

The Pattern of Social Parasitism in Maculinea teleius Butterfly Is Driven by the Size and Spatial Distribution of the Host Ant Nests

Abstract
:Simple Summary
Abstract
1. Introduction
2. Materials and Methods
2.1. Study Areas
2.2. Field Survey
2.3. Statistical Analyses
3. Results
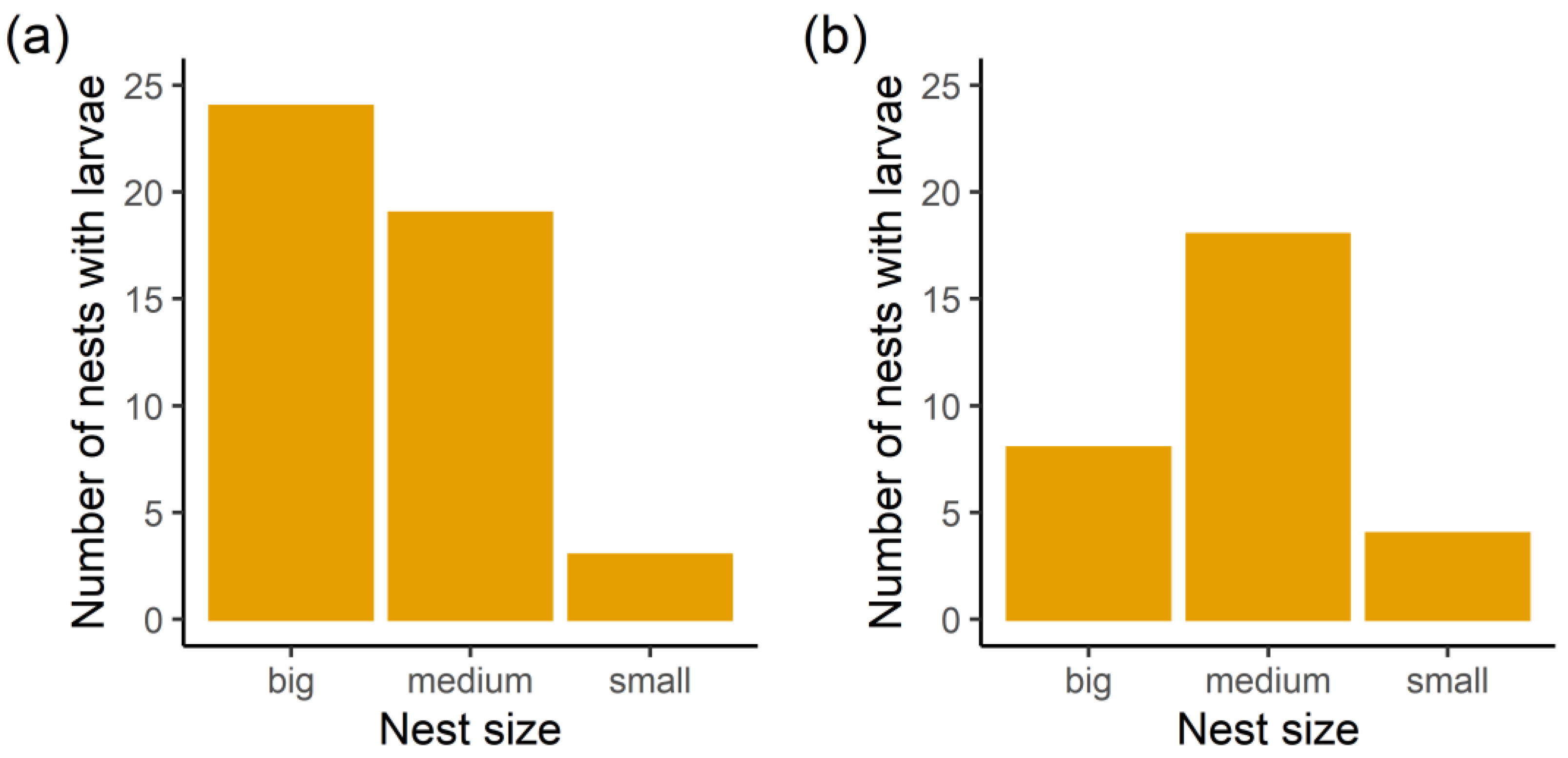
3.1. Infestation Rates
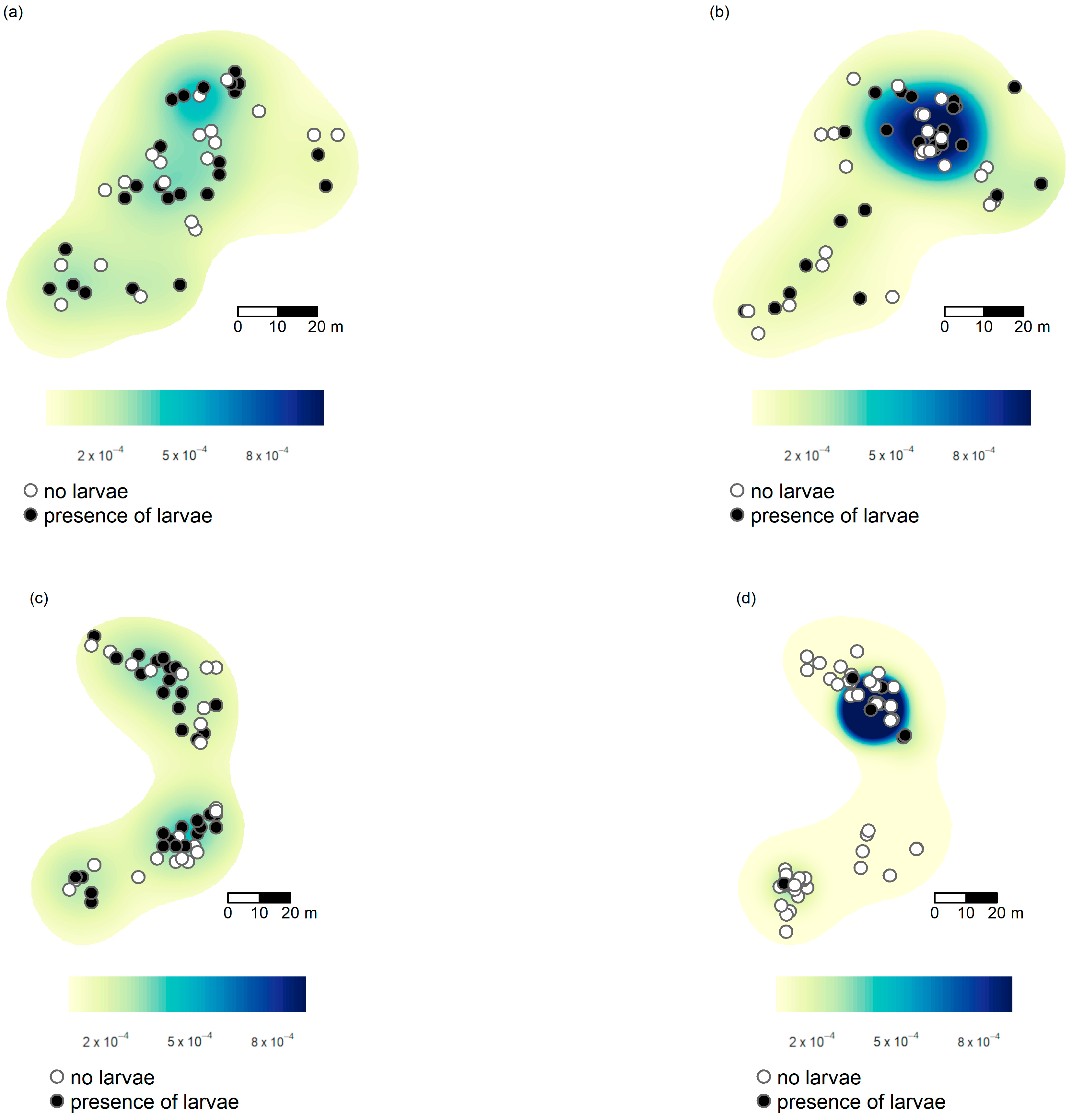
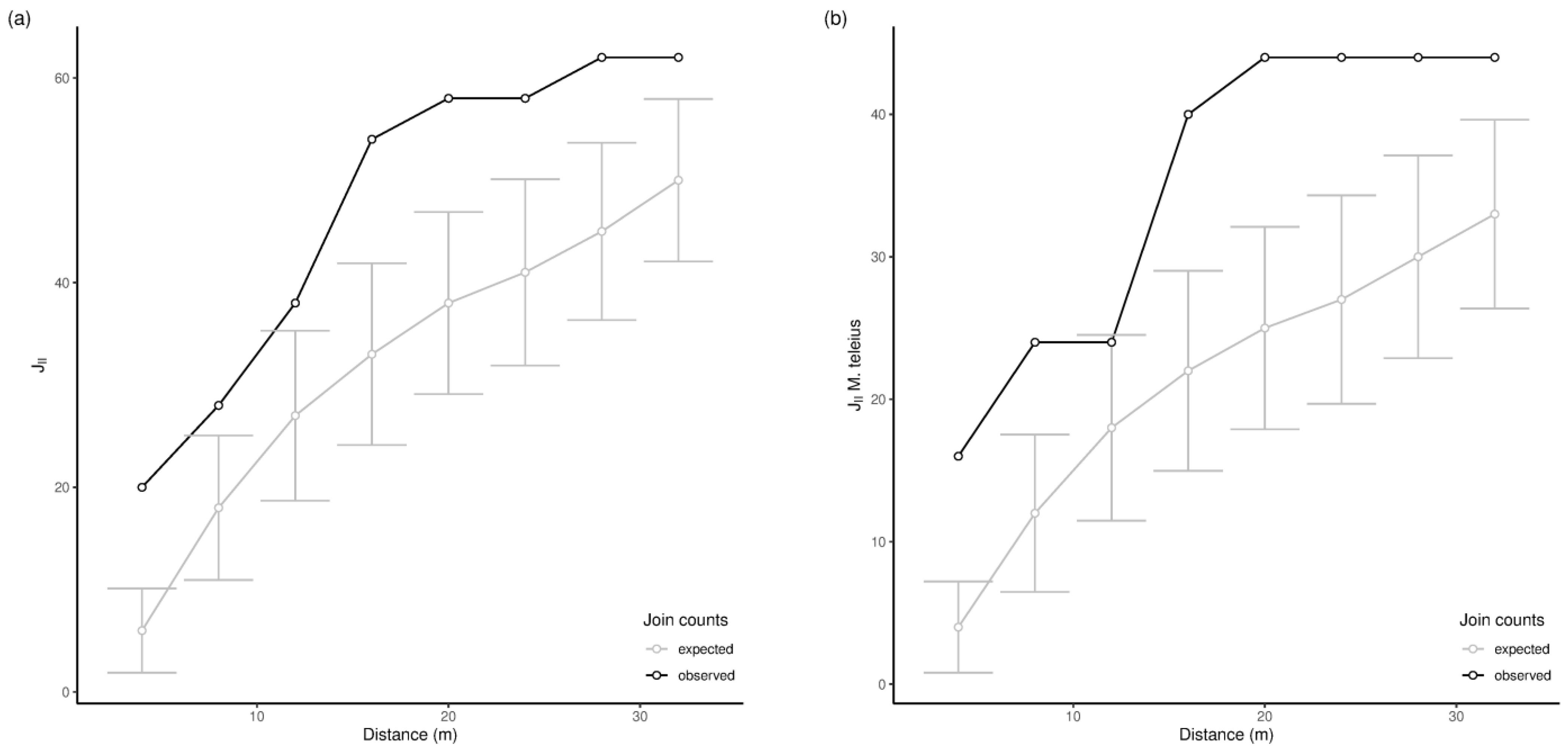
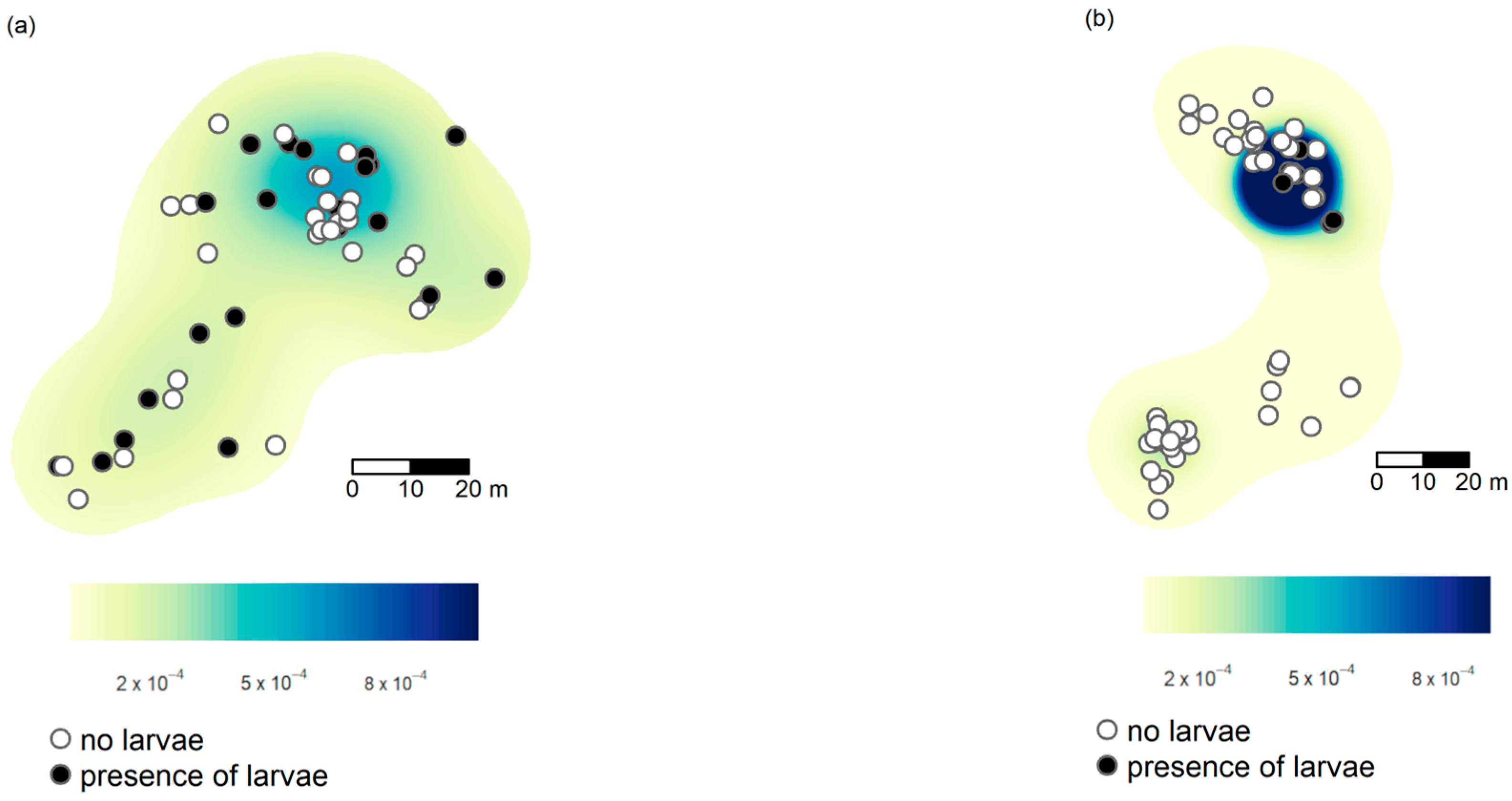
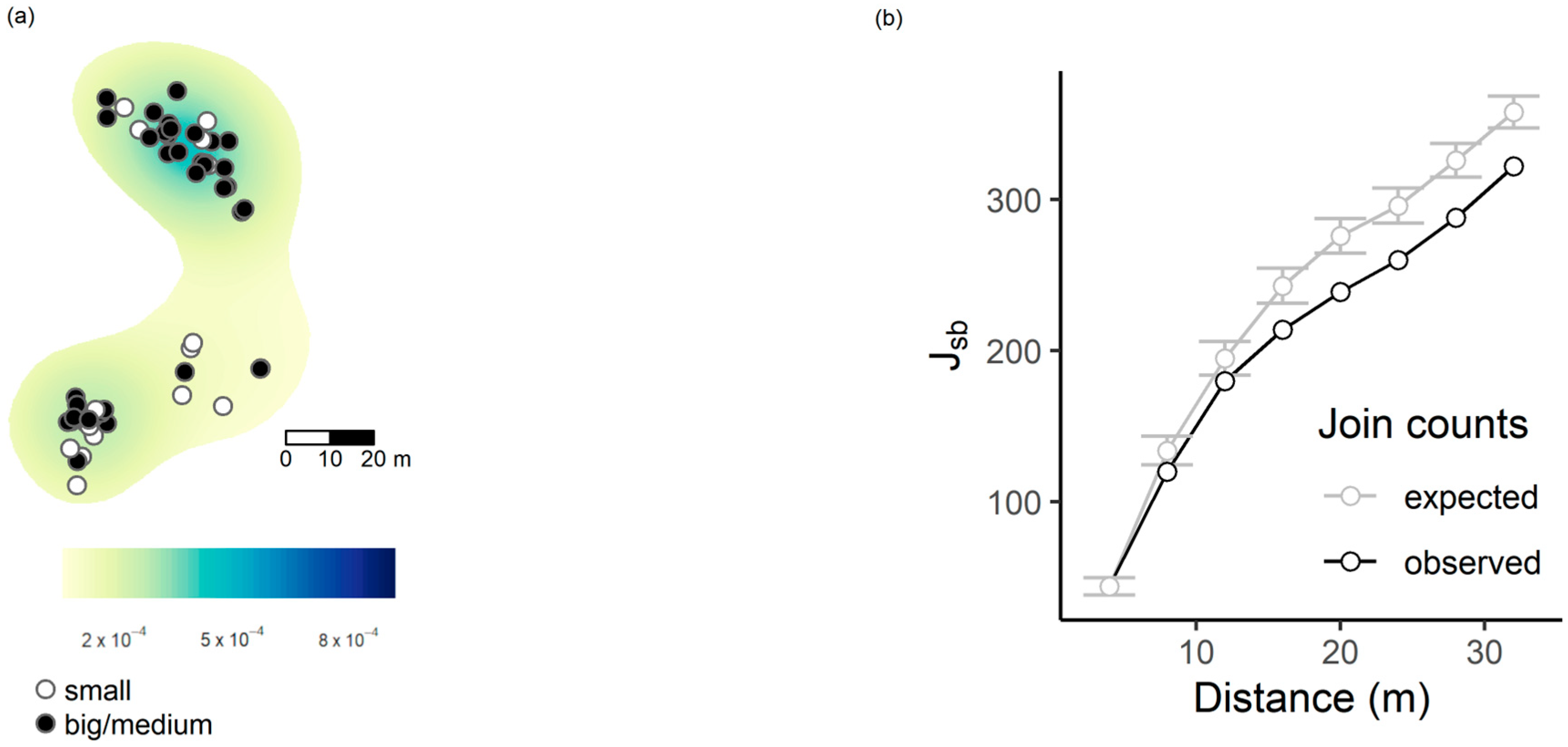
3.2. Spatial Patterns
3.3. Factors Correlated with the Presence of Ma. teleius and Mi. myrmicae Larvae Inside Myrmica Nests
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Hölldobler, B.E.; Wilson, E.O. The Ants; The Belknap Press of Harvard University Press: Cambridge, MA, USA, 1990. [Google Scholar]
- Pierce, N.E.; Dankowicz, E. Behavioral, ecological and evolutionary mechanisms underlying caterpillar-ant symbioses. Curr. Opin. Insect. Sci. 2022, 52, 100898. [Google Scholar] [CrossRef] [PubMed]
- New, T.R. Mutualisms and Insect Conservation; Springer: Cham, Switzerland, 2017. [Google Scholar]
- Hölldobler, B.; Kwapich, C.L. The Guests of Ants: How Myrmecophiles Interact with Their Hosts; Harvard University Press: Cambridge, MA, USA, 2022. [Google Scholar]
- Thomas, J.A.; Schönrogge, K.; Elmes, G.W. Specializations and host associations of social parasites of ants. In Insect Evolutionary Ecology; Fellowes, M.D.E., Holloway, G.J., Rolff, J., Eds.; Royal Entomological Society: London, UK, 2005; pp. 475–514. [Google Scholar]
- Radchenko, A.G.; Elmes, G.W. Myrmica (Hymenoptera: Formicidae) Ants of the Old World; Fauna Mundi 3; Natura Optima Dux Foundation: Warsaw, Poland, 2010; Volume 6, ISBN 9788393077311. [Google Scholar]
- Buschinger, A. Social parasitism among ants: A review (Hymenoptera: Formicidae). Myrmecol. News 2009, 12, 219–235. [Google Scholar]
