
Managing Practical Resistance of Lepidopteran Pests to Bt Cotton in China

Abstract
:Simple Summary
Abstract
1. Introduction
2. Bt Cotton Planting and the Target Pest Resistance Management Strategy in China

{kind=link}
{kind=link}
| Insects | Resistance Mechanism | Inheritance | Reference | ||
|---|---|---|---|---|---|
| Mutations | Genes | Frequency (%) | |||
| H. armigera | Protease | Haserpin-e (up) | / 2 | / | [36] |
| HaTryR (down) | / | / | [37] | ||
| Cadherin 1 | HaCad (down) | 8.7% | Recessive | [24,31] | |
| Tetraspanin 1 | HaTSPAN1 (down) | 10% | Dominant | [38] | |
| ATP-binding cassette transporters (ABCC2/3) 1 | HaABCC2/3 (down) | / | Recessive | [39] | |
| Aminopeptidase N (HaAPN1-96S) | apn1 | / | / | [40] | |
| Alkaline phosphatase (HaALP1f) | HaALP1/2 | / | Matrilineal | [41] | |
| P. gossypiella | Cadherin 1 | r1/2PgCad1 | / | Recessive | [42] |
| r13PgCad1 | / | Recessive | [43] | ||
| r14PgCad1 | / | Recessive | [44] | ||
| r15PgCad1 | / | Recessive | [45] | ||
| r16PgCad1 | / | Recessive | [46] | ||
| r17/18PgCad1 | / | Recessive | [42] | ||
| r19/20PgCad1 | / | Recessive | [47] | ||
| PgCad189/88 | 0.02/3% | Recessive | [48] | ||
| PgCad47 | 0.03% | Recessive | [48] | ||
| PgCad65 | / | Recessive | [48] | ||
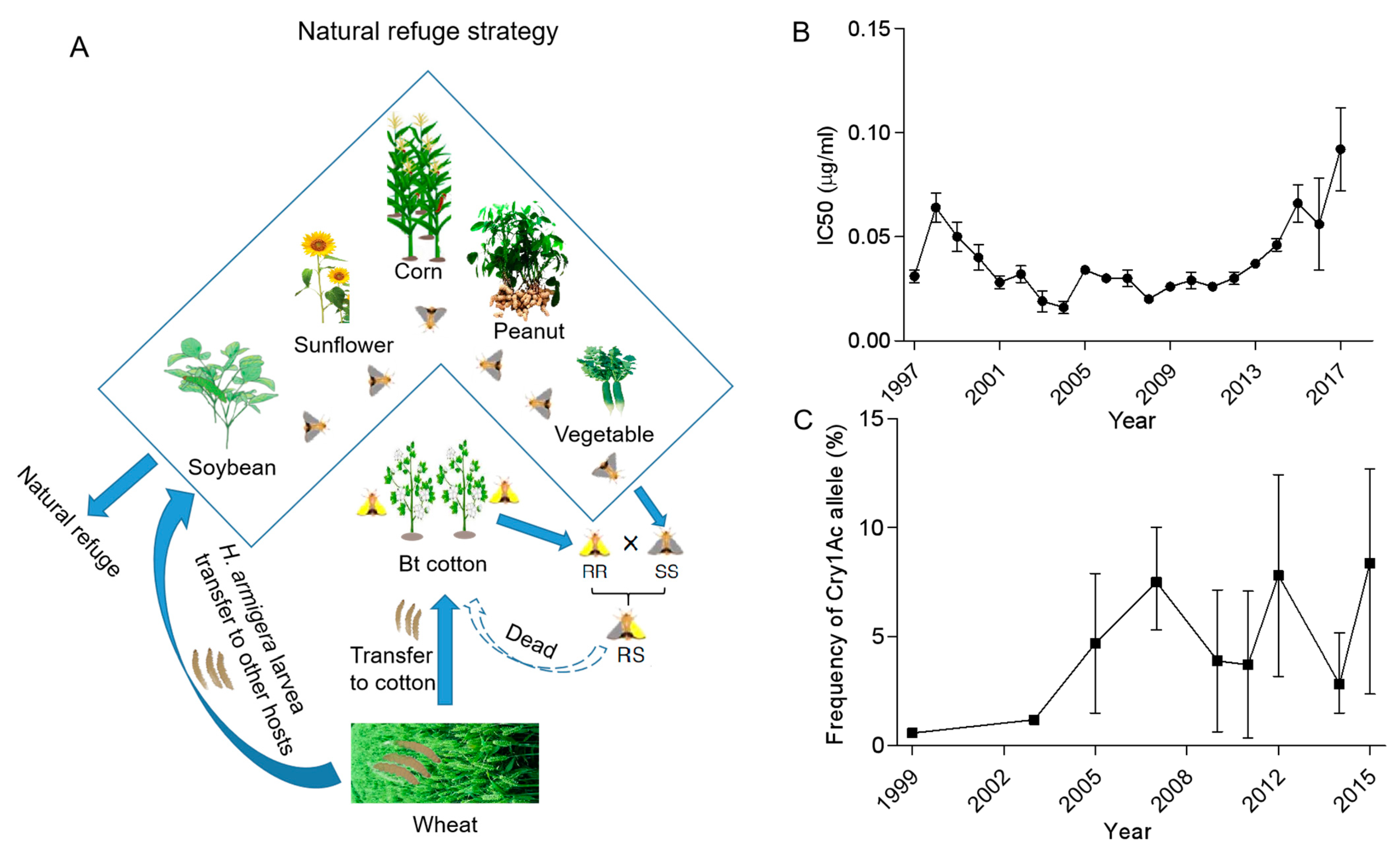
3. Resistance Management Tactic Type I: Natural Refuge for Polyphagous Cotton Bollworm

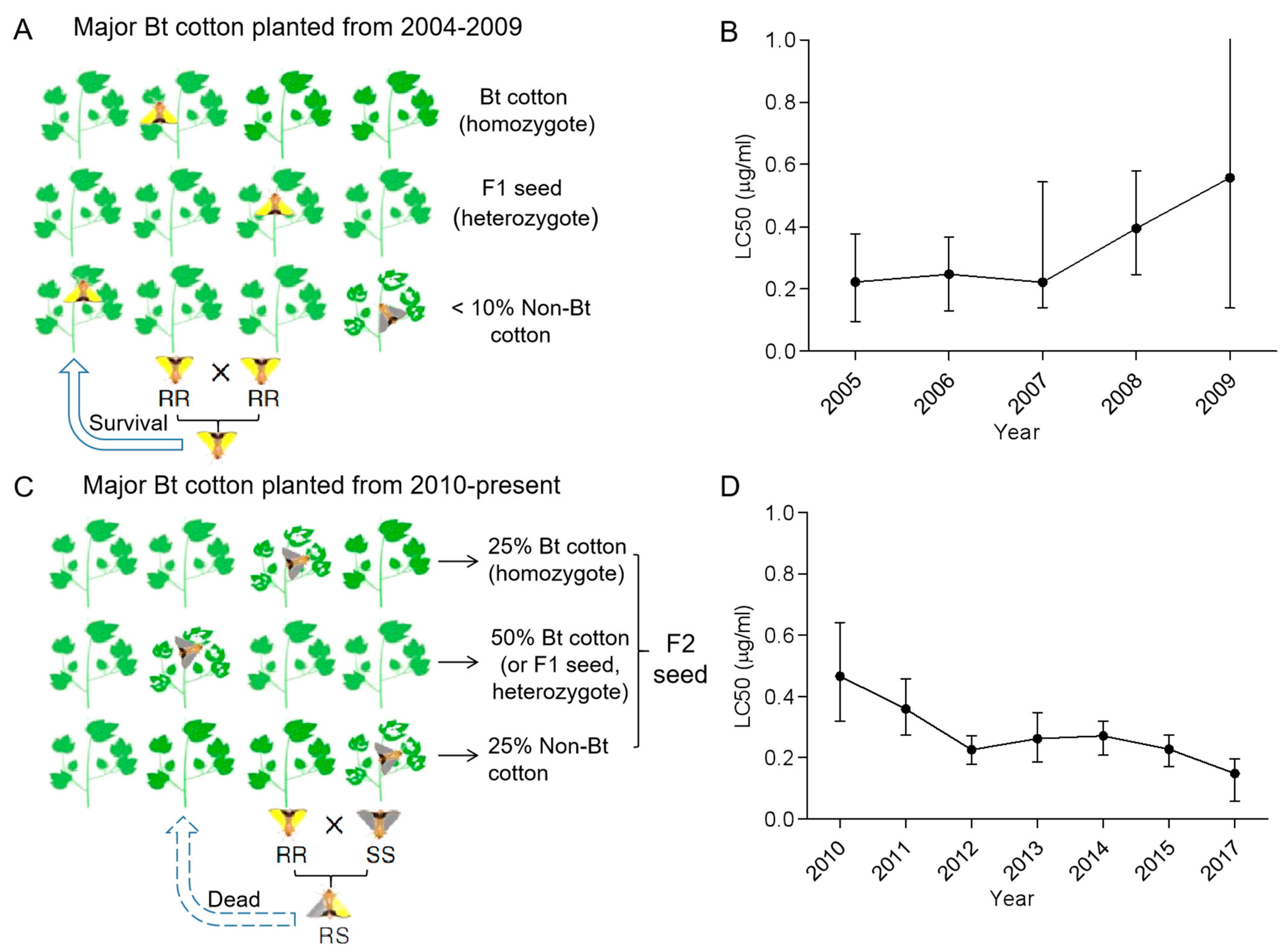
4. Resistance Management Tactic Type II: Seed Mix Refuge for Oligophagic Pink Bollworm
