
Molecular Genetic Basis of Lab- and Field-Selected Bt Resistance in Pink Bollworm

Abstract
:Simple Summary
Abstract
1. Introduction
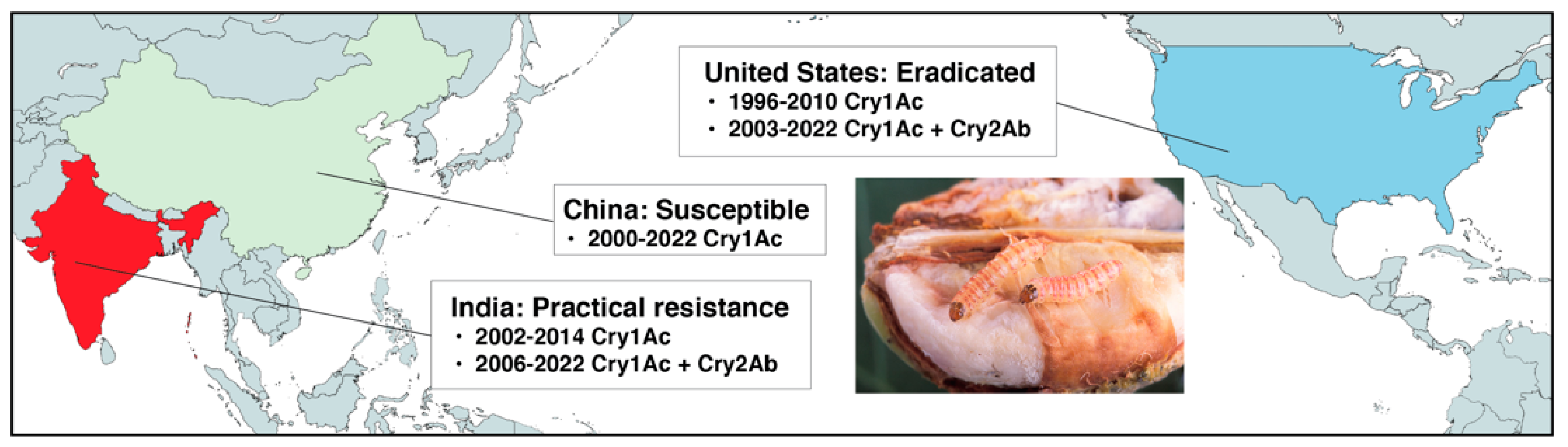
2. Field Outcomes: Pink Bollworm and Bt Cotton in the U.S., China, and India
2.1. Sustained Susceptibilty to Bt Cotton and Eradication of Pink Bollworm in the U.S.
2.2. Pink Bollworm Susceptibility to Bt Cotton Producing Cry1Ac Restored in China
2.3. Practical Resistance of Pink Bollworm to Bt Cotton in India
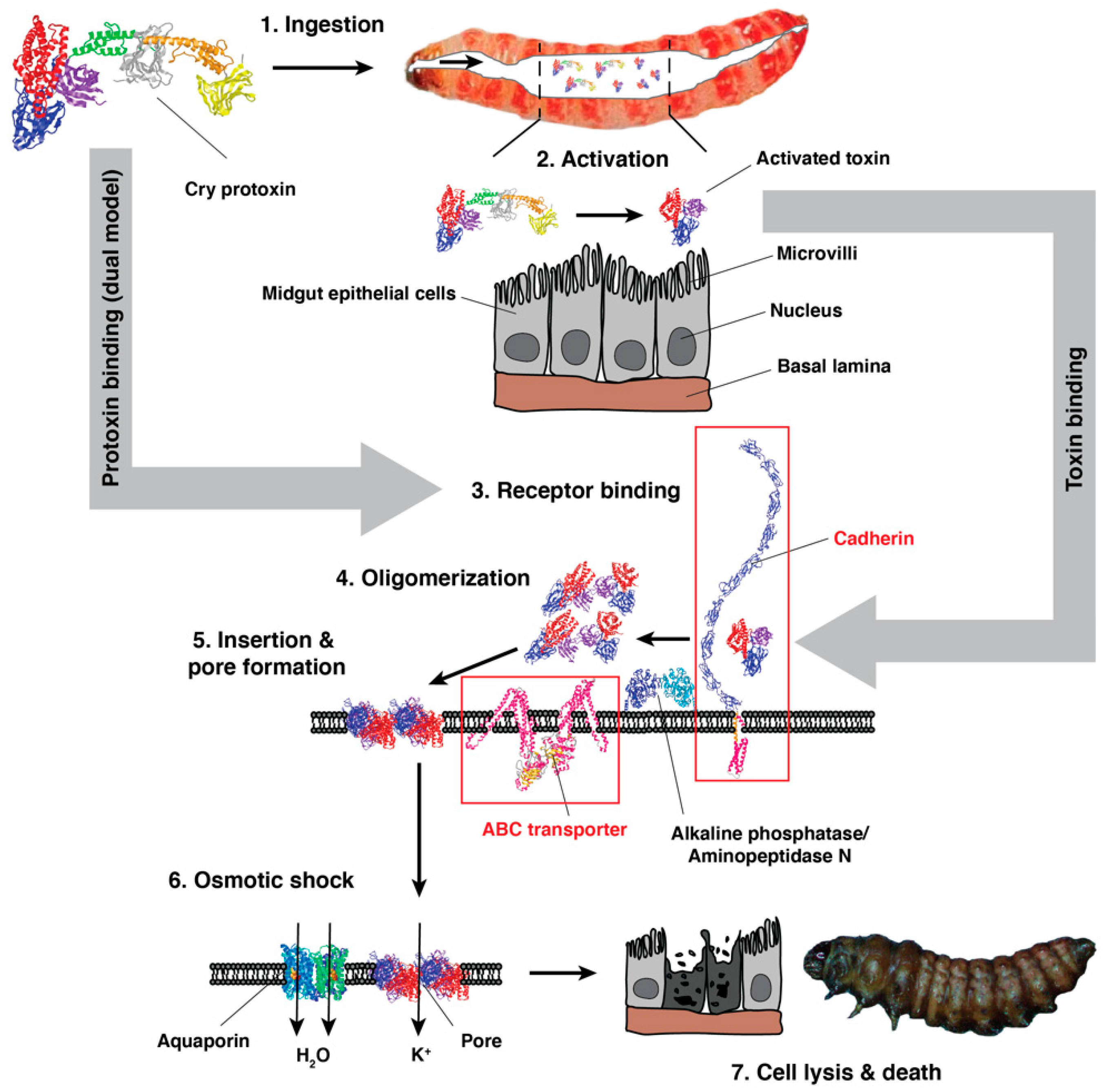
3. Cry Toxin Mode of Action
4. Pink Bollworm Resistance to Cry1Ac
4.1. Shared Mode of Resistance to Cry1Ac in the U.S., China, and India
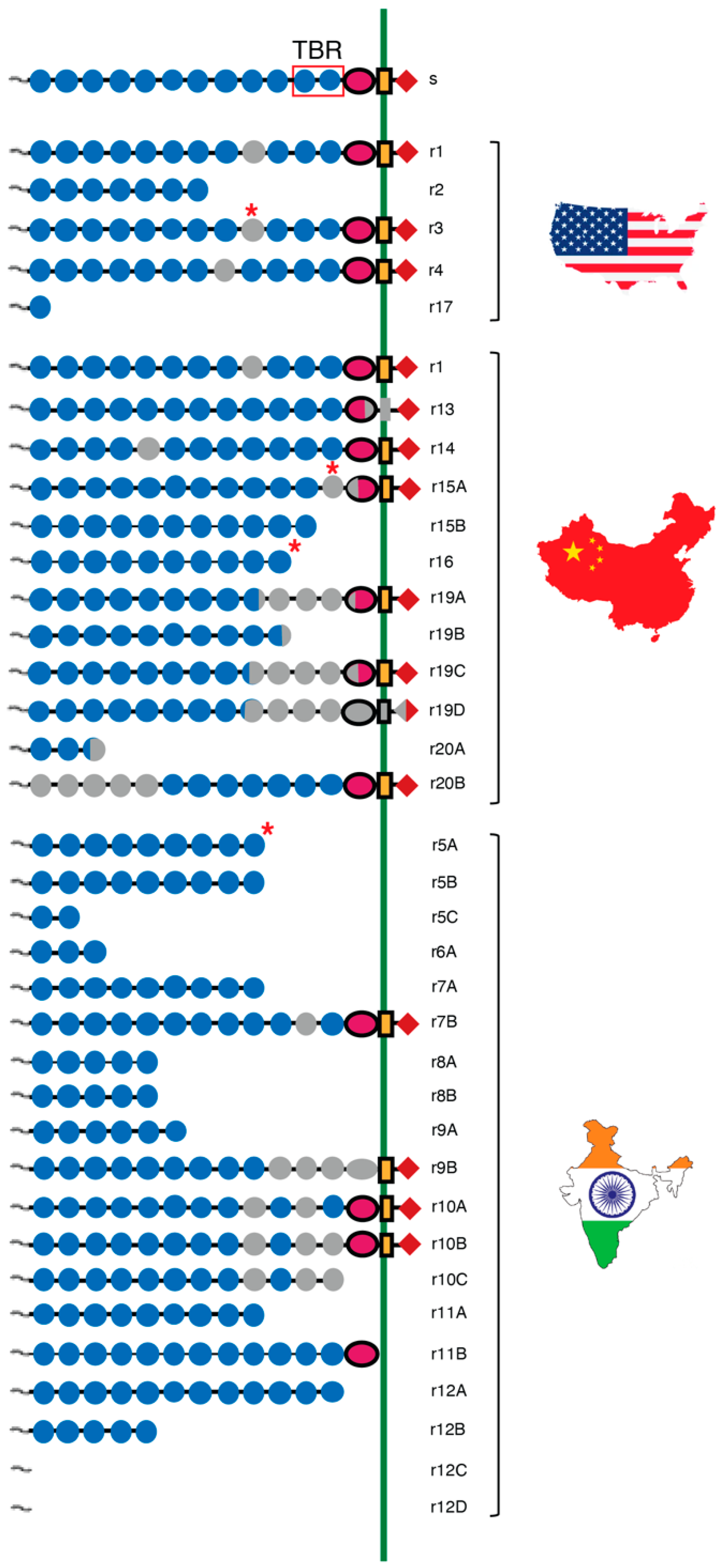
4.2. Cadherin Mutations Associated with Pink Bollworm Resistance to Cry1Ac
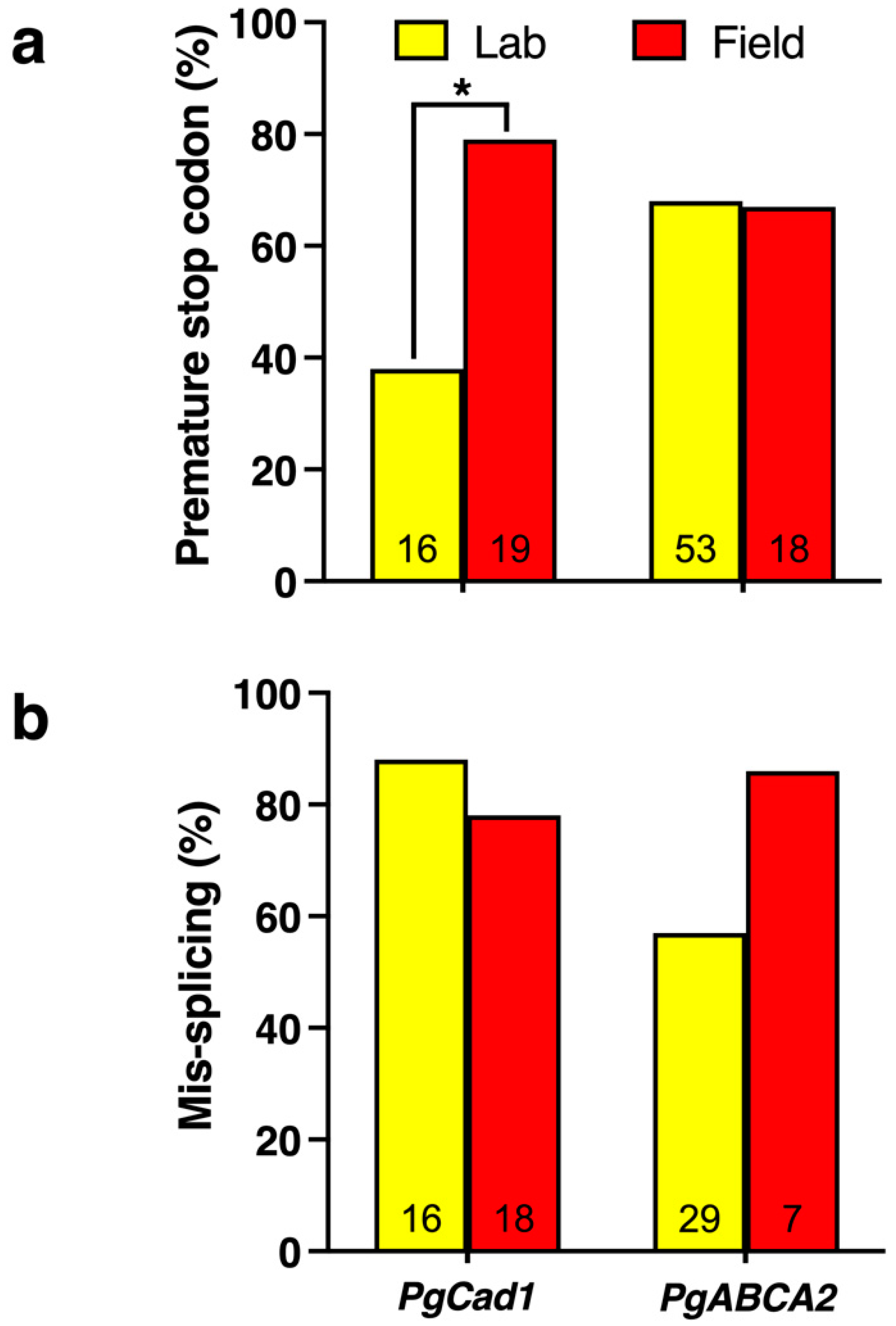
4.2.1. PgCad1 Resistance Alleles Differ between Lab- and Field-Selected Pink Bollworm
4.2.2. PgCad1 Resistance Alleles in Field Populations in the U.S., China, and India
4.2.3. PgCad1 Mutations Caused by Transposable Elements
4.2.4. Localization of PgCad1 on the Cell Membrane
4.2.5. Reduced Expression of PgCad1
5. Pink Bollworm Resistance to Cry2Ab
5.1. Modes of Resistance to Cry2Ab in India and the U.S.
5.2. ABC Transporter Mutations Associated with Pink Bollworm Resistance to Cry2Ab
5.3. PgABCA2 Mutations Introduced by CRISPR/Cas9 Gene Editing
5.4. An Alternative Mechanism of Resistance to Cry2Ab in the BX-R Strain from the U.S.
6. Conclusions
Supplementary Materials
Author Contributions
Funding
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Mendelsohn, M.; Kough, J.; Vaituzis, Z.; Matthews, K. Are Bt crops safe? Nat. Biotechnol. 2003, 21, 1003–1009. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sanahuja, G.; Banakar, R.; Twyman, R.M.; Capell, T.; Christou, P. Bacillus thuringiensis: A century of research, development and commercial applications. Plant Biotechnol. J. 2011, 9, 283–300. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Comas, C.; Lumbierres, B.; Pons, X.; Albajes, R. No effects of Bacillus thuringiensis maize on nontarget organisms in the field in southern Europe: A meta-analysis of 26 arthropod taxa. Transgenic Res. 2014, 23, 135–143. [Google Scholar] [CrossRef] [PubMed]
- Romeis, J.; Naranjo, S.E.; Meissle, M.; Shelton, A.M. Genetically engineered crops help support conservation biological control. Biol. Control 2019, 130, 136–154. [Google Scholar] [CrossRef]
- Krogh, P.H.; Kostov, K.; Damgaard, C.F. The effect of Bt crops on soil invertebrates: A systematic review and quantitative meta-analysis. Transgenic Res. 2020, 29, 487–498. [Google Scholar] [CrossRef]
- ISAAA. Global Status of Commercialized Biotech/GM Crops in 2019: Biotech Crops Drive Socio-Economic Development and Sustainable Environment in the New Frontier; ISAAA Brief No. 55; ISAAA: Ithaca, NY, USA, 2019. [Google Scholar]
- Addae, P.C.; Ishiyaku, M.F.; Tignegre, J.B.; Ba, M.N.; Bationo, J.B.; Atokple, I.D.K.; Abudulai, M.; Dabiré-Binso, C.L.; Traore, F.; Saba, M.; et al. Efficacy of a cry1Ab gene for control of Maruca vitrata (Lepidoptera: Crambidae) in cowpea (Fabales: Fabaceae). J. Econ. Entomol. 2020, 113, 974–979. [Google Scholar] [CrossRef] [Green Version]
- Shelton, A.M.; Sarwer, S.H.; Hossain, M.J.; Brookes, G.; Paranjape, V. Impact of Bt brinjal cultivation in the market value chain in five districts of Bangladesh. Front. Bioeng. Biotechnol. 2020, 8, 498. [Google Scholar] [CrossRef]
- de Oliveira, W.S.; Sakuno, C.I.R.; Miraldo, L.I.; Tavares, M.A.G.C.; Komada, K.M.A.; Teresani, D.; Santos, J.L.X.; Huang, F. Varied frequencies of resistance alleles to Cry1Ab and Cry1Ac among Brazilian populations of the sugarcane borer, Diatraea saccharalis (F.). Pest Manag. Sci. 2022, 78, 5150–5163. [Google Scholar] [CrossRef]
- Hutchison, W.D.; Burkness, E.C.; Mitchell, P.D.; Moon, R.D.; Leslie, T.W.; Fleischer, S.J.; Abrahamson, M.; Hamilton, K.L.; Steffey, K.L.; Gray, M.E.; et al. Areawide suppression of European corn borer with Bt maize reaps savings to non-Bt maize growers. Science 2010, 330, 222–225. [Google Scholar] [CrossRef] [Green Version]
