
Population Density and Host Preference of the Japanese Pine Sawyer (Monochamus alternatus) in the Qinling–Daba Mountains of China


 and
and
Abstract
:Simple Summary
Abstract
1. Introduction
2. Materials and Methods
2.1. Experimental Field
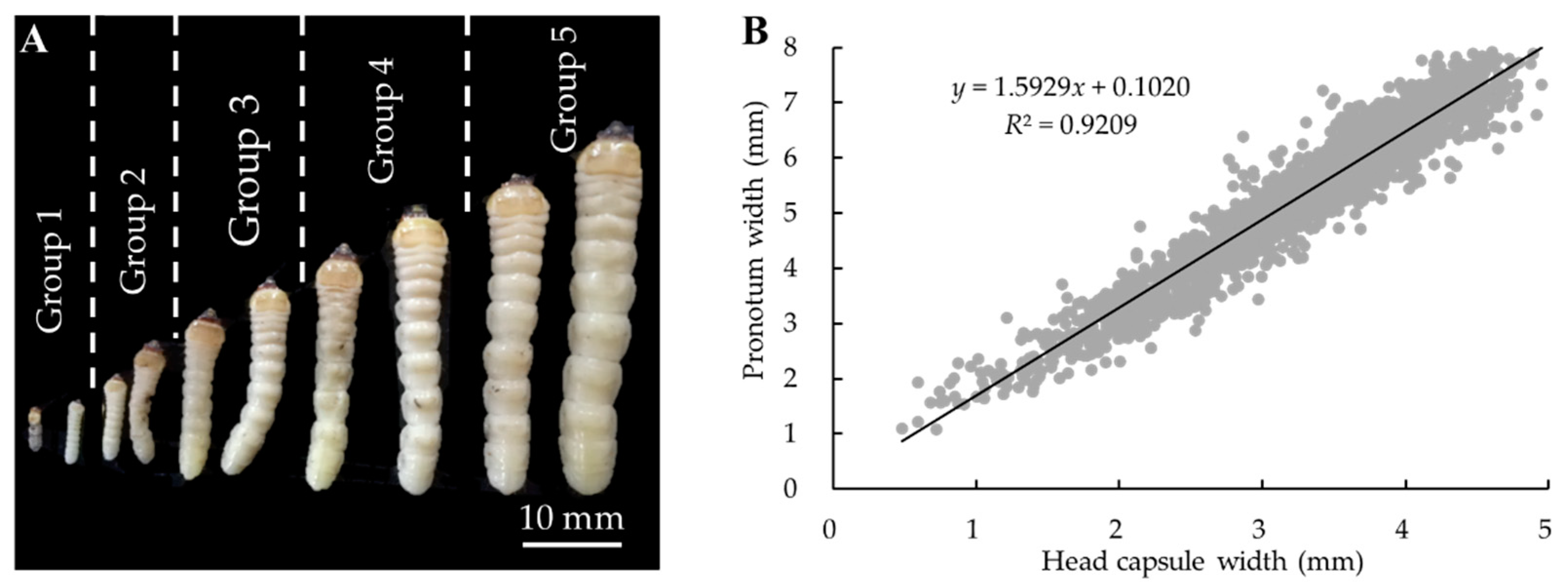
2.2. Population Density and Development Stages of M. alternatus on Different Host Plants
2.3. Feeding and Oviposition Selection of M. alternatus Adults
2.4. Statistical Analysis
3. Results
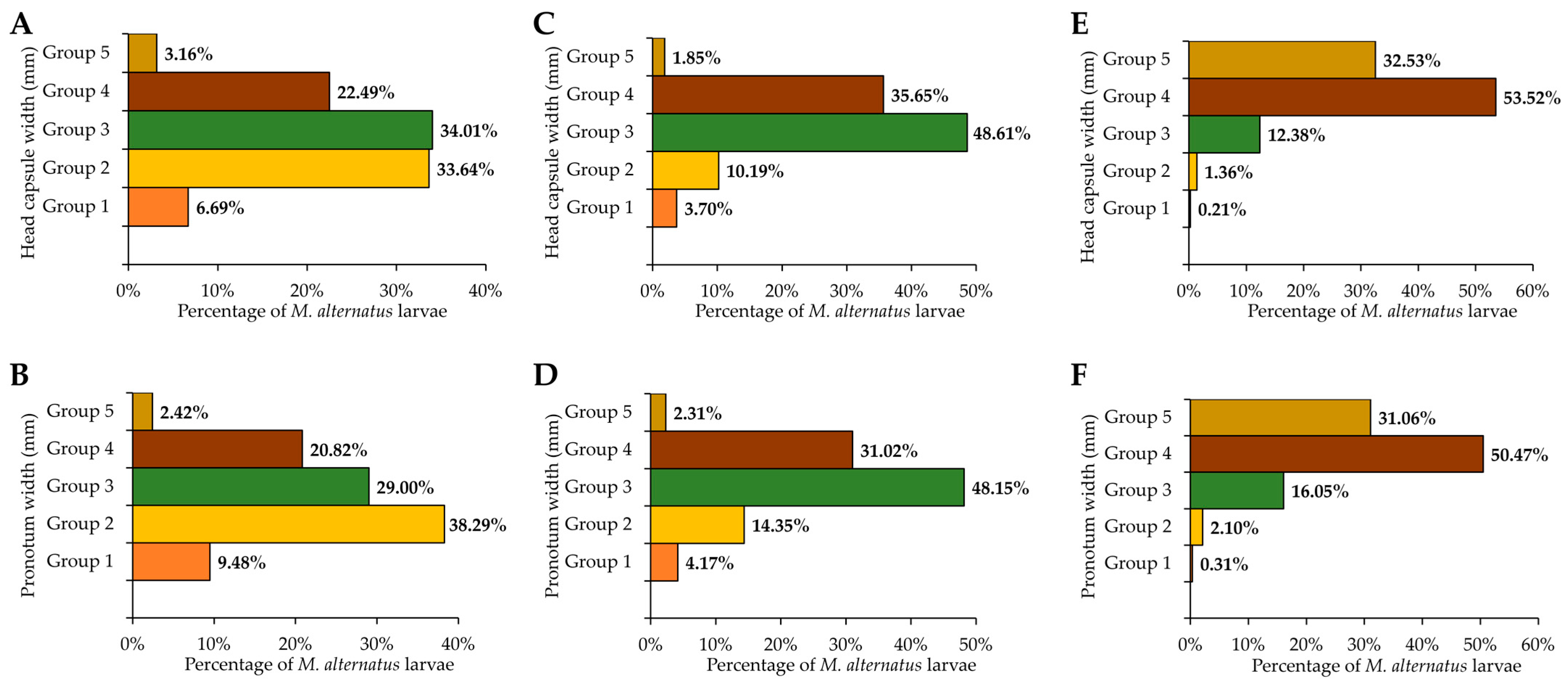
3.1. Population Density and Developmental Stages of M. alternatus Overwintering Larvae on Different Host Plants
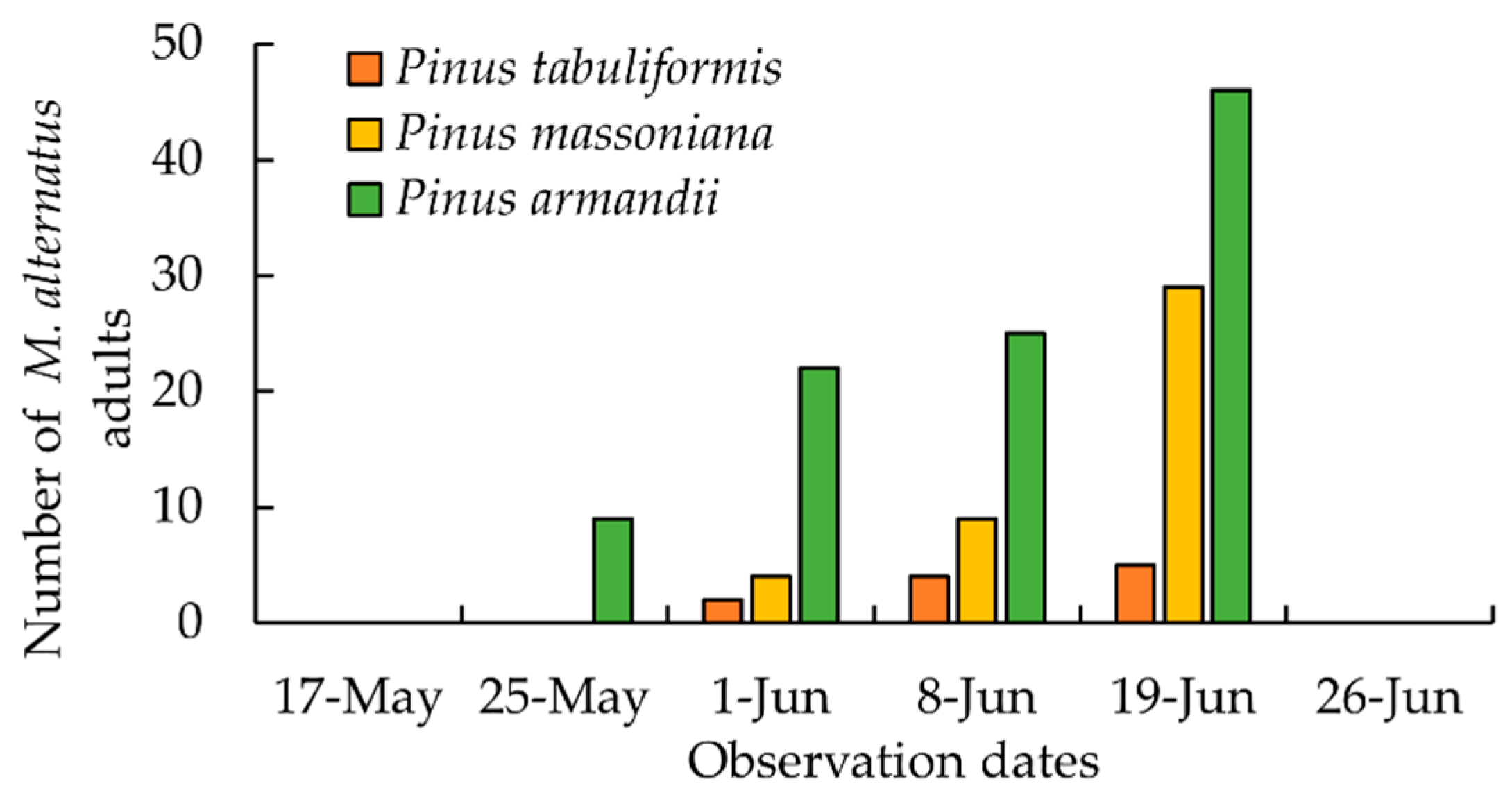
3.2. The Emergence of M. alternatus Adults on Different Pine Trees
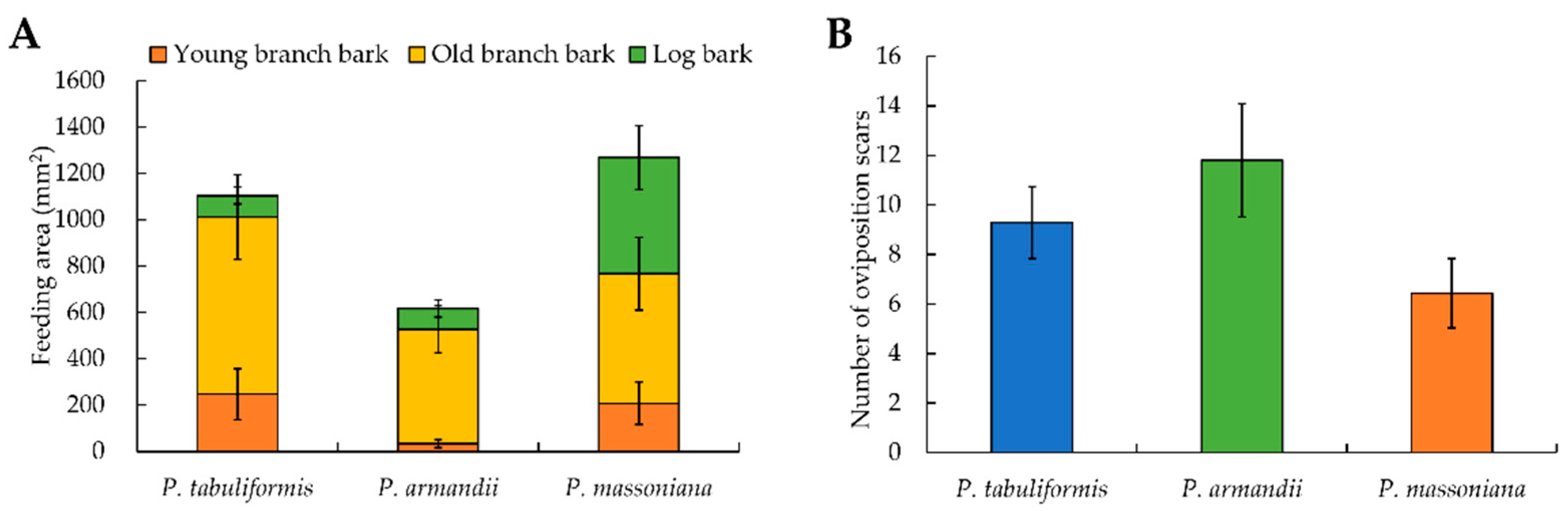
3.3. Feeding Preference of M. alternatus Adults
3.4. Oviposition Selection of M. alternatus Adults
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Linit, M.J. Nematode–vector relationships in the pine wilt disease system. J. Nematol. 1988, 20, 227–235. [Google Scholar] [PubMed]
- Akbulut, S.; Stamps, W.T. Insect vectors of the pinewood nematode: A review of the biology and ecology of Monochamus species. For. Pathol. 2012, 42, 89–99. [Google Scholar] [CrossRef]
- Li, H.; Shen, P.; Fu, P.; Lin, M.; Moens, M. Characteristics of the emergence of Monochamus alternatus, the vector of Bursapelenchus xylophilus (Nematoda: Aphelenchoididae), from Pinus thunbergii logs in Nanjing, China, and of the transmission of the nematodes through feeding wounds. Nematology 2007, 9, 807–816. [Google Scholar]
- Futai, K. Pine wood nematode, Bursaphelenchus xylophilus. Annu. Rev. Phytopathol. 2013, 51, 61–83. [Google Scholar] [CrossRef] [PubMed]