5. Challenges and Directions of Cotton Pest Resistance Management Strategies in China
Author Contributions
Funding
Institutional Review Board Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Dohlman, E.; Johnson, J.; MacDonald, S.; Meyer, L.; Soley, G. The World and United States Cotton Outlook. In Proceedings of the USDA’s 95 Annual Agricultural Outlook Forum, Arlington, VA, USA, 21–22 February 2019; pp. 21–22. [Google Scholar]
- Zafar, M.M.; Razzaq, A.; Farooq, M.A.; Rehman, A.; Firdous, H.; Shakeel, A.; Mo, H.; Ren, M. Insect resistance management in Bacillus thuringiensis cotton by MGPS (multiple genes pyramiding and silencing). J. Cotton Res. 2020, 3, 33. [Google Scholar] [CrossRef]
- Cui, J.; Chen, H.; Zhao, X.; Luo, J. Research course of the cotton IPM and its prospect. Cotton Sci. 2007, 19, 385–390. [Google Scholar]
- Chen, P.; Xiao, Q.; Zhang, J.; Xie, C.; Wang, B. Occurrence prediction of cotton pests and diseases by bidirectional long short-term memory networks with climate and atmosphere circulation. Comput. Electron. Agric. 2020, 176, 105612. [Google Scholar] [CrossRef]
- Tarazi, R.; Jimenez, J.L.S.; Vaslin, M.F. Biotechnological solutions for major cotton (Gossypium hirsutum) pathogens and pests. Biotechnol. Res. Innov. 2019, 3, 19–26. [Google Scholar] [CrossRef]
- Sanahuja, G.; Banakar, R.; Twyman, R.M.; Capell, T.; Christou, P. Bacillus thuringiensis: A century of research, development and commercial applications. Plant Biotechnol. J. 2011, 9, 283–300. [Google Scholar] [CrossRef]