- Bharti, H. Myrmica nefaria sp.n. (Hymenoptera: Formicidae)—A new social parasite from Himalaya. Myrmecol. News 2012, 16, 149–156. [Google Scholar]
- Bharti, H.; Radchenko, A.; Sasi, S. Socially-parasitic Myrmica species (Hymenoptera, Formicidae) of Himalaya, with the description of a new species. ZooKeys 2016, 605, 113. [Google Scholar] [CrossRef] [PubMed]
- Casacci, L.P.; Barbero, F.; Ślipiński, P.; Witek, M. The inquiline ant Myrmica karavajevi uses both chemical and vibroacoustic deception mechanisms to integrate into its host colonies. Biology 2021, 10, 654. [Google Scholar] [CrossRef] [PubMed]
- Kutter, H. Über die morphologischen Beziehungen der Gattung Myrmica zu ihren Satellitengenera Sifolinia Em., Symbiomyrma Arnoldi und Sommimyrma Menozzi. Mitt. Schweiz. entomol. Ges. 1973, 46, 253–268. [Google Scholar]
- Radchenko, A.G.; Elmes, G.W. A taxonomic revision of the socially parasitic Myrmica ants (Hymenoptera: Formicidae) of the Palaearctic region. Ann. Zool. 2003, 53, 217–243. [Google Scholar]
- Jansen, G.; Savolainen, R.; Vepsäläinen, K. Phylogeny, divergence-time estimation, biogeography and social parasite-host relationships of the Holarctic ant genus Myrmica (Hymenoptera: Formicidae). Mol. Phylogenet. Evol. 2010, 56, 294–304. [Google Scholar] [CrossRef]
- Witek, M.; Barbero, F.; Markó, B. Myrmica ants host highly diverse parasitic communities: From social parasites to microbes. Insectes Soc. 2014, 61, 307–323. [Google Scholar] [CrossRef]
- Tartally, A.; Thomas, J.A.; Anton, C.; Balletto, E.; Barbero, F.; Bonelli, S.; Bräu, M.; Casacci, L.P.; Csősz, S.; Czekes, Z.; et al. Patterns of host use by brood parasitic Maculinea butterflies across Europe. Philos. Trans. R Soc. B Biol. Sci. 2019, 374, 20180202. [Google Scholar] [CrossRef]