- National Academies of Sciences, Engineering, and Medicine. Genetically Engineered Crops: Experiences and Prospects; The National Academies Press: Washington, WA, USA, 2016. [Google Scholar] [CrossRef]
- Dively, G.P.; Venugopal, P.D.; Bean, D.; Whalen, J.; Holmstrom, K.; Kuhar, T.P.; Doughty, H.B.; Patton, T.; Cissel, W.; Hutchison, W.D. Regional pest suppression associated with widespread Bt maize adoption benefits vegetable growers. Proc. Natl. Acad. Sci. USA 2018, 115, 3320–3325. [Google Scholar] [CrossRef] [Green Version]
- Tabashnik, B.E.; Liesner, L.R.; Ellsworth, P.C.; Unnithan, G.C.; Fabrick, J.A.; Naranjo, S.E.; Li, X.; Dennehy, T.J.; Antilla, L.; Staten, R.T.; et al. Transgenic cotton and sterile insect releases synergize eradication of pink bollworm a century after it invaded the United States. Proc. Natl. Acad. Sci. USA 2021, 118, e2019115118. [Google Scholar] [CrossRef] [PubMed]
- Tabashnik, B.E.; Fabrick, J.A.; Carrière, Y. Global patterns of insect resistance to transgenic Bt crops: The first 25 years. J. Econ. Entomol. 2023, toac183. [Google Scholar] [CrossRef] [PubMed]
- Soberón, M.; Pardo-López, L.; López, I.; Gómez, I.; Tabashnik, B.E.; Bravo, A. Engineering modified Bt toxins to counter insect resistance. Science 2007, 318, 1640–1642. [Google Scholar] [CrossRef]
- Tabashnik, B.E.; Fabrick, J.A.; Unnithan, G.C.; Yelich, A.J.; Masson, L.; Zhang, J.; Bravo, A.; Soberón, M. Efficacy of genetically modified Bt toxins alone and in combinations against pink bollworm resistant to Cry1Ac and Cry2Ab. PLoS ONE 2013, 8, e80496. [Google Scholar] [CrossRef]
- Jin, L.; Wang, J.; Guan, F.; Zhang, J.; Yu, S.; Liu, S.; Xue, Y.; Lia, L.; Wu, S.; Wang, X.; et al. Dominant point mutation in a tetraspanin gene associated with field-evolved resistance of cotton bollworm to transgenic Bt cotton. Proc. Natl. Acad. Sci. USA 2018, 115, 11760–11765. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Banerjee, R.; De Bortoli, C.P.; Huang, F.; Lamour, K.; Meagher, R.; Buntin, D.; Ni, X.; Reay-Jones, F.P.F.; Stewart, S.; Jurat-Fuentes, J.L. Large genomic deletion linked to field-evolved resistance to Cry1F corn in fall armyworm (Spodoptera frugiperda) from Florida. Sci. Rep. 2022, 12, 13580. [Google Scholar] [CrossRef]
- Heckel, D.G.; Gahan, L.J.; Baxter, S.W.; Zhao, J.Z.; Shelton, A.M.; Gould, F.; Tabashnik, B.E. The diversity of Bt resistance genes in species of Lepidoptera. J. Invertebr. Pathol. 2007, 95, 192–197. [Google Scholar] [CrossRef]
- Wu, Y. Detection and mechanisms of resistance evolved in insects to Cry toxins from Bacillus thuringiensis. In Advances in Insect Physiology; Dhadialla, T.S., Gill, S.S., Eds.; Academic Press: Oxford, UK; San Diego, CA, USA, 2014; Volume 47, pp. 297–342. [Google Scholar] [CrossRef]
- De Bortoli, C.P.; Jurat-Fuentes, J.L. Mechanisms of resistance to commercially relevant entomopathogenic bacteria. Curr. Opin. Insect Sci. 2019, 33, 56–62. [Google Scholar] [CrossRef]
- Heckel, D.G. The essential and enigmatic role of ABC transporters in Bt resistance of noctuids and other insect pests of agriculture. Insects 2021, 12, 389. [Google Scholar] [CrossRef]
- Qi, L.; Hanyang, D.; Jin, Z.; Shen, H.; Guan, F.; Yang, Y.; Tabashnik, B.E.; Wu, Y. Evaluating cross-resistance to Cry and Vip toxins in four strains of Helicoverpa armigera with different genetic mechanisms of resistance to Bt toxin Cry1Ac. Front. Microbiol. 2021, 12, 670402. [Google Scholar] [CrossRef]
- Endo, H. Molecular and kinetic models for pore formation of Bacillus thuringiensis Cry toxin. Toxins 2022, 14, 433. [Google Scholar] [CrossRef]
- Kain, W.; Cotto-Rivera, R.O.; Wang, P. Resistance of cabbage loopers to Bacillus thuringiensis (Bt) toxin Cry1F and to dual-Bt toxin WideStrike cotton plants. Appl. Environ. Microbiol. 2022, 88, e0119422. [Google Scholar] [CrossRef] [PubMed]