- Luo, S.; Naranjo, S.E.; Wu, K. Biological control of cotton pests in China. Biol. Control 2014, 68, 6–14. [Google Scholar] [CrossRef]
- Wu, K.M.; Guo, Y.Y. The evolution of cotton pest management practices in China. Annu. Rev. Entomol. 2005, 50, 31–52. [Google Scholar] [CrossRef]
- Vaeck, M.; Reynaerts, A.; Hofte, H.; Jansens, S.; De Beuckeleer, M.; Dean, C.; Zabean, M.; Van Montagu, M.; Leemans, J. Transgenic plants protected from insect attack. Nature 1987, 328, 33–37. [Google Scholar] [CrossRef]
- Perlak, F.; Deaton, R.; Armstrong, T.; Fuchs, R.; Sims, S.; Greenplate, J.; Fischhoff, D. Insect resistant cotton plants. Nat. Biotechnol. 1990, 8, 939–943. [Google Scholar] [CrossRef]
- Benedict, J.H.; Ring, D.R. Transgenic crops expressing Bt proteins: Current status, challenges and outlook. In Transgenic Crop Protection: Concepts and Strategies; Koul, O., Dhaliwal, G.S., Eds.; Science Publishers: Enfield, NH, USA, 2004; pp. 15–84. [Google Scholar]
- James, C. Global Status of Commercialized Biotech/GM Crops: 2007; ISAAA Brief No. 37; International Service for the Acquisition of Agri-Biotech Applications: Ithaca, NY, USA, 2007; Available online: https://www.cambridge.org/core/journals/experimental-agriculture/article/global-status-of-commercialized-biotechgm-crops-2007-brief-37-by-c-james-ithaca-ny-usa-international-service-for-the-acquisition-of-agribiotech-applications-2007-pp-125-15-pp-executive-summary-us5000-free-to-eligible-nationals-of-developing-countries-isbn-9781892456427/0DF80AF3B90031224922AC266BE0BE94 (accessed on 20 December 2022).