- Thomas, J.A.; Settele, J. Butterfly mimics of ants. Nature 2004, 432, 283–284. [Google Scholar] [CrossRef] [PubMed]
- Thomas, J.A. Why did the large blue become extinct in Britain? Oryx 1980, 15, 243–247. [Google Scholar] [CrossRef]
- Thomas, J.A. The ecology and conservation of Maculinea arion and other European species of large blue butterfly. In Ecology and Conservation of Butterflies; Pullin, A.S., Ed.; Chapman and Hall: London, UK, 1995; pp. 180–197. [Google Scholar]
- Casacci, L.P.; Bonelli, S.; Balletto, E.; Barbero, F. Multimodal signaling in myrmecophilous butterflies. Front. Ecol. Evol. 2019, 7, 454. [Google Scholar] [CrossRef]
- Elmes, G.W.; Akino, T.; Thomas, J.A.; Clarke, R.T.; Knapp, J.J. Interspecific differences in cuticular hydrocarbon profiles of Myrmica ants are sufficiently consistent to explain host specificity by Maculinea (Large Blue) butterflies. Oecologia 2002, 130, 525–535. [Google Scholar] [CrossRef] [PubMed]
- Schönrogge, K.; Barbero, F.; Casacci, L.P.; Settele, J.; Thomas, J.A. Acoustic communication within ant societies and its mimicry by mutualistic and socially parasitic myrmecophiles. Anim. Behav. 2017, 134, 249–256. [Google Scholar] [CrossRef]
- Thomas, J.A.; Wardlaw, J.C. The capacity of a Myrmica ant nest to support a predacious species of Maculinea butterfly. Oecologia 1992, 91, 101–109. [Google Scholar] [CrossRef]
- Thomas, J.A.; Elmes, G.W. Higher productivity at the cost of increased host-specificity when Maculinea butterfly larvae exploit ant colonies through trophallaxis rather than by predation. Ecol. Entomol. 1998, 23, 457–464. [Google Scholar] [CrossRef]
- Stankiewicz, A.; Sielezniew, M. Host specificity of Maculinea teleius Bgstr. and M. nausithous Bgstr. (Lepidoptera: Lycaenidae). The new sight. Ann. Zool. 2022, 52, 403–408. [Google Scholar]