- Liao, C.Y.; Zhang, D.D.; Cheng, Y.; Yang, Y.B.; Liu, K.Y.; Wu, K.M.; Xiao, Y.T. Down-regulation of HaABCC3, potentially mediated by a cis-regulatory mechanism, is involved in resistance to Cry1Ac in the cotton bollworm, Helicoverpa armigera. Insect Sci. 2022, 30, 135–145. [Google Scholar] [CrossRef] [PubMed]
- Mathew, L.G.; Ponnuraj, J.; Mallappa, B.; Chowdary, L.R.; Zhang, J.; Tay, W.T.; Walsh, T.K.; Gordon, K.H.J.; Heckel, D.G.; Downes, S.; et al. ABC transporter mis-splicing associated with resistance to Bt toxin Cry2Ab in laboratory- and field-selected pink bollworm. Sci. Rep. 2018, 8, 13531. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wang, J.; Zhang, H.; Wang, H.; Zhao, S.; Zuo, Y.; Yang, Y.; Wu, Y. Functional validation of cadherin as a receptor of Bt toxin Cry1Ac in Helicoverpa armigera utilizing the CRISPR/Cas9 system. Insect Biochem. Mol. Biol. 2016, 76, 11–17. [Google Scholar] [CrossRef]
- Wang, J.; Wang, H.; Liu, S.; Liu, L.; Tay, W.T.; Walsh, T.K.; Yang, Y.; Wu, Y. CRISPR/Cas9 mediated genome editing of Helicoverpa armigera with mutations of an ABC transporter gene HaABCA2 confers resistance to Bacillus thuringiensis Cry2A toxins. Insect Biochem. Mol. Biol. 2017, 87, 147–153. [Google Scholar] [CrossRef]
- Yang, X.; Chen, W.; Song, X.; Ma, X.; Cotto-Rivera, R.O.; Kain, W.; Chu, H.; Chen, Y.R.; Fei, Z.; Wang, P. Mutation of ABC transporter ABCA2 confers resistance to Bt toxin Cry2Ab in Trichoplusia ni. Insect Biochem. Mol. Biol. 2019, 112, 103209. [Google Scholar] [CrossRef]
- Guo, Z.; Sun, D.; Kang, S.; Zhou, J.; Gong, L.; Qin, J.; Guo, L.; Zhu, L.; Bai, Y.; Luo, L.; et al. CRISPR/Cas9-mediated knockout of both the PxABCC2 and PxABCC3 genes confers high-level resistance to Bacillus thuringiensis Cry1Ac toxin in the diamondback moth, Plutella xylostella (L.). Insect Biochem. Mol. Biol. 2019, 107, 31–38. [Google Scholar] [CrossRef]
- Huang, J.L.; Xu, Y.J.; Zuo, Y.Y.; Yang, Y.H.; Tabashnik, B.E.; Wu, Y.D. Evaluation of five candidate receptors for three Bt toxins in the beet armyworm using CRISPR-mediated gene knockouts. Insect Biochem. Mol. Biol. 2020, 121, 103361. [Google Scholar] [CrossRef]
- Wang, X.; Xu, Y.; Huang, J.; Jin, W.; Yang, Y.; Wu, Y. CRISPR-mediated knockout of the ABCC2 gene in Ostrinia furnacalis confers high-level resistance to the Bacillus thuringiensis Cry1Fa toxin. Toxins 2020, 12, 246. [Google Scholar] [CrossRef] [Green Version]
- Perera, O.P.; Little, N.S.; Abdelgaffar, H.; Jurat-Fuentes, J.L.; Reddy, G.V.P. Genetic knockouts indicate that the ABCC2 protein in the bollworm Helicoverpa zea is not a major receptor for the Cry1Ac insecticidal protein. Genes 2021, 12, 1522. [Google Scholar] [CrossRef] [PubMed]
- Fabrick, J.A.; Heu, C.C.; LeRoy, D.M.; DeGain, B.A.; Yelich, A.J.; Unnithan, G.C.; Wu, Y.; Li, X.; Carrière, Y.; Tabashnik, B.E. Knockout of ABC transporter gene ABCA2 confers resistance to Bt toxin Cry2Ab in Helicoverpa zea. Sci. Rep. 2022, 12, 16706. [Google Scholar] [CrossRef] [PubMed]
- Sun, D.; Zhu, L.; Guo, L.; Wang, S.; Wu, Q.; Crickmore, N.; Zhou, X.; Bravo, A.; Soberón, M.; Guo, Z.; et al. A versatile contribution of both aminopeptidases N and ABC transporters to Bt Cry1Ac toxicity in the diamondback moth. BMC Biol. 2022, 20, 33. [Google Scholar] [CrossRef] [PubMed]
- Gao, Q.; Lin, Y.; Wang, X.; Jing, D.; Wang, Z.; He, K.; Bai, S.; Zhang, Y.; Zhang, T. Knockout of ABC transporter ABCG4 gene confers resistance to Cry1 proteins in Ostrinia furnacalis. Toxins 2022, 14, 52. [Google Scholar] [CrossRef]
- Henneberry, T.J.; Naranjo, S.E. Integrated management approaches for pink bollworm in the southwestern United States. Integr. Pest Manag. Rev. 1998, 3, 31–52. [Google Scholar] [CrossRef]
- CABI. Pectinophora gossypiella (Pink Bollworm). 2022. Available online: https://www.cabi.org/isc/datasheet/39417 (accessed on 16 December 2022).
- BCH-LMO-SCBD-14775-1-6. Bollgard Cotton, Transformation Event 531 (MON-00531-6). Available online: https://bch.cbd.int/en/database/LMO/BCH-LMO-SCBD-14775 (accessed on 17 October 2022).