- ISAAA. Gobal Status or Commercialized Biotech/GM Crops in 2019: Biotech Crops Drive Socio-Economic Development and Sustainable Environment in the New Frontier; International Service for the Acquisition of Agri-Biotech Applications: Ithaca, NY, USA, 2019. [Google Scholar]
- Tabashnik, B.E.; Carrière, Y. Global patterns of resistance to Bt crops highlighting pink bollworm in the United States, China, and India. J. Econ. Entomol. 2019, 112, 2513–2523. [Google Scholar] [CrossRef]
- Li, Y.; Eric, H.; Wu, K.; Peng, Y. Insect-resistant genetically engineered crops in China: Development, application, and prospects for use. Annu. Rev. Entomol. 2019, 65, 1–20. [Google Scholar] [CrossRef] [Green Version]
- Knight, K.M.; Head, G.P.; Rogers, D.J. Successful development and implementation of a practical proactive resistance management plan for Bt cotton in Australia. Pest Manag. Sci. 2021, 77, 4262–4273. [Google Scholar] [CrossRef]
- Bates, S.; Zhao, J.Z.; Roush, R.; Shelton, A.M. Insect resistance management in GM crops: Past, present and future. Nat. Biotechnol. 2005, 23, 57–62. [Google Scholar] [CrossRef]
- Wu, K.M.; Lu, Y.H.; Feng, H.Q.; Jiang, Y.Y.; Zhao, J.Z. Suppression of cotton bollworm in multiple crops in China in areas with Bt toxin containing cotton. Science 2008, 321, 1676–1678. [Google Scholar] [CrossRef]
- Feng, L.; Chi, B.; Dong, H. Cotton cultivation technology with Chinese characteristics has driven the 70-year development of cotton production in China. J. Integr. Agric. 2022, 21, 597–609. [Google Scholar] [CrossRef]
- Qiao, F. Fifteen years of Bt cotton in China: The economic impact and its dynamics. World Dev. 2015, 70, 177–185. [Google Scholar] [CrossRef]
- Guo, S. Engineering of insect-resistant plant with Bacillus thuringiensis crystal protein genes. Sci. Agric. Sin. 1995, 28, 8–13. (In Chinese) [Google Scholar]
- Yu, S.; Fan, S. Research and commercialization of national Bt cotton in China. Biobusiness 2010, 10, 35–41. (In Chinese) [Google Scholar]
- Wan, P.; Xu, D.; Cong, S.; Jiang, Y.; Huang, Y.; Wang, J.; Wu, H.; Wang, L.; Wu, K.; Carrière, Y.; et al. Hybridizing transgenic Bt cotton with non-Bt cotton counters resistance in pink bollworm. Proc. Natl. Acad. Sci. USA 2017, 114, 5413–5418. [Google Scholar] [CrossRef]
- Zhang, D.; Xiao, Y.; Chen, W.; Lu, Y.; Wu, K. Field monitoring of Helicoverpa armigera (Lepidoptera:Noctuidae) Cry1Ac insecticidal protein resistance in China (2005–2017). Pest Manag. Sci. 2019, 75, 753–759. [Google Scholar]
- Lü, L.; Luo, J.; Zhang, S.; Yu, Q.; Ma, L.; Liu, X.; Wang, C.; Ma, X.; Ma, Y.; Cui, J. Efficiency of cotton bollworm (Helicoverpa armigera Hübner) control of different Bt cotton varieties in North China. J. Cott. Res. 2018, 1, 4. [Google Scholar] [CrossRef]
- Wu, G.; Harris, M.K.; Guo, J.Y.; Wan, F.H. Response of multiple generations of beet armyworm, Spodoptera exigua (Hübner), feeding on transgenic Bt cotton. J. Appl. Entomol. 2009, 133, 90–100. [Google Scholar] [CrossRef]
- Wan, P.; Wu, K.; Huang, M.; Yu, D.; Wu, J. Population dynamics of Spodoptera litura (Lepidoptera: Noctuidae) on Bt cotton in the Yangtze River valley of China. Environ. Entomol. 2008, 37, 1043–1048. [Google Scholar] [CrossRef] [PubMed]
- Xu, W.; Liu, B.; Wang, R.; Zheng, Y.; Zhang, Y.; Li, X. Effects of transgenic Bt cotton on insect population in cotton field in coastal agricultural region of Jiangsu province. J. Ecol. Rural Environ. 2008, 24, 32–38. (In Chinses) [Google Scholar]
- Tabashnik, B.E.; Carrière, Y. Successes and Failures of Transgenic Bt Crops: Global Patterns of Field-evolved Resistance. In Bt Resistance—Characterization and Strategies for GM Crops Producing Bacillus Thuringiensis Toxins; Soberón, M., Gao, Y., Bravo, A., Eds.; CABI: Wallingford, UK, 2015; Volume 4, pp. 1–14. [Google Scholar]
- Qiao, F.; Huang, J. Sustainability of the economic benefit of Bt cotton in China: Results from household surveys. J. Dev. Stud. 2020, 56, 2045–2060. [Google Scholar] [CrossRef]
- Jurat-fuentes, J.L.; Heckel, D.G.; Ferre, J. Mechanisms of Resistance to Insecticidal Proteins from Bacillus thuringiensis. Annu. Rev. Entomol. 2020, 66, 121–140. [Google Scholar] [CrossRef]