- Filipiak, A.; Malewski, T.; Matczyńska, E.; Tomalak, M. Molecular variation among virulent and avirulent strains of the quarantine nematode Bursaphelenchus xylophilus. Mol. Genet. Genom. 2021, 296, 259–269. [Google Scholar] [CrossRef] [PubMed]
- Zhao, L.; Mota, M.; Vieira, P.; Butcher, R.A.; Sun, J. Interspecific communication between pinewood nematode, its insect vector, and associated microbes. Trends Parasitol. 2014, 30, 299–308. [Google Scholar] [CrossRef] [PubMed]
- Wang, Y.; Chen, F.; Wang, L.; Min, L. Investigation of beetle species that carry the pine wood nematode, Bursaphelenchus xylophilus (Steiner and Buhrer) Nickle, in China. J. For. Res. 2021, 23, 981–987. [Google Scholar] [CrossRef]
- Sun, Y. First report of pine wilt disease from Pinus thunbergii Parl in Sun Yat-sen Mausoleum. J. Jiangsu Univ. Sci. Technol. 1982, 2, 27. [Google Scholar]
- Ren, Z.; Zhang, Y.; Li, L. The value of vegetation ecosystem services: A case of Qinling–Daba Mountains. J. Geogr. Sci. 2003, 2, 195–200. [Google Scholar]
- Wang, B.; Jiang, J.P.; Xie, F.; Li, C. Postglacial colonization of the Qinling Mountains: Phylogeography of the swelled vent frog (Feirana quadranus). PLoS ONE 2012, 7, e41579. [Google Scholar] [CrossRef]
- Zhao, F.; Liu, J.; Zhu, W.; Zhang, B.; Zhu, L. Spatial variation of altitudinal belts as dividing index between warm temperate and subtropical zones in the Qinling–Daba Mountains. J. Geogr. Sci. 2020, 30, 642–656. [Google Scholar] [CrossRef]
- Lian, J.; Zhang, X. Forest pest ecological control. Sci. Silvae Sin. 2005, 41, 168–176. [Google Scholar]