- Woyciechowski, M.; Slowik, J.; Muehlenberg, M. Hosts of the butterfly, Maculinea teleius, among Myrmica ants in Northern Mongolia (Lepidoptera: Lycaenidae; Hymenoptera: Formicidae). Sociobiology 2006, 48, 493–502. [Google Scholar]
- Tartally, A.; Varga, Z. Host ant use of Maculinea teleius in the Carpathian Basin (Lepidoptera: Lycaenidae). Acta Zool. Acad. Sci. Hung. 2008, 54, 257–268. [Google Scholar]
- Witek, M.; Nowicki, P.; Sliwinska, E.B.; Skórka, P.; Settele, J.; Schönrogge, K.; Woyciechowski, M. Local host ant specificity of Phengaris (Maculinea) teleius butterfly, an obligatory social parasite of Myrmica ants. Ecol. Entomol. 2010, 35, 557–564. [Google Scholar] [CrossRef]
- Sielezniew, M.; Stankiewicz-Fiedurek, A.M. Behavioural evidence for a putative oviposition-deterring pheromone in the butterfly, Phengaris (Maculinea) teleius (Lepidoptera: Lycaenidae). Eur. J. Entomol. 2013, 110, 71–80. [Google Scholar] [CrossRef]
- Wynhoff, I.; Grutters, M.; Van Langevelde, F. Looking for the ants: Selection of oviposition sites by two myrmecophilous butterfly species. Anim. Biol. 2008, 58, 371–388. [Google Scholar] [CrossRef]
- Casacci, L.P.; Schönrogge, K.; Thomas, J.A.; Balletto, E.; Bonelli, S.; Barbero, F. Host specificity pattern and chemical deception in a social parasite of ants. Sci. Rep. 2019, 9, 1619. [Google Scholar] [CrossRef]
- Nowicki, P.; Witek, M.; Skórka, P.; Settele, J.; Woyciechowski, M. Population ecology of the endangered butterflies Maculinea teleius and M. nausithous and the implications for conservation. Popul. Ecol. 2005, 47, 193–202. [Google Scholar] [CrossRef]
- Seifert, B. The ecology of Central European non-arboreal ants—37 years of a broad-spectrum analysis under permanent taxonomic control. Soil Org. 2017, 89, 1–67. [Google Scholar]