- Gould, F.; Anderson, A.; Jones, A.; Sumerford, D.; Heckel, D.G.; Lopez, J.; Micinski, C.; Leonard, R.; Laster, M. Initial frequency of alleles for resistance to Bacillus thuringiensis toxin in field populations of Heliothis virescens. Proc. Natl. Acad. Sci. USA 1997, 94, 3519–3523. [Google Scholar] [CrossRef]
- Cerda, H.; Wright, D.J. Modeling the spatial and temporal location of refugia to manage resistance in Bt transgenic crops. Agric. Ecosyst. Environ. 2004, 102, 163–174. [Google Scholar] [CrossRef]
- Mohan, K.S.; Ravi, K.C.; Suresh, P.J.; Sumerford, D.; Head, G.P. Field resistance to the Bacillus thuringiensis protein Cry1Ac expressed in Bollgard® hybrid cotton in pink bollworm, Pectinophora gossypiella (Saunders), populations in India. Pest Manag. Sci. 2016, 72, 738–746. [Google Scholar] [CrossRef]
- Wu, K.; Guo, Y.; Gao, S. Evaluation of the natural refuge function for Helicoverpa armigera (Lepidoptera: Noctuidae) within Bacillus thuringiensis transgenic cotton growing areas in northern China. J. Econ. Entomol. 2002, 95, 832–837. [Google Scholar] [CrossRef]
- Zhang, C.; Wei, J.; Naing, Z.L.; Soe, E.T.; Tang, J.; Liang, G. Up-regulated serpin gene involved in Cry1Ac resistance in Helicoverpa armigera. Pestic. Biochem. Phys. 2022, 188, 105269. [Google Scholar] [CrossRef]
- Liu, C.; Xiao, Y.; Li, X.; Oppert, B.; Tabashnik, B.E.; Wu, K.L. Cis-mediated down-regulation of a trypsin gene associated with Bt resistance in cotton bollworm. Sci. Rep. 2014, 4, 7219. [Google Scholar] [CrossRef]
- Jin, L.; Wang, J.; Guan, F.; Zhang, J.; Yu, S.; Liu, S.; Xue, Y.; Li, L.; Wu, S.; Wang, X.; et al. Dominant point mutation in a tetraspanin gene associated with field-evolved resistance of cotton bollwrom to transgenic Bt cotton. Proc. Natl. Acad. Sci. USA 2018, 115, 11760–11765. [Google Scholar] [CrossRef]
- Wang, J.; Ma, H.; Zhao, S.; Huang, J.; Yang, Y.; Tabashnik, B.E.; Wu, Y. Functional redundancy of two ABC transporter proteins in mediating toxicity of Bacillus thuringiensis to cotton bollworm. PLoS Pathog. 2021, 16, e1008427. [Google Scholar] [CrossRef]
- Zhang, S.; Cheng, H.; Gao, Y.; Wang, G.; Liang, G.; Wu, K. Mutation of an aminopeptidase N gene is associated with Helicoverpa armigera resistance to Bacillus thuringiensis Cry1Ac toxin. Insect Biochem. Molec. 2009, 39, 421–429. [Google Scholar] [CrossRef]
- Ningthoujam, K.; Sorokhaibam, M.; Thangjam, R.; Gujar, G.T. Association of alkaline phosphatase activity with inheritance of Cry1Ac resistance in Helicoverpa armigera (Hübner). J. Entomol. Zool. Stud. 2020, 8, 198–204. [Google Scholar]
- Wang, J.; Xu, D.; Wang, L.; Cong, S.; Wan, P.; Lei, C.; Fabrick, J.A.; Li, X.; Tabashnik, B.E.; Wu, K. Bt resistance alleles in field populations of pink bollworm from China: Similarities with the United States and decreased frequency from 2012 to 2015. Pest Manag. Sci. 2020, 76, 527–533. [Google Scholar] [CrossRef]
- Wang, L.; Ma, Y.; Wan, P.; Liu, K.; Xiao, Y.; Wang, J.; Cong, S.; Xu, D.; Wu, K.; Fabrick, J.A.; et al. Resistance to Bacillus thuringiensis linked with a cadherin transmembrane mutation affecting cellular trafficking in pink bollworm from China. Insect Biochem. Mol. Biol. 2018, 94, 28–35. [Google Scholar] [CrossRef]
- Wang, L.; Ma, Y.; Wei, W.; Wan, P.; Liu, K.; Xu, M.; Cong, S.; Wang, J.; Xu, D.; Xiao, Y.; et al. Cadherin repeat 5 mutation associated with Bt resistance in a field-derived strain of pink bollworm. Sci. Rep. 2020, 10, 16840. [Google Scholar] [CrossRef]
- Wang, L.; Wang, J.; Ma, Y.; Wan, P.; Liu, K.; Cong, S.; Xiao, Y.; Xu, D.; Wu, K.; Fabrick, J.A.; et al. Transposon insertion causes cadherin mis-splicing and confers resistance to Bt cotton in pink bollworm from China. Sci. Rep. 2019, 9, 7479. [Google Scholar] [CrossRef]
- Wang, L.; Ma, Y.; Guo, X.; Wan, P.; Liu, K.; Cong, S.; Wang, J.; Xu, D.; Xiao, Y.; Li, X.; et al. Pink bollworm resistance to Bt toxin Cry1Ac associated with an insertion in cadherin exon 20. Toxins 2019, 11, 186. [Google Scholar] [CrossRef] [PubMed]
- Wang, L.; Xu, D.; Huang, Y.; Zhou, H.; Liu, W.; Cong, S.; Wang, J.; Li, W.; Wan, P. Mutation in the cadherin gene is a key factor for pink bollworm resistance to Bt cotton in China. Toxins 2022, 14, 23. [Google Scholar] [CrossRef] [PubMed]
- Wang, L. Identification and Analysis of Resistance Alleles of Pink Bollworm Cadherin in the Yangtze River Valley. Doctorate Thesis, Chinese Academy of Agriculture Sciences, Beijing, China, 2017. (In Chinese). [Google Scholar]