- Tabashnik, B.E.; Liu, Y.B.; De Maagd, R.A.; Dennehy, T.J. Cross-resistance of pink bollworm (Pectinophora gossypiella) to Bacillus thuringiensis toxins. Appl. Environ. Microbiol. 2000, 66, 4582–4584. [Google Scholar] [CrossRef] [Green Version]
- Tabashnik, B.E.; Unnithan, G.C.; Yelich, A.J.; Fabrick, J.A.; Dennehy, T.J.; Carrière, Y. Responses to Bt toxin Vip3Aa by pink bollworm larvae resistant or susceptible to Cry toxins. Pest Manag. Sci. 2022, 78, 3973–3979. [Google Scholar] [CrossRef]
- Tabashnik, B.E.; Carrière, Y. Global patterns of resistance to Bt crops highlighting pink bollworm in the United States, China, and India. J. Econ. Entomol. 2019, 112, 2513–2523. [Google Scholar] [CrossRef]
- Wan, P.; Xu, D.; Cong, S.; Jiang, Y.; Huang, Y.; Wang, J.; Wu, H.; Wang, L.; Wu, K.; Carrière, Y.; et al. Hybridizing transgenic Bt cotton with non-Bt cotton counters resistance in pink bollworm. Proc. Natl. Acad. Sci. USA 2017, 114, 5413–5418. [Google Scholar] [CrossRef] [Green Version]
- Naik, V.C.; Kumbhare, S.; Kranthi, S.; Satija, U.; Kranthi, K.R. Field-evolved resistance of pink bollworm, Pectinophora gossypiella (Saunders) (Lepidoptera: Gelechiidae), to transgenic Bacillus thuringiensis (Bt) cotton expressing crystal 1Ac (Cry1Ac) and Cry2Ab in India. Pest Manag. Sci. 2018, 74, 2544–2554. [Google Scholar] [CrossRef]
- Carrière, Y.; Ellers-Kirk, C.; Kumar, K.; Heuberger, S.; Whitlow, M.; Antilla, L.; Dennehy, T.J.; Tabashnik, B.E. Long-term evaluation of compliance with refuge requirements for Bt cotton. Pest Manag. Sci. 2005, 61, 327–330. [Google Scholar] [CrossRef] [PubMed]
- Tabashnik, B.E.; Dennehy, T.J.; Carrière, Y. Delayed resistance to transgenic cotton in pink bollworm. Proc. Natl. Acad. Sci. USA 2005, 102, 15389–15393. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Tabashnik, B.E.; Sisterson, M.S.; Ellsworth, P.C.; Dennehy, T.J.; Antilla, L.; Liesner, L.; Whitlow, M.; Staten, R.T.; Fabrick, J.A.; Unnithan, G.C.; et al. Suppressing resistance to Bt cotton with sterile insect releases. Nat. Biotechnol. 2010, 28, 1304–1307. [Google Scholar] [CrossRef] [PubMed]
- Tabashnik, B.; Morin, S.; Unnithan, G.; Yelich, A.; Ellers-Kirk, C.; Harpold, V.; Sisterson, M.; Ellsworth, P.; Dennehy, T.; Antilla, L.; et al. Sustained susceptibility of pink bollworm to Bt cotton in the United States. GM Crops Food 2012, 3, 194–200. [Google Scholar] [CrossRef] [Green Version]
- Carrière, Y.; Ellers-Kirk, C.; Patin, A.L.; Sims, M.; Meyer, S.; Liu, Y.B.; Dennehy, T.J.; Tabashnik, B.E. Overwintering cost associated with resistance to transgenic cotton in the pink bollworm. J. Econ. Entomol. 2001, 94, 935–941. [Google Scholar] [CrossRef] [PubMed]
- Carrière, Y.; Ellers-Kirk, C.; Biggs, R.W.; Nyboer, M.E.; Unnithan, G.C.; Dennehy, T.J.; Tabashnik, B.E. Cadherin-based resistance to Bacillus thuringiensis cotton in hybrid strains of pink bollworm: Fitness costs and incomplete resistance. J. Econ. Entomol. 2006, 99, 1925–1935. [Google Scholar] [CrossRef]
- Carrière, Y.; Williams, J.L.; Crowder, D.W.; Tabashnik, B.E. Genotype-specific fitness cost of resistance to Bt toxin Cry1Ac in pink bollworm. Pest Manag. Sci. 2018, 74, 2496–2503. [Google Scholar] [CrossRef]
- Carrière, Y.; Tabashnik, B.E. Reversing insect adaptation to transgenic insecticidal plants. Proc. R. Soc. B 2001, 268, 1475–1480. [Google Scholar] [CrossRef] [Green Version]
- Gassmann, A.J.; Carrière, Y.; Tabashnik, B.E. Fitness costs of insect resistance to Bacillus thuringiensis. Ann. Rev. Entomol. 2009, 54, 147–163. [Google Scholar] [CrossRef]
- Carrière, Y.; Crowder, D.W.; Tabashnik, B.E. Evolutionary ecology of insect adaptation to Bt crops. Evol. Appl. 2010, 3, 561–573. [Google Scholar] [CrossRef]
- Grefenstette, B.; El-Lissy, O.; Staten, R.T.; Pink Bollworm Eradication Plan in the US. US Department of Agriculture, Animal and Plant Health Protection Service. 2009. Available online: https://www.aphis.usda.gov/plant_health/plant_pest_info/cotton_pests/downloads/pbw-erad-plan2-09.pdf (accessed on 17 October 2022).