- Nash, D.R.; Als, T.D.; Maile, R.; Jones, G.R.; Boomsma, J.J. A mosaic of chemical coevolution in a large blue butterfly. Science 2008, 319, 88–90. [Google Scholar] [CrossRef]
- Czechowski, W.; Radchenko, A.; Czechowska, W.; Vepsäläinen, K. The Ants of Poland with Reference to the Myrmecofauna of Europe; Fauna Poloniae (New Series); Natura optima dux Foundation: Warszawa, Poland, 2012; Volume 4, ISBN 978-83-930773-4-2. [Google Scholar]
- Śliwińska, E.B.; Nowicki, P.; Nash, D.R.; Witek, M.; Settele, J.; Woyciechowski, M. Morphology of caterpillars and pupae of European Maculinea species (Lepidoptera: Lycaenidae) with an identification Table. Entomol. Fenn. 2006, 17, 351–358. [Google Scholar] [CrossRef]
- Moran, P.A.P. The interpretation of statistical maps. J. R. Stat. Soc. Ser. B Methodol. 1948, 10, 243–251. [Google Scholar] [CrossRef]
- Cliff, A.D.; Ord, J.K. Spatial Processes: Models and Applications; Pion: London, UK, 1981. [Google Scholar]
- Fortin, M.J.; Dale, M. Spatial Analysis. A Guide for Ecologists; Cambridge University Press: Cambridge, UK, 2005. [Google Scholar]
- R Core Team. R: A Language and Environment for Statistical Computing; R Foundation for Statistical Computing: Vienna, Austria, 2022. [Google Scholar]
- Calenge, C. The package adehabitat for the R software: A tool for the analysis of space and habitat use by animals. Ecol. Model. 2006, 197, 516–519. [Google Scholar] [CrossRef]
- Worton, B.J. Kernel methods for estimating the utilization distribution in home-range studies. Ecology 1989, 70, 164–168. [Google Scholar] [CrossRef]
- Burnham, K.P.; Anderson, D.R. Model Selection and Multimodel Inference: A Practical Information-Theoretic Approach; Springer: New York, NY, USA, 2002. [Google Scholar]