- Chen, R.; Huang, J.; Qiao, F. Farmers’ knowledge on pest management and pesticide use in Bt cotton production in China. China Econ. Rev. 2013, 27, 15–24. [Google Scholar] [CrossRef]
- Huang, J.; Hu, R.; van Meijl, H.; van Tongeren, F. Economic impacts of genetically modified crops in China. Genet. Mol. Biol. 2003, 24, 18390. [Google Scholar]
- Feng, H.Q.; Wu, K.M.; Cheng, D.F.; Guo, Y.Y. Northward migration of Helicoverpa armigera (Lepidoptera: Noctuidae) and other moths in early summer observed with radar in northern China. J. Econ. Entomol. 2004, 97, 1874–1883. [Google Scholar] [CrossRef]
- Feng, H.; Wu, K.; Ni, Y.; Cheng, D.; Guo, Y. Return migration of Helicoverpa armigera (Lepidoptera: Noctuidae) during autumn in northern China. B. Entomol. Res. 2005, 95, 361–370. [Google Scholar] [CrossRef]
- Cheng, D.; Wu, K.; Tian, Z.; Wen, L.; Shen, Z. Acquisition and analysis of migration data from the digitized display of a scanning entomological radar. Comput. Electron. Agric. 2020, 35, 63–75. [Google Scholar] [CrossRef]
- Wu, K.; Guo, Y.; Wu, Y. Ovarian development of adult females of cotton bollworm and its relation to migratory behavior around Bohai Bay of China. Acta Ecol. Sin. 2002, 22, 1075–1078. (In Chinese) [Google Scholar]
- Wu, K. Monitoring and management strategy for Helicoverpa armigera resistance to Bt cotton in China. J. Invertebr. Pathol. 2007, 95, 220–223. [Google Scholar] [CrossRef]
- Wu, K.; Guo, Y.; Lv, N. Geographic variation in susceptibility of Helicoverpa armigera (Lepidoptera: Noctuidae) to Bacillus thuringiensis insecticidal protein in China. J. Econ. Entomol. 1999, 92, 273–278. [Google Scholar] [CrossRef]
- Wu, K.; Guo, Y.; Lv, N.; Greenplate, J.T.; Deaton, R. Resistance monitoring of Helicoverpa armigera (Lepidoptera: Noctuidae) to Bacillus thuringiensis insecticidal protein in China. J. Econ. Entomol. 2002, 95, 826–831. [Google Scholar] [CrossRef]
- Liu, F.; Xu, Z.; Chang, J.; Chen, J.; Meng, F.; Zhu, Y.C.; Shen, J. Resistance allele frequency to Bt Cotton in field populations of Helicoverpa armigera (Lepidoptera: Noctuidae) in China. J. Econ. Entomol. 2008, 101, 933–943. [Google Scholar] [CrossRef]
- Li, G.; Wu, K.; Gould, F.; Feng, H.; He, Y.; Guo, Y. Frequency of Bt resistance genes in Helicoverpa armigera populations from the Yellow River cotton-farming region of China. Entomol. Exp. Appl. 2004, 112, 135–143. [Google Scholar] [CrossRef]
- Pan, L.; Shi, M.; Chen, J.; Wei, Q.; Gao, C. Resistance monitoring of larvae treated with Bt cotton and pesticides in Helicoverpa armigera (Lepidoptera: Noctuidae). Orient. Insects 2017, 51, 285–296. [Google Scholar] [CrossRef]
- Li, G.P.; Feng, H.Q.; Gao, Y.L.; Wyckhuys, K.A.C.; Wu, K.M. Frequency of Bt resistance alleles in Helicoverpa armigera in the Xinjiang cotton-planting region of China. Environ. Entomol. 2010, 39, 1698–1704. [Google Scholar] [CrossRef]
- Jin, L.; Zhang, H.; Lu, Y.; Yang, Y.; Wu, K.; Tabashnik, B.E.; Wu, Y. Large-scale test of the natural refuge strategy for delaying insect resistance to transgenic Bt crops. Nat. Biotechnol. 2015, 33, 169–174. [Google Scholar] [CrossRef]
- Li, Y.; Gao, Y.; Wu, K. Function and effectiveness of natural refuge in IRM strategies for Bt crops. Curr. Opin. Insect Sci. 2017, 21, 1–6. [Google Scholar] [CrossRef]
- Zhao, J.; Cao, J.; Li, Y.; Collins, H.L.; Roush, R.T.; Earle, E.D.; Shelton, A.M. Transgenic plants expressing two Bacillus thuringiensis toxins delay insect resistance evolution. Nat. Biotechnol. 2003, 21, 1493–1497. [Google Scholar] [CrossRef]
- Brevault, T.; Heuberger, S.; Zhang, M.; Ellers-Kirk, C.; Ni, X.; Masson, L.; Li, X.; Tabashnik, B.E.; Carrière, Y. Potential shortfall of pyramided transgenic cotton for insect resistance management. Proc. Natl. Acad. Sci. USA 2013, 110, 5806–5811. [Google Scholar] [CrossRef]
- Liu, Y.D.; Wu, K.M.; Guo, Y.Y. Population structure and introduction history of the pink bollworm, Pectinophora gossypiella, in China. Entomol. Exp. Appl. 2009, 130, 160–172. [Google Scholar] [CrossRef]
- Centre for Agriculture and Biosciences International (CABI). Invasive Species Compendium: Datasheet Report for Pectinophora gossypiella (Pink Bollworm). 2016. Available online: www.cabi.org/isc/datasheet/39417 (accessed on 20 December 2022).