- Staten, R.T.; Walters, M.L. Technology used by field managers for pink bollworm eradication with its successful outcome in the United States and Mexico. In Area-Wide Integrated Pest Management: Development and Field Application; Hendrichs, J., Pereira, R., Vreysen, M.J.B., Eds.; CRC Press: Boca Raton, FL, USA, 2020; pp. 51–92. [Google Scholar]
- USDA. Eradication of Pink Bollworm. 2018. Available online: https://www.usda.gov/sites/default/files/documents/usda-pink-bollworm-proclamation.pdf (accessed on 17 October 2022).
- Liu, Y.D.; Wu, K.M.; Guo, Y.Y. Population structure and introduction history of the pink bollworm, Pectinophora gossypiella, in China. Entomol. Exp. Appl. 2009, 130, 160–172. [Google Scholar] [CrossRef]
- Wan, P.; Huang, Y.; Wu, H.; Huang, M.; Cong, S.; Tabashnik, B.E.; Wu, K. Increased frequency of pink bollworm resistance to Bt toxin Cry1Ac in China. PLoS ONE 2012, 7, e29975. [Google Scholar] [CrossRef] [PubMed]
- Wang, J.; Xu, D.; Wang, L.; Cong, S.; Wan, P.; Lei, C.; Fabrick, J.A.; Li, X.; Tabashnik, B.E.; Wu, K. Bt resistance alleles in field populations of pink bollworm from China: Similarities with the United States and decreased frequency from 2012 to 2015. Pest Manag. Sci. 2020, 76, 527–533. [Google Scholar] [CrossRef]
- Wang, L.; Xu, D.; Huang, Y.; Zhou, H.; Liu, W.; Cong, S.; Wang, J.; Li, W.; Wan, P. Mutation in the cadherin gene is a key factor for pink bollworm resistance to Bt cotton in China. Toxins 2022, 14, 23. [Google Scholar] [CrossRef] [PubMed]
- Herring, R.J. Stealth seeds: Bioproperty, biosafety, biopolitics. J. Dev. Stud. 2007, 43, 130–157. [Google Scholar] [CrossRef]
- Lalitha, N.; Ramaswami, B.; Viswanathan, P.K. India’s experience with Bt cotton: Case studies from Gujarat and Maharashtra. In Biotechnology and Agricultural Development: Transgenic Cotton, Rural Institutions and Resource-Poor Farmers; Tripp, R., Ed.; Routledge: New York, NY, USA, 2009; pp. 135–167. [Google Scholar]
- Dhurua, S.; Gujar, G.T. Field-evolved resistance to Bt toxin Cry1Ac in the pink bollworm, Pectinophora gossypiella (Saunders) (Lepidoptera: Gelechiidae), from India. Pest Manag. Sci. 2011, 67, 898–903. [Google Scholar] [CrossRef] [PubMed]
- Mohan, K.S.; Ravi, K.C.; Suresh, P.J.; Sumerford, D.; Head, G.P. Field resistance to the Bacillus thuringiensis protein Cry1Ac expressed in Bollgard® hybrid cotton in pink bollworm, Pectinophora gossypiella (Saunders), populations in India. Pest Manag. Sci. 2016, 72, 738–746. [Google Scholar] [CrossRef]
- Ojha, A.; Sowjanya Sree, K.; Sachdev, B.; Rashmi, M.A.; Ravi, K.C.; Suresh, P.J.; Mohan, K.S.; Bhatnagar, R.K. Analysis of resistance to Cry1Ac in field-collected pink bollworm, Pectinophora gossypiella (Lepidoptera: Gelechiidae), populations. GM Crops Food 2014, 5, 280–286. [Google Scholar] [CrossRef] [Green Version]
- ISAAA. Global Status of Commercialized Biotech/GM Crops in 2017: Biotech Crop Adoption Surges as Economic Benefits Accumulate in 22 Years; ISAAA Brief No. 53; ISAAA: Ithaca, NY, USA, 2017. [Google Scholar]
- Najork, K.; Gadela, S.; Nadiminti, P.; Gosikonda, S.; Reddy, R.; Haribabu, E.; Keck, M. The return of pink bollworm in India’s Bt cotton fields: Livelihood vulnerabilities of farming households in Karimnagar District. Prog. Dev. Stud. 2021, 21, 68–85. [Google Scholar] [CrossRef]
- Najork, K.; Keck, M. Mistranslating refuge crops: Analyzing policy mobilities in the context of Indian Bt cotton production. Geogr. Helv. 2022, 77, 213–230. [Google Scholar] [CrossRef]
- Najork, K.; Friedrich, J.; Keck, M. Bt cotton, pink bollworm, and the political economy of sociobiological obsolescence: Insights from Telangana, India. Agric. Hum. Values 2022, 39, 1007–1026. [Google Scholar] [CrossRef]
- Fand, B.B.; Nagrare, V.S.; Gawande, S.P. Widespread infestation of pink bollworm, Pectinophora gossypiella (Saunders) (Lepidoptera: Gelechidae) on Bt cotton in Central India: A new threat and concerns for cotton production. Phytoparasitica 2019, 47, 313–325. [Google Scholar] [CrossRef]
- Naik, V.C.B.; Subbireddy, K.B.; Kranthi, S.; Nagrare, V.S.; Kumbhare, S.; Gokte-Narkhedkar, N.; Waghmare, V.N. Pink bollworm, Pectinophora gossypiella (Saunders) (Lepidoptera: Gelechiidae) survival on transgenic cotton in India. Egypt J. Biol. Pest Control 2021, 31, 40. [Google Scholar] [CrossRef]
- Kranthi, K.R.; Stone, G.D. Long-term impacts of Bt cotton in India. Nat. Plants 2020, 6, 188–196. [Google Scholar] [CrossRef] [PubMed]
- Giridhar, B.J.; Praveen, K.V.; Singh, A. Estimating the effect of Bt cotton on input expenditure and yield: An Indian perspective. Natl. Acad. Sci. Lett. 2022, 45, 129–133. [Google Scholar] [CrossRef]
- Singh, T.V.K.; Kukanur, V.S.; Supriya, G.B. Frequency of resistance alleles to Cry1Ac toxin from cotton bollworm, Helicoverpa armigera (Hübner) collected from Bt-cotton growing areas of Telangana state of India. J. Invertebr. Pathol. 2021, 183, 107559. [Google Scholar] [CrossRef]
- Knowles, B.H.; Ellar, D.J. Colloid-osmotic lysis is a general feature of the mechanism of action of Bacillus thuringiensis [delta]-endotoxins with different insect specificity. Biochim. Biophys. Acta Gen. Subj. 1987, 924, 509–518. [Google Scholar] [CrossRef]
- Bravo, A.; Likitvivatanavong, S.; Gill, S.S.; Soberón, M. Bacillus thuringiensis: A story of a successful bioinsecticide. Insect Biochem. Mol. Biol. 2011, 41, 423–431. [Google Scholar] [CrossRef] [Green Version]
- Adang, M.J.; Crickmore, N.; Jurat-Fuentes, J.L. Diversity of Bacillus thuringiensis crystal toxins and mechanism of action. In Advances in Insect Physiology; Dhadialla, T.S., Gill, S.S., Eds.; Academic Press: Oxford, UK; San Diego, CA, USA, 2014; Volume 47, pp. 39–87. [Google Scholar] [CrossRef]
- Heckel, D.G. How do toxins from Bacillus thuringiensis kill insects? An evolutionary perspective. Arch. Insect Biochem. Physiol. 2020, 104, e21673. [Google Scholar] [CrossRef] [Green Version]
- [EPA], U.S. Environmental Protection Agency. Biopesticide Registration Action Document. Bacillus thuringiensis Cry2Ab2 Protein and Its Production in Cotton. 2003. Available online: https://www3.epa.gov/pesticides/chem_search/reg_actions/registration/decision_PC-006487_4-Mar-03.pdf (accessed on 16 December 2022).