- Dormann, C.F.; McPherson, J.M.; Araújo, M.B.; Bivand, R.; Bollinger, J.; Carl, G.; Davies, R.G.; Hirzel, A.; Jetz, W.; Kissling, W.D.; et al. Methods to account for spatial autocorrelation in the analysis of species distributional data: A review. Ecography 2007, 30, 609–628. [Google Scholar] [CrossRef]
- Zuur, A.F.; Ieno, E.N.; Walker, N.J.; Saveliev, A.A.; Smith, G.M. Mixed Effects Models and Extensions in Ecology with R.; Springer: New York, NY, USA, 2009. [Google Scholar]
- Maák, I.; Camera, J.; Casacci, L.P.; Barbero, F.; Trigos-Peral, G.; Ślipiński, P.; Bonelli, S.; Zaccagno, M.; Witek, M. The influence of colony traits on the collective behaviour of Myrmica scabrinodis ants. Insect Conserv. Diver. 2019, 12, 481–491. [Google Scholar] [CrossRef]
- Gardner, M.G.; Schönrogge, K.; Elmes, G.W.; Thomas, J.A. Increased genetic diversity as a defence against parasites is undermined by social parasites: Microdon mutabilis hoverflies infesting Formica lemani ant colonies. Proc. R. Soc. B. 2007, 274, 103–110. [Google Scholar] [CrossRef] [PubMed]
- Elmes, G.W. Temporal variation in colony populations of the ant Myrmica sulcinodis. II. Sexual production and sex ratios. J. Anim. Ecol. 1987, 56, 573–583. [Google Scholar] [CrossRef]
- Thomas, J.A.; Elmes, G.W.; Wardlaw, J.C. Contest competition among Maculinea rebeli butterfly larvae in ant nests. Ecol. Entomol. 1993, 18, 73–76. [Google Scholar] [CrossRef]
- Witek, M.; Casacci, L.P.; Barbero, F.; Patricelli, D.; Sala, M.; Bossi, S.; Maffei, M.; Woyciechowski, M.; Balletto, E.; Bonelli, S. Interspecific relationships in co-occurring populations of social parasites and their host ants. Biol. J. Linn. Soc. 2013, 109, 699–709. [Google Scholar] [CrossRef]
- Witek, M.; Śliwińska, E.B.; Skórka, P.; Nowicki, P.; Wantuch, M.; Vrabec, V.; Settele, J.; Woyciechowski, M. Host ant specificity of Large Blue butterflies Phengaris (Maculinea) (Lepidoptera: Lycaenidae) inhabiting humid grasslands in East-Central Europe. Eur. J. Entomol. 2008, 105, 871–877. [Google Scholar] [CrossRef]