- Tu, Y.G.; Yu, A.L.; Que, S.Q.; Yu, N.F.; Liu, X.H. Effects of different host plants on longevity and reproduction of Monochamus alternatus Hope adults. Southwest China J. Agric. Sci. 2019, 32, 1801–1804. [Google Scholar]
- Bonifácio, L.; Naves, P.; Sousa, E. Vector-plant. In Pine Wilt Disease in Europe: Biological Interactions and Integrated Management; Sousa, E., Vale, F., Abrantes, I., Eds.; FNAPF-Federação Nacional das Associações de Proprietários Florestais: Lisboa, Portugal, 2015; pp. 125–157. [Google Scholar]
- Gonalves, E.; Figueiredo, A.C.; Barroso, J.G.; Henriques, J.; Sousa, E.; Bonifácio, L. Effect of Monochamus galloprovincialis feeding on Pinus pinaster and Pinus pinea, oleoresin and insect volatiles. Phytochemistry 2019, 169, 112159. [Google Scholar] [CrossRef] [PubMed]
- Nan, J.K.; Yang, Y.X.; Zhang, L.H.; Li, P.F.; Guo, L.J.; He, H.; Wei, C. The damage of Monochamus alternatus on different host pine species in Qinling–Daba Mountains. J. Environ. Entomol. 2021, 43, 1376–1388. [Google Scholar]
- Jiang, S.N. Cerambycid Larvae of China; Chongqing Publishing House: Chongqing, China, 1989; pp. 43–44. [Google Scholar]
- Yang, X.K.; Lin, M.Y. Insect Fauna of the Qinling Mountains. 6. Coleoptera. 2. Longicorn; World Book Publishing Xi’an Co., Ltd.: Xi’an, China, 2017; p. 329. [Google Scholar]
- Caltagirone, L.E.; Getz, W.; Meals, D.W. Head capsule width as an index of age in larvae of navel orangeworm, Amyelois stansitella (Lepidoptera: Pyralidae). Environ. Entomol. 1983, 12, 219–221. [Google Scholar] [CrossRef]
- Beaver, R.J.; Sanderson, J.P. Classifying instars of the Navel orangeworm (Lepidoptera: Pyralidae) based on observed head capsule widths. J. Econ. Entomol. 1989, 82, 716–720. [Google Scholar] [CrossRef]