- Tabashnik, B.E.; Zhang, M.; Fabrick, J.A.; Wu, Y.; Gao, M.; Huang, F.; Wei, J.; Zhang, J.; Yelich, A.; Unnithan, G.C.; et al. Dual mode of action of Bt proteins: Protoxin efficacy against resistant insects. Sci. Rep. 2015, 5, 15107. [Google Scholar] [CrossRef] [Green Version]
- Liu, S.; Wang, S.; Wu, S.; Wu, Y.; Yang, Y. Proteolysis activation of Cry1Ac and Cry2Ab protoxins by larval midgut juice proteases from Helicoverpa armigera. PLoS ONE 2020, 15, e0228159. [Google Scholar] [CrossRef] [PubMed]
- Fabrick, J.A.; Tabashnik, B.E. Binding of Bacillus thuringiensis toxin Cry1Ac to multiple sites of cadherin in pink bollworm. Insect Biochem. Mol. Biol. 2007, 37, 97–106. [Google Scholar] [CrossRef] [PubMed]
- Gómez, I.; Sánchez, J.; Munoz-Garay, C.; Matus, V.; Gill, S.S.; Soberón, M.; Bravo, A. Bacillus thuringiensis Cry1A toxins are versatile-proteins with multiple modes of action: Two distinct pre-pores are involved in toxicity. Biochem. J. 2014, 459, 383–396. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Li, X.; Zhao, F.; Qiu, X.; Ren, X.; Mo, X.; Ding, X.; Xia, L.; Sun, Y. The full-length Cry1Ac protoxin without proteolytic activation exhibits toxicity against insect cell line CF-203. J. Invertebr. Pathol. 2018, 152, 25–29. [Google Scholar] [CrossRef]
- Peña-Cardeña, A.; Grande, R.; Sánchez, J.; Tabashnik, B.E.; Bravo, A.; Soberón, M.; Gómez, I. The C-terminal protoxin region of Bacillus thuringiensis Cry1Ab toxin has a functional role in binding to GPI-anchored receptors in the insect midgut. J. Biol. Chem. 2018, 293, 20263–20272. [Google Scholar] [CrossRef] [Green Version]
- Qi, L.L.; Qiu, X.F.; Yang, S.S.; Li, R.; Wu, B.B.; Cao, X.M.; He, T.; Ding, X.Z.; Xia, L.Q.; Sun, Y.J. Cry1Ac protoxin and its activated toxin from Bacillus thuringiensis act differentially during the pathogenic process. J. Agric. Food Chem. 2020, 68, 5816–5824. [Google Scholar] [CrossRef]
- Liao, C.Y.; Jin, M.H.; Cheng, Y.; Yang, Y.B.; Soberón, M.; Bravo, A.; Liu, K.Y.; Xiao, Y.T. Bacillus thuringiensis Cry1Ac protoxin and activated toxin exert differential toxicity due to a synergistic interplay of cadherin with ABCC transporters in the cotton bollworm. Appl. Environ. Microbiol. 2022, 88, e02505-21. [Google Scholar] [CrossRef]
- Endo, H.; Azuma, M.; Adegawa, S.; Kikuta, S.; Sato, R. Water influx via aquaporin directly determines necrotic cell death induced by the Bacillus thuringiensis Cry toxin. FEBS Lett. 2017, 591, 56–64. [Google Scholar] [CrossRef] [Green Version]
- Zhang, X.; Candas, M.; Griko, M.; Rose-Young, L.; Bulla, L.A., Jr. Cytotoxicity of Bacillus thuringiensis Cry1Ab toxin depends on specific binding of the toxin to the cadherin receptor BT-R1 expressed in insect cells. Cell Death Differ. 2005, 12, 1407–1416. [Google Scholar] [CrossRef] [Green Version]
- Zhang, X.; Candas, M.; Griko, M.; Taussig, R.; Bulla, L.A., Jr. A mechanism of cell death involving an adenylyl cyclase/PKA signaling pathway is induced by the Cry1Ab toxin of Bacillus thuringiensis. Proc. Natl. Acad. Sci. USA 2006, 103, 9897–9902. [Google Scholar] [CrossRef] [Green Version]
- Soberón, M.; Gill, S.S.; Bravo, A. Signaling versus punching hole: How do Bacillus thuringiensis toxins kill insect midgut cells? Cell. Mol. Life Sci. 2009, 66, 1337–1349. [Google Scholar] [CrossRef] [PubMed]
- Ocelotl, J.; Sánchez, J.; Arroyo, R.; García-Gómez, B.I.; Gómez, I.; Unnithan, G.C.; Tabashnik, B.E.; Bravo, A.; Soberón, M. Binding and oligomerization of modified and native Bt toxins in resistant and susceptible pink bollworm. PLoS ONE 2015, 10, e0144086. [Google Scholar] [CrossRef] [PubMed]
- Tabashnik, B.E.; Liu, Y.B.; Malvar, T.; Masson, L.; Ferré, J. Insect resistance to Bacillus thuringiensis: Uniform or diverse? Phil. Trans. Roy. Soc. Lond. B 1998, 353, 1751–1756. [Google Scholar] [CrossRef]
- Morin, S.; Biggs, R.W.; Sisterson, M.S.; Shriver, L.; Ellers-Kirk, C.; Higginson, D.; Holley, D.; Gahan, L.J.; Heckel, D.G.; Carrière, Y.; et al. Three cadherin alleles associated with resistance to Bacillus thuringiensis in pink bollworm. Proc. Natl. Acad. Sci. USA 2003, 100, 5004–5009. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Fabrick, J.A.; Forlow Jech, L.; Henneberry, T.J. Novel pink bollworm resistance to the Bt toxin Cry1Ac: Effects on mating, oviposition, larval development and survival. J. Insect Sci. 2009, 9, 24. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Fabrick, J.A.; Mathew, L.G.; LeRoy, D.M.; Hull, J.J.; Unnithan, G.C.; Yelich, A.J.; Carrière, Y.; Li, X.; Tabashnik, B.E. Reduced cadherin expression associated with resistance to Bt toxin Cry1Ac in pink bollworm. Pest Manag. Sci. 2020, 76, 67–74. [Google Scholar] [CrossRef]
- Wang, L.; Ma, Y.; Wan, P.; Liu, K.; Xiao, Y.; Wang, J.; Cong, S.; Xu, D.; Wu, K.; Fabrick, J.A.; et al. Resistance to Bacillus thuringiensis linked with a cadherin transmembrane mutation affecting cellular trafficking in pink bollworm from China. Insect Biochem. Mol. Biol. 2018, 94, 28–35. [Google Scholar] [CrossRef]
- Wang, L.; Ma, Y.; Guo, X.; Wan, P.; Liu, K.; Cong, S.; Wang, J.; Xu, D.; Xiao, Y.; Li, X.; et al. Pink bollworm resistance to Bt toxin Cry1Ac associated with an insertion in cadherin exon 20. Toxins 2019, 11, 186. [Google Scholar] [CrossRef] [Green Version]
- Wang, L.; Wang, J.; Ma, Y.; Wan, P.; Liu, K.; Cong, S.; Xiao, Y.; Xu, D.; Wu, K.; Fabrick, J.A.; et al. Transposon insertion causes cadherin mis-splicing and confers resistance to Bt cotton in pink bollworm from China. Sci. Rep. 2019, 9, 7479. [Google Scholar] [CrossRef] [Green Version]