- Ariela, N.; Liora, S.H.; Mario, R.; Roee, S.; Rami, H.A. Can tomato be a potential host plant for pink bollworm? World Cott. Res. 2011, 42, 258–260. [Google Scholar]
- Tabashnik, B.E.; Morin, S.; Unnithan, G.C.; Yelich, A.J.; Ellers-Kirk, C.; Harpold, V.S.; Sisterson, M.S.; Ellsworth, P.C.; Dennehy, T.J.; Antilla, L.; et al. Sustained susceptibility of pink bollworm to Bt cotton in the United States. GM Crops Food 2012, 3, 194–200. [Google Scholar] [CrossRef] [PubMed]
- Grefenstette, B.; El-Lissy, O.; Staten, R.T. Pink Bollworm Eradication Plan in the U.S. 2009. Available online: https://citeseerx.ist.psu.edu/document?repid=rep1&type=pdf&doi=94468d9d02b6c89992db5fd47d5024a3070a81ce (accessed on 10 January 2023).
- Tabashnik, B.E.; Sisterson, M.S.; Ellsworth, P.C.; Dennehy, T.J.; Antilla, L.; Liesner, L.; Whitlow, M.; Staten, R.T.; Fabrick, J.A.; Unnithan, G.C.; et al. Suppressing resistance to Bt cotton with sterile insect releases. Nat. Biotechnol. 2010, 28, 1304–1307. [Google Scholar] [CrossRef] [PubMed]
- Liesner, L.; Antilla, L.; Whitlow, M.; Staten, R.T. Arizona pink bollworm eradication program 2010 update. In Proceedings of the 2011 Beltwide Cotton Conferences, National Cotton Council of America, Memphis, TN, USA, 4–7 January 2011; pp. 1143–1149. [Google Scholar]
- Liesner, L.R.; Fairchild, D.; Solis, J. Arizona pink bollworm eradication program 2017 update. In Proceedings of the Beltwide Cotton Conferences, National Cotton Council of America, Memphis, TN, USA, 3–5 January 2018; pp. 718–720. [Google Scholar]
- Liesner, L.R.; Fairchild, D.; Brengle, J. Arizona pink bollworm eradication program 2018 update. In Proceedings of the Beltwide Cotton Conferences, National Cotton Council of America, Memphis, TN, USA, 8–10 January 2019; pp. 371–373. [Google Scholar]
- Persue, S. Eradication of Pink Bollworm Proclamation. 2018. Available online: https://www.usda.gov/sites/default/files/documents/usda-pink-bollworm-proclamation.pdf (accessed on 10 January 2023).