- McClellan, Q.C.; Logan, J.A. Instar determination for the Gypsy moth (Lepidoptera: Lymantriidae) based on the frequency distribution of head capsule widths. Environ. Entomol. 1994, 23, 248–253. [Google Scholar] [CrossRef]
- Zhou, Z.J.; Togashi, K. Oviposition and larval performance of Monochamus alternatus (Coleoptera: Cerambycidae) on the Japanese cedar Cryptomeria japonica. J. For. Res. 2006, 11, 35–40. [Google Scholar] [CrossRef]
- Li, M.; Dai, Y.; Wang, Y.; Wang, L.; Sun, S.; Chen, F. New insights into the life history of Monochamus saltuarius (Cerambycidae: Coleoptera) can enhance surveillance strategies for pine wilt disease. J. For. Res. 2021, 32, 2699–2707. [Google Scholar] [CrossRef]
- Togashi, K. Development of Monochamus alternatus Hope (Coleoptera: Cerambycidae) in relation to oviposition time. Jpn. J. Appl. Entomol. Zool. 1989, 33, 1–8. [Google Scholar] [CrossRef]
- Hwang, I.C.; Kim, J.H.; Park, J.B.; Shin, S.C.; Park, Y.C. Growth, development, and reproduction of Monochamus saltuarius (Coleoptera: Cerambycidae) on conifers fed to larvae. J. Appl. Entomol. 2008, 47, 385–394. [Google Scholar] [CrossRef]
- Togashi, K. Influence of oviposition timing on offspring diapause and growth in Monochamus alternatus endai (Coleoptera: Cerambycidae). J. For. Res. 2019, 24, 313–319. [Google Scholar] [CrossRef]
- Togashi, K. Effects of three host pine species on diapause induction and life–history traits of Monochamus alternatus (Coleoptera: Cerambycidae). J. For. Res. 2021, 26, 230–236. [Google Scholar] [CrossRef]
- Han, J.H.; Kim, H.K.; Kang, W.J.; Kim, G.H. Feeding and oviposition preference of the Sakhalin pine sawyer Monochamus saltuarius (Coleoptera: Cerambycidae) for different hosts. Entomol. Res. 2016, 46, 331–336. [Google Scholar] [CrossRef]