- Nowicki, P.; Bonelli, S.; Barbero, F.; Balletto, E. Relative importance of density-dependent regulation and environmental stochasticity for butterfly population dynamics. Oecologia 2009, 161, 227–239. [Google Scholar] [CrossRef]
- Kajzer-Bonk, J.; Nowicki, P.; Bonk, M.; Skórka, P.; Witek, M.; Woyciechowski, M. Local populations of endangered Maculinea (Maculinea) butterflies are flood resistant. J. Insect Conserv. 2013, 17, 1105–1112. [Google Scholar] [CrossRef]
- Batáry, P.; Örvössy, N.; Körösi, Á.; Vályinagy, M.; Peregovits, L. Microhabitat preferences of Maculinea teleius (Lepidoptera: Lycaenidae) in a mosaic landscape. Eur. J. Entomol. 2007, 104, 731–736. [Google Scholar] [CrossRef]
- Pedersen, J.S.; Boomsma, J.J. Genetic analysis of colony structure in polydomous and polygynous ant populations. Biol. J. Linn. Soc. 1999, 66, 115–144. [Google Scholar] [CrossRef]
- Elmes, G.W.; Thomas, J.A.; Wardlaw, J.C.; Hochberg, M.E.; Clark, R.T.; Simcox, D.J. The ecology of Myrmica ants in relation to the conservation of Maculinea butterflies. J. Insect Conserv. 1998, 2, 67–78. [Google Scholar] [CrossRef]






{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Site | Ant Species | No. and (%) of Nests | No. of Infested Nests and (Number of Larvae) | ||
|---|---|---|---|---|---|
| Autumn | Late Spring | Autumn | Late Spring | ||
| Kosyń | My. scabrinodis | 45 (79%) | 41 (68%) | 19T (48), 13M (61) | 4T (6), 3M (4) |
| My. rubra | 10 (18%) | 10 (15%) | 3T (9), 3N (6), 3M (15) | 3T (62), 2N (36) | |
| My. gallienii | 2 (3%) | 9 (17%) | 1T (3) | 2T (3), 1N (1) | |
| Kraków | My. scabrinodis | 33 (73%) | 33 (67%) | 15T (49), 2A (7), 3M (5) | 12T (24), 1A (6), 9M (22) |
| My. ruginodis | 10 (22%) | 15 (31%) | 7T (17) | 8T (15) | |