- Mohan, K.S. Refuge-in-bag for Bt cotton. Curr. Sci. 2018, 114, 726–727. [Google Scholar]
- Naik, V.C.; Kumbhare, S.; Kranthi, S.; Satija, U.; Kranthi, K.R. Field-evolved resistance of pink bollworm, Pectinophora gossypiella (Saunders)(Lepidoptera: Gelechiidae), to transgenic Bacillus thuringiensis (Bt) cotton expressing crystal 1Ac (Cry1Ac) and Cry2Ab in India. Pest Manag. Sci. 2018, 74, 2544–2554. [Google Scholar] [CrossRef]
- Kukanar, V.S.; Singh, T.V.K.; Kranthi, K.R.; Andow, D.A. Cry1Acresistance allele frequency in field populations of Helicoverpa armigera (Hübner) collected in Telangana and Andhra Pradesh, India. Crop Prot. 2018, 107, 34–40. [Google Scholar] [CrossRef]
- Carroll, M.W.; Head, G.; Caprio, M. When and where a seed mix refuge makes sense for managing insect resistance to Bt plants. Crop Prot. 2012, 38, 74–79. [Google Scholar] [CrossRef]
- Tabashnik, B.E.; Wu, K.; Wu, Y. Early detection of field-evolved resistance to Bt cotton in China: Cotton bollworm and pink bollworm. J. Invertebr. Pathol. 2012, 110, 301–306. [Google Scholar] [CrossRef]
- Dong, H.; Li, W.; Tang, W.; Zhang, D. Development of hybrid Bt cotton in China—A successful integration of transgenic technology and conventional techniques. Curr. Sci. 2004, 86, 778–782. [Google Scholar]
- Feng, G.; Gan, X.; Yao, Y.; Luo, H.; Zhang, Y.; Zhang, W. Comparisons of photosynthetic characteristics in relation to lint yield among F1 hybrids, their F2 descendants and parental lines of cotton. J. Integr. Agric. 2014, 13, 1909–1920. [Google Scholar] [CrossRef]
- Zhang, B.H.; Guo, T.; Wang, Q. Inheritance and segregation of exogenous genes in transgenic cotton. J. Genet. 2000, 79, 71–75. [Google Scholar] [CrossRef]
- Heuberger, S.; Ellers-Kirk, C.; Yafuso, C.; Grassmann, A.J.; Tabashnik, B.E.; Dennehy, T.J.; Carrière, Y. Effects of refuge contamination by transgenes on Bt resistancein pink bollworm (Lepidoptera: Gelechiidae). J. Econ. Entomol. 2008, 101, 504–514. [Google Scholar] [CrossRef]
- Wan, P.; Huang, Y.; Wu, H.; Huang, M.; Cong, S.; Tabashnik, B.E.; Wu, K. Increased frequency of pink bollworm resistance to Bt toxinCry1Ac in China. PLoS ONE 2012, 7, e29975. [Google Scholar]
- MARA. The Release of National GM Soybean/Corn Variety Certification Standard (Try Out). 2022. Available online: http://www.fgs.moa.gov.cn/flfg/202206/t20220608_6401924.htm (accessed on 20 December 2022).
- Yang, X.; Zhao, S.; Liu, B.; Gao, Y.; Hu, C.; Li, W.; Yang, Y.; Li, G.; Wang, L.; Yang, X.; et al. Bt maize can provide non-chemical pest control and enhance food safety in China. Plant Biotechnol. J. 2022, 21, 391–404. [Google Scholar] [CrossRef]
- Liang, J.; Yang, X.; Jiao, Y.; Wang, D.; Zhao, Q.; Sun, Y.; Li, Y.; Wu, K. The evolution of China’s regulation of agricultural biotechnology. Abiotech 2022, 3, 237–249. [Google Scholar] [CrossRef]
- Hutchison, W.D.; Burkness, E.C.; Mitchell, P.D.; Moon, R.D.; Leslie, T.W.; Fleischer, S.J.; Abrahamson, M.; Hamilton, K.L.; Steffey, K.L.; Gray, M.E.; et al. Areawide suppression of European corn borer with Bt maize reaps savings to non-Bt maize growers. Science 2010, 330, 222–225. [Google Scholar] [CrossRef]
- Dively, G.P.; Venugopal, P.D.; Bean, D.; Whalen, J.; Holmstrom, K.; Kuhar, T.P.; Doughty, H.B.; Patton, T.; Cissel, W.; Hutchison, W.D. Regional pest suppression associated with widespread Bt maize adoption benefits vegetable growers. Proc. Natl. Acad. Sci. USA 2018, 115, 3320–3325. [Google Scholar] [CrossRef]
- Roush, R.T. Bt-transgenic crops: Just another pretty insecticide or a chance for a new start in resistance management? Pestic. Sci. 1997, 51, 328–334. [Google Scholar] [CrossRef]
- Tabashnik, B.E.; Carrière, Y. Surge in insect resistance to transgenic crops and prospects for sustainability. Nat. Biotechnol. 2017, 35, 926–935. [Google Scholar] [CrossRef]

Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Quan, Y.; Wu, K. Managing Practical Resistance of Lepidopteran Pests to Bt Cotton in China. Insects 2023, 14, 179. https://doi.org/10.3390/insects14020179
Quan Y, Wu K. Managing Practical Resistance of Lepidopteran Pests to Bt Cotton in China. Insects. 2023; 14(2):179. https://doi.org/10.3390/insects14020179
Chicago/Turabian StyleQuan, Yudong, and Kongming Wu. 2023. "Managing Practical Resistance of Lepidopteran Pests to Bt Cotton in China" Insects 14, no. 2: 179. https://doi.org/10.3390/insects14020179