- Go, M.S.; Kwon, S.H.; Kim, S.B.; Kim, D.S. The developmental characteristics for the head capsule width of Monochamus alternatus (Coleoptera: Cerambycidae) larvae and determination of the number of instars. J. Insect Sci. 2019, 19, 26. [Google Scholar] [CrossRef] [Green Version]
- Kozima, K.; Katagiri, K. On the larval instar and changes of its composition of Monochamus alternalus Hope. J. Jpn. For. Soc. 2008, 46, 307–310. [Google Scholar]
- Liu, J.D.; Li, B.W.; Wang, J.Y.; Zhao, B.A.; Shu, J.P. Primary study on method to determine instars of Monochamus alternatus larvae. J. Zhejiang For. Sci. Technol. 2008, 28, 84–86. [Google Scholar]
- Guo, Y.Q.; Xin, Y.C.; Tao, X.; Yu, H.; Liu, Z.D. Body size difference of male and female adults as well as the relationship between the sizes of pupae and adults and the body weight of overwintering larvae in the Japanese pine sawyer, Monochamus alternatus (Coleoptera: Cerambycidae). Acta Entomol. Sin. 2015, 58, 989–996. [Google Scholar]
- Kim, S.B.; Kim, D.S. A new direction to understand the life cycle of the Japanese pine sawyer considering the selection strategy of instar pathways. Sci. Rep. 2020, 10, 16952. [Google Scholar] [CrossRef]
- Dyar, H.G. The number of molts of lepidopterous larvae. Psyche A J. Èntomol. 1890, 5, 420–422. [Google Scholar] [CrossRef]
- Hou, Z.; Li, Q.; Yang, M.; Liu, Y.; Wei, C. Ecology of Meimuna mongolica (Hemiptera: Cicadidae) nymphs: Instars, morphological variation, vertical distribution and population density, host-plant selection, and emergence phenology. J. Insect Sci. 2015, 15, 1–6. [Google Scholar] [CrossRef]
