Australia may proliferate with relatively little impact from natural enemies [
68,
69,
70]. If
N. kartliana is deemed suitable for introduction into Australia as a classical biocontrol agent, inoculative mass-releases of this fly may be guided by the seasonal patterns of Australian
M. hellenica documented here. For example, the effectiveness of biological control agents can be heavily dependent on the presence of particular life stages, not just any individual, from a given target species [
71]. This may be critical to any future release of
N. kartliana, as the timing of its phenology matches the seasonal phenology of
M. hellenica [
53].
Host species is also likely important for
M. hellenica growth and survival. In the Mediterranean,
Pinus halepensis is the primary host for
M. hellenica, a tree that shares a long history of hosting
M. hellenica, which provide honeydew as an alternative to floral nectar for apicultural purposes [
17]. This relationship dates back to the late Roman and early Byzantine Empires along the east coast of the Mediterranean Sea [
19,
29,
72]. A long co-evolutionary history may also be implied by
P. halepensis possessing targeted defence responses that coincide with seasonal nymph emergence [
51,
73]. With
M. hellenica recorded as acquiring the novel host in 2014 [
16],
P. radiata in Australia may be lacking specific defence adaptations that have evolved in
P. halepensis. Even if
P. radiata has a similar seasonal-specific defence, the shifting phenology of
M. hellenica in Australia may allow the insect to escape a temporally restricted defence by emerging earlier. A similar process is documented in other insects (e.g., aphids, bark beetles, Heteroptera), notably involving interactive effects with regional climate [
74,
75,
76] or broadscale climate change [
5,
42,
77].
This study’s findings are similar to previous investigations linking climate to changes in both plant and herbivore activity [
76,
78,
79,
80,
81]. Climate effects may therefore affect the quality of
M. hellenica as a host for predatory insects, as has been observed in parasitic/predatory Diptera [
82,
83].
Marchalina hellenica occurs in areas of similar climate conditions to its endemic Greek distribution [
67] and is largely dependent on
P. radiata phloem feeding—the quality of which is influenced by climate conditions [
74,
84]. For example, VPD is known to be a driver of daily cycles of transpiration and photosynthesis in
Pinus [
40,
85,
86] and responses to novel climate conditions may alter phloem quality for herbivores. In
Pinus, these responses are a result of tight regulation of stomatal pore closure and, therefore, strict responses in transpiration, photosynthetic activity, or enrichment of phloem elements [
87,
88,
89]. With such rapid responses to environmental changes,
M. hellenica, by expropriating carbohydrates, amino acids, and other macromolecules [
24], may illicit these strict responses [
90] and potentially exacerbate stressful climate effects on the host tree. As global climate conditions deviate from previous means, this insect may experience changes in phenological timing and feeding activity [
5,
7,
91]. Investigation of how
M. hellenica life cycle and feeding activity may respond to changes to environmental conditions in its endemic geographic range or on native hosts would provide valuable comparisons with the relationships identified in Melbourne populations.





{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}