- Nair, R.; Kamath, S.P.; Mohan, K.S.; Head, G.; Sumerford, D.V. Inheritance of field-relevant resistance to the Bacillus thuringiensis protein Cry1Ac in Pectinophora gossypiella (Lepidoptera: Gelechiidae) collected from India. Pest Manag. Sci. 2016, 72, 558–565. [Google Scholar] [CrossRef]
- Mittal, A.; Ningthoukhongjam, J.; Malthankar, P.; Ningthoujam, K.; Kalia, V.; Gujar, G. Toxicity of Cry1Ac inheritance and fitness cost association in pink bollworm, Pectinophora gossypiella (Lepidoptera: Gelichiidae). Biopestic. Int. 2016, 12, 127–138. [Google Scholar]
- Tabashnik, B.E.; Biggs, R.W.; Higginson, D.; Henderson, S.; Unnithan, D.C.; Unnithan, G.C.; Ellers-Kirk, C.; Sisterson, M.S.; Dennehy, T.J.; Carrière, Y.; et al. Association between resistance to Bt cotton and cadherin genotype in pink bollworm. J. Econ. Entomol. 2005, 98, 635–644. [Google Scholar] [CrossRef] [PubMed]
- Liu, Y.-B.; Tabashnik, B.E.; Meyer, S.K.; Carrière, Y.; Bartlett, A.C. Genetics of pink bollworm resistance to Bacillus thuringiensis toxin Cry1Ac. J. Econ. Entomol. 2001, 94, 248–252. [Google Scholar] [CrossRef] [PubMed]
- Liu, Y.B.; Tabashnik, B.E.; Dennehy, T.J.; Patin, A.L.; Sims, M.A.; Meyer, S.K.; Carrière, Y. Effects of Bt cotton and Cry1Ac toxin on survival and development of pink bollworm (Lepidoptera: Gelechiidae). J. Econ. Entomol. 2001, 94, 1237–1242. [Google Scholar] [CrossRef]
- Tabashnik, B.E.; Liu, Y.B.; Dennehy, T.J.; Sims, M.A.; Sisterson, M.S.; Biggs, R.W.; Carrière, Y. Inheritance of resistance to Bt toxin Cry1Ac in a field-derived strain of pink bollworm (Lepidoptera: Gelechiidae). J. Econ. Entomol. 2002, 95, 1018–1026. [Google Scholar] [CrossRef] [PubMed]
- Tabashnik, B.E.; Liu, Y.B.; Unnithan, G.C.; Carrière, Y.; Dennehy, T.J.; Morin, S. Shared genetic basis of resistance to Bt toxin Cry1Ac in independent strains of pink bollworm. J. Econ. Entomol. 2004, 97, 721–726. [Google Scholar] [CrossRef] [PubMed]
- Fabrick, J.A.; Tabashnik, B.E. Similar genetic basis of resistance to Bt toxin Cry1Ac in boll-selected and diet-selected strains of pink bollworm. PLoS ONE 2012, 7, e35658. [Google Scholar] [CrossRef] [Green Version]
- Liu, Y.B.; Tabashnik, B.E. Inheritance of resistance to Bacillus thuringiensis toxin Cry1C in the diamondback moth. Appl. Environ. Microbiol. 1997, 63, 2218–2223. [Google Scholar] [CrossRef] [Green Version]
- Liu, Y.B.; Tabashnik, B.E.; Dennehy, T.J.; Patin, A.L.; Bartlett, A.C. Development time and resistance to Bt crops. Nature 1999, 400, 519. [Google Scholar] [CrossRef]
- Henneberry, T.J.; Forlow Jech, L.; de la Torre, T. Effects of transgenic cotton on mortality and development of pink bollworm (Lepidoptera: Gelechiidae) larvae. Southwest. Entomol. 2001, 26, 115–128. [Google Scholar]
- Malthankar, P.A.; Gujar, G.T. Toxicity of Bacillus thuringiensis Cry2Ab and the inheritance of Cry2Ab resistance in the pink bollworm, Pectinophora gossypiella (Saunders). Indian J. Exp. Biol. 2016, 54, 586–596. [Google Scholar] [PubMed]
- Tabashnik, B.E.; Dennehy, T.J.; Sims, M.A.; Larkin, K.; Head, G.P.; Moar, W.J.; Carrière, Y. Control of resistant pink bollworm (Pectinophora gossypiella) by transgenic cotton that produces Bacillus thuringiensis toxin Cry2Ab. Appl. Environ. Microbiol. 2002, 68, 3790–3794. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Tabashnik, B.E.; Unnithan, G.C.; Masson, L.; Crowder, D.W.; Li, X.; Carrière, Y. Asymmetrical cross-resistance between Bacillus thuringiensis toxins Cry1Ac and Cry2Ab in pink bollworm. Proc. Natl. Acad. Sci. USA 2009, 106, 11889–11894. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Fabrick, J.A.; Unnithan, G.C.; Yelich, A.J.; DeGain, B.; Masson, L.; Zhang, J.; Carrière, Y.; Tabashnik, B.E. Multi-toxin resistance enables pink bollworm survival on pyramided Bt cotton. Sci. Rep. 2015, 5, 16554. [Google Scholar] [CrossRef] [Green Version]
- Karim, S.; Riazuddin, S.; Gould, F.; Dean, D.H. Determination of receptor binding properties of Bacillus thuringiensis delta-endotoxins to cotton bollworm (Helicoverpa zea) and pink bollworm (Pectinophora gossypiella) midgut brush border membrane vesicles. Pestic. Biochem. Physiol. 2000, 67, 198–216. [Google Scholar] [CrossRef]
- Gonzalez-Cabrera, J.; Escriche, B.; Tabashnik, B.E.; Ferre, J. Binding of Bacillus thuringiensis toxins in resistant and susceptible strains of pink bollworm (Pectinophora gossypiella). Insect Biochem. Mol. Biol. 2003, 33, 929–935. [Google Scholar] [CrossRef]
- Tabashnik, B.E.; Biggs, R.W.; Fabrick, J.A.; Gassmann, A.J.; Dennehy, T.J.; Carrière, Y.; Morin, S. High-level resistance to Bacillus thuringiensis toxin Cry1Ac and cadherin genotype in pink bollworm. J. Econ. Entomol. 2006, 99, 2125–2131. [Google Scholar] [CrossRef]
- Fabrick, J.A.; Mathew, L.G.; Tabashnik, B.E.; Li, X. Insertion of an intact CR1 retrotransposon in a cadherin gene linked with Bt resistance in the pink bollworm, Pectinophora gossypiella. Insect Mol. Biol. 2011, 20, 651–665. [Google Scholar] [CrossRef]
- Fabrick, J.A.; Ponnuraj, J.; Singh, A.; Tanwar, R.K.; Unnithan, G.C.; Yelich, A.J.; Li, X.; Carrière, Y.; Tabashnik, B.E. Alternative splicing and highly variable cadherin transcripts associated with field-evolved resistance of pink bollworm to Bt cotton in India. PLoS ONE 2014, 9, e97900. [Google Scholar] [CrossRef]