- Kang, S.H.; Kim, M.K.; Yang, J.O.; Yoon, C.; Goh, S.H.; Shin, S.C.; Kim, C.S.; Kim, G.H. Attraction effects to various conditions of Pinus koraiensis against pine sawyer, Monochamus saltuarius (Coleoptera: Cerambycidae). J. Korean Soc. Appl. Biol. Chem. 2009, 52, 456–465. [Google Scholar] [CrossRef]
- Ginzel, M.D.; Hanks, L.M. Role of host plant volatiles in mate location for three species of longhorned beetles. J. Chem. Ecol. 2005, 31, 213–217. [Google Scholar] [CrossRef]
- Li, S.Q.; Fang, Y.L.; Zhang, Z.N. Effects of volatiles of non–host plants and other chemicals on oviposition of Monochamus alternatus (Coleoptera: Cerambycidae). J. Pest Sci. 2007, 80, 119–123. [Google Scholar] [CrossRef]
- Fan, J.; Sun, J.; Shi, J. Attraction of the Japanese pine sawyer, Monochamus alternatus, to volatiles from stressed host in China. Ann. For. Sci. 2007, 64, 67–71. [Google Scholar] [CrossRef]
- Vicente, C.S.L.; Soares, M.; Faria, J.M.S.; Ramos, A.P.; Inácio, M.L. Insights into the role of fungi in pine wilt disease. J. Fungi 2021, 7, 780. [Google Scholar] [CrossRef]
- Hao, D.J.; Zhang, Y.H.; Dai, H.G.; Wang, Y. Oviposition preference of Monochamus alternatus Hope (Coleoptera: Cerambycidae) to host plants. Acta Entomol. Sin. 2005, 48, 460–464. [Google Scholar]






{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Surveying Sites | Tree Species | Longitude (E) | Latitude (N) | Altitude (m) | ||
|---|---|---|---|---|---|---|
| Qinling Mountains | Zhashui County | Fenghuang Town | P. tabuliformis, P. armandii | 109°21′34″ | 33°43′59″ | 659.00 |
| Yingpan Town | P. armandii | 109°06′03″ | 33°74′21″ | 931.33 | ||