| My. rubra | 2 (5%) | 1 (2%) | 1T (3), 1N (1) | 1T (1) | |
| Ma. teleius Presence | ||
|---|---|---|
| Model Structure | Autumn | Late Spring |
| Ant species × Other parasites + Site + Other parasites × Nest size | 146.1 | 109.7 |
| Ant species × Other parasites + Site + Nest size | 145.5 | 108.0 |
| Ant species + Site + Nest size × Other parasites | 144.2 | 110.1 |
| Ant species + Other parasites + Nest size + Site | 143.5 | 108.9 |
| Ant species × Other parasites + Nest size × Other parasites | 145.3 | 117.5 |
| Ant species + Nest size × Other parasites | 143.3 | 116.6 |
| Nest size + Site + Ant species | 142.1 | 115.8 |
| Nest size + Site | 140.2 | 116.0 |
| Nest size + Ant species | 141.0 | 125.7 |
| Nest size + Site + Other parasites | 141.6 | 110.9 |
| Ant species × Other parasites | 146.6 | 118.2 |
| Nest size × Other parasites | 141.4 | 119.1 |
| Nest size | 139.0 | 126.0 |
| Ant species | 144.1 | 128.4 |
| Site | 143.2 | 121.7 |
| Other parasites | 143.8 | 122.0 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Witek, M.; La Morgia, V.; Casacci, L.P.; Barbero, F. The Pattern of Social Parasitism in Maculinea teleius Butterfly Is Driven by the Size and Spatial Distribution of the Host Ant Nests. Insects 2023, 14, 180. https://doi.org/10.3390/insects14020180
Witek M, La Morgia V, Casacci LP, Barbero F. The Pattern of Social Parasitism in Maculinea teleius Butterfly Is Driven by the Size and Spatial Distribution of the Host Ant Nests. Insects. 2023; 14(2):180. https://doi.org/10.3390/insects14020180
Chicago/Turabian StyleWitek, Magdalena, Valentina La Morgia, Luca Pietro Casacci, and Francesca Barbero. 2023. "The Pattern of Social Parasitism in Maculinea teleius Butterfly Is Driven by the Size and Spatial Distribution of the Host Ant Nests" Insects 14, no. 2: 180. https://doi.org/10.3390/insects14020180