
Effect of Wolbachia Infection and Adult Food on the Sexual Signaling of Males of the Mediterranean Fruit Fly Ceratitis capitata



Abstract
:Simple Summary
Abstract
1. Introduction
2. Materials and Methods
2.1. Experimental Conditions and Ceratitis Capitata Lines
2.2. Effect of Age and Food Type on Signaling Performance
2.3. Statistical Analyses
3. Results