| Xialiang Town | P. tabuliformis | 109°08′54″ | 33°37′42″ | 736.06 | ||
| Foping County | Daheba Town | P. tabuliformis, P. massoniana | 108°61′34″ | 33°29′43″ | 769.05 | |
| Ningshan County | Chengguan Town | P. tabuliformis, P. armandii | 108°19′48″ | 33°18′00″ | 807.02 | |
| Daba Mountains | Xixiang County | Yankou Town | P. massoniana | 107°50′42″ | 32°59′46″ | 509.88 |
| Langao County | Chengguan Town | P. massoniana | 108°54′35″ | 32°18′30″ | 758.63 | |
| Zuolong Town | P. massoniana | 108°53′08″ | 32°30′19″ | 457.31 | ||
| Tree Species | N | Number of Monochamus alternatus Larvae ± SE | Variance Analysis |
|---|---|---|---|
| Pinus tabuliformis | 10 | 73.00 ± 21.81 b | F = 10.480, p = 0.000 |
| Pinus massoniana | 14 | 91.21 ± 18.80 b | |
| Pinus armandii | 14 | 235.50 ± 36.06 a |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Nan, J.; Qi, J.; Yang, Y.; Zhao, M.; Liang, C.; He, H.; Wei, C. Population Density and Host Preference of the Japanese Pine Sawyer (Monochamus alternatus) in the Qinling–Daba Mountains of China. Insects 2023, 14, 181. https://doi.org/10.3390/insects14020181
Nan J, Qi J, Yang Y, Zhao M, Liang C, He H, Wei C. Population Density and Host Preference of the Japanese Pine Sawyer (Monochamus alternatus) in the Qinling–Daba Mountains of China. Insects. 2023; 14(2):181. https://doi.org/10.3390/insects14020181
Chicago/Turabian StyleNan, Junke, Jingyu Qi, Yuexiang Yang, Mengqin Zhao, Chaoqiong Liang, Hong He, and Cong Wei. 2023. "Population Density and Host Preference of the Japanese Pine Sawyer (Monochamus alternatus) in the Qinling–Daba Mountains of China" Insects 14, no. 2: 181. https://doi.org/10.3390/insects14020181