- Tabashnik, B.E.; Fabrick, J.A.; Henderson, S.; Biggs, R.W.; Yafuso, C.M.; Nyboer, M.E.; Manhardt, N.M.; Coughlin, L.A.; Sollome, J.; Carrière, Y.; et al. DNA screening reveals pink bollworm resistance to Bt cotton remains rare after a decade of exposure. J. Econ. Entomol. 2006, 99, 1525–1530. [Google Scholar] [CrossRef]
- Li, X.; Schuler, M.A.; Berenbaum, M.R. Molecular mechanisms of metabolic resistance to synthetic and natural xenobiotics. Ann. Rev. Entomol. 2007, 52, 231–253. [Google Scholar] [CrossRef] [PubMed]
- Rostant, W.G.; Wedell, N.; Hosken, D.J. Chapter 2—Transposable Elements and Insecticide Resistance. In Advances in Genetics; Goodwin, S.F., Friedmann, T., Dunlap, J.C., Eds.; Academic Press: Cambridge, MA, USA, 2012; Volume 78, pp. 169–201. [Google Scholar] [CrossRef]
- Li, S.; Hussain, F.; Unnithan, G.C.; Dong, S.; UlAbdin, Z.; Gu, S.; Mathew, L.G.; Fabrick, J.A.; Ni, X.; Carrière, Y.; et al. A long non-coding RNA regulates cadherin transcription and susceptibility to Bt toxin Cry1Ac in pink bollworm, Pectinophora gossypiella. Pestic. Biochem. Physiol. 2019, 158, 54–60. [Google Scholar] [CrossRef] [PubMed]
- Tabashnik, B.E.; Patin, A.L.; Dennehy, T.J.; Liu, Y.B.; Carrière, Y.; Sims, M.A.; Antilla, L. Frequency of resistance to Bacillus thuringiensis in field populations of pink bollworm. Proc. Natl. Acad. Sci. USA 2000, 97, 12980–12984. [Google Scholar] [CrossRef] [Green Version]
- Causier, B.; Li, Z.; De Smet, R.; Lloyd, J.P.B.; Van de Peer, Y.; Davies, B. Conservation of nonsense-mediated mRNA decay complex components throughout eukaryotic evolution. Sci. Rep. 2017, 7, 16692. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Nickless, A.; Bailis, J.M.; You, Z. Control of gene expression through the nonsense-mediated RNA decay pathway. Cell Biosci. 2017, 7, 26. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Fabrick, J.A.; LeRoy, D.M.; Unnithan, G.C.; Yelich, A.J.; Carrière, Y.; Li, X.; Tabashnik, B.E. Shared and independent genetic basis of resistance to Bt toxin Cry2Ab in two strains of pink bollworm. Sci. Rep. 2020, 10, 7988. [Google Scholar] [CrossRef] [PubMed]
- Fabrick, J.A.; LeRoy, D.M.; Mathew, L.G.; Wu, Y.; Unnithan, G.C.; Yelich, A.J.; Carrière, Y.; Li, X.; Tabashnik, B.E. CRISPR-mediated mutations in the ABC transporter gene ABCA2 confer pink bollworm resistance to Bt toxin Cry2Ab. Sci. Rep. 2021, 11, 10377. [Google Scholar] [CrossRef]
- Malthankar, P.A.; Gujar, G.T. Association of Cry2Ab resistance with alkaline phosphatase in pink bollworm Pectinophora gossypiella (Saunders) (Lepidoptera: Gelechiidae). Biopestic. Int. 2015, 10, 143–151. [Google Scholar]
- Schlum, K.; Lamour, K.; Tandy, P.; Emrich, S.J.; de Bortoli, C.P.; Rao, T.; Viteri Dillon, D.M.; Linares-Ramirez, A.M.; Jurat-Fuentes, J.L. Genetic screening to identify candidate resistance alleles to Cry1F corn in fall armyworm using targeted sequencing. Insects 2021, 12, 618. [Google Scholar] [CrossRef]
- Gould, F.; Anderson, A.; Jones, A.; Sumerford, D.; Heckel, D.G.; Lopez, J.; Micinski, S.; Leonard, R.; Laster, M. Initial frequency of alleles for resistance to Bacillus thuringiensis toxins in field populations of Heliothis virescens. Proc. Natl. Acad. Sci. USA 1997, 94, 3519–3523. [Google Scholar] [CrossRef] [Green Version]
- Zhang, H.; Tian, W.; Zhao, J.; Jin, L.; Yang, J.; Liu, C.; Yang, Y.; Wu, S.; Wu, K.; Cui, J.; et al. Diverse genetic basis of field-evolved resistance to Bt cotton in cotton bollworm from China. Proc. Natl. Acad. Sci. USA 2012, 109, 10275–10280. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Morin, S.; Henderson, S.; Fabrick, J.A.; Carrière, Y.; Dennehy, T.J.; Tabashnik, B.E.; Brown, J.K. DNA-based detection of Bt resistance alleles in pink bollworm. Insect Biochem. Mol. Biol. 2004, 34, 1225–1233. [Google Scholar] [CrossRef] [PubMed]
- Caccia, S.; Hernandez-Rodriguez, C.S.; Mahon, R.J.; Downes, S.; James, W.; Bautsoens, N.; Van Rie, J.; Ferre, J. Binding site alteration is responsible for field-isolated resistance to Bacillus thuringiensis Cry2A insecticidal proteins in two Helicoverpa species. PLoS ONE 2010, 5, e9975. [Google Scholar] [CrossRef] [PubMed] [Green Version]






{kind=link}
{kind=link}
{kind=link}
{kind=link}
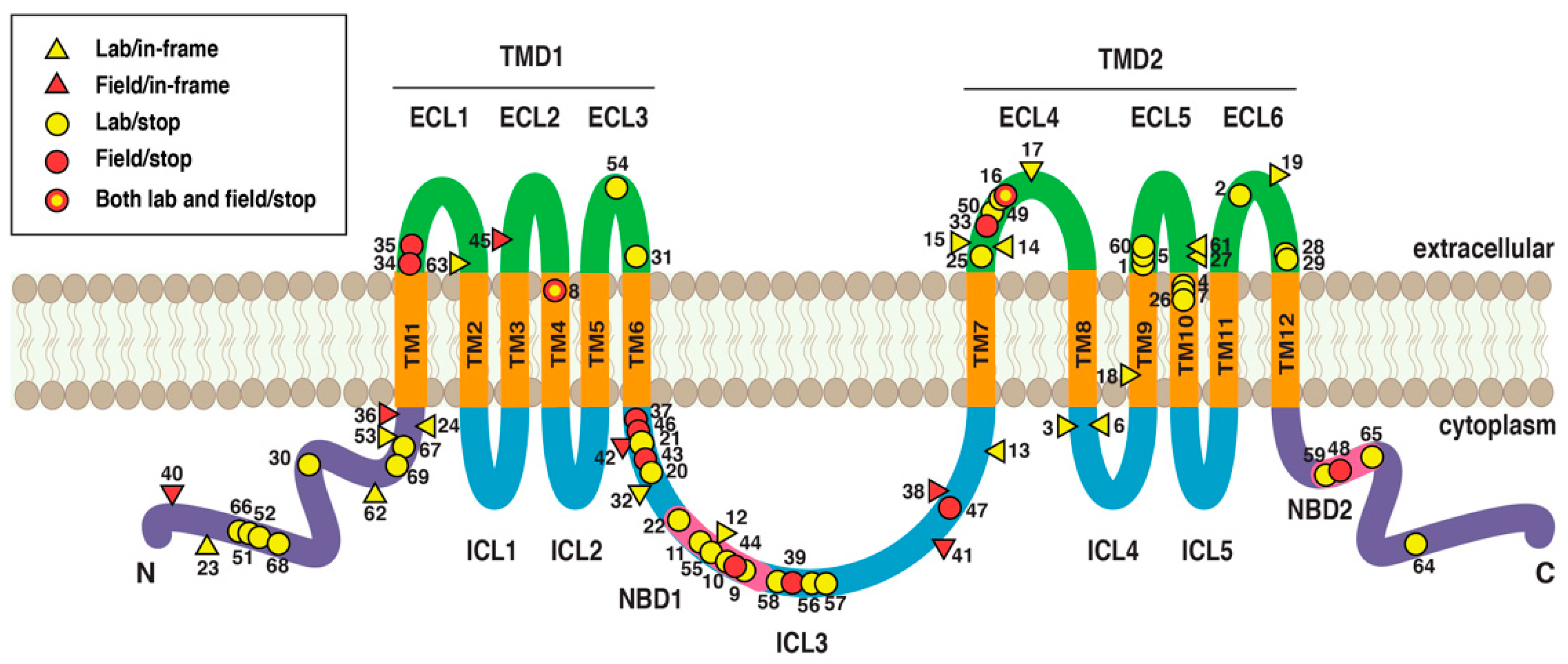{kind=link}
| Resistance Trait | Country | ||
|---|---|---|---|
| China a | India b | U.S. a | |
| Cry1Ac | |||
| Resistance ratio > 100 c | Yes | Yes | Yes |
| Recessive inheritance | Yes | Yes | Yes |
| Weak cross-resistance d | Yes | Yes | Yes |
| Reduced binding e | Yes | Yes | Yes f |
| PgCad1 resistance alleles | r1, r2, r13–r16, r18–r20 | r5–r12 | r1–r4, r17 |
| Cry2Ab | |||
| Resistance ratio > 100 c | NA g | Varies h | Yes |
| Recessive inheritance | NA | NA | Varies |
| Weak cross-resistance i | NA | NA | Varies |
| Reduced binding e | NA | NA | NA |
| PgABCA2 mutations j | NA | Yes | Varies |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Fabrick, J.A.; Li, X.; Carrière, Y.; Tabashnik, B.E. Molecular Genetic Basis of Lab- and Field-Selected Bt Resistance in Pink Bollworm. Insects 2023, 14, 201. https://doi.org/10.3390/insects14020201
Fabrick JA, Li X, Carrière Y, Tabashnik BE. Molecular Genetic Basis of Lab- and Field-Selected Bt Resistance in Pink Bollworm. Insects. 2023; 14(2):201. https://doi.org/10.3390/insects14020201
Chicago/Turabian StyleFabrick, Jeffrey A., Xianchun Li, Yves Carrière, and Bruce E. Tabashnik. 2023. "Molecular Genetic Basis of Lab- and Field-Selected Bt Resistance in Pink Bollworm" Insects 14, no. 2: 201. https://doi.org/10.3390/insects14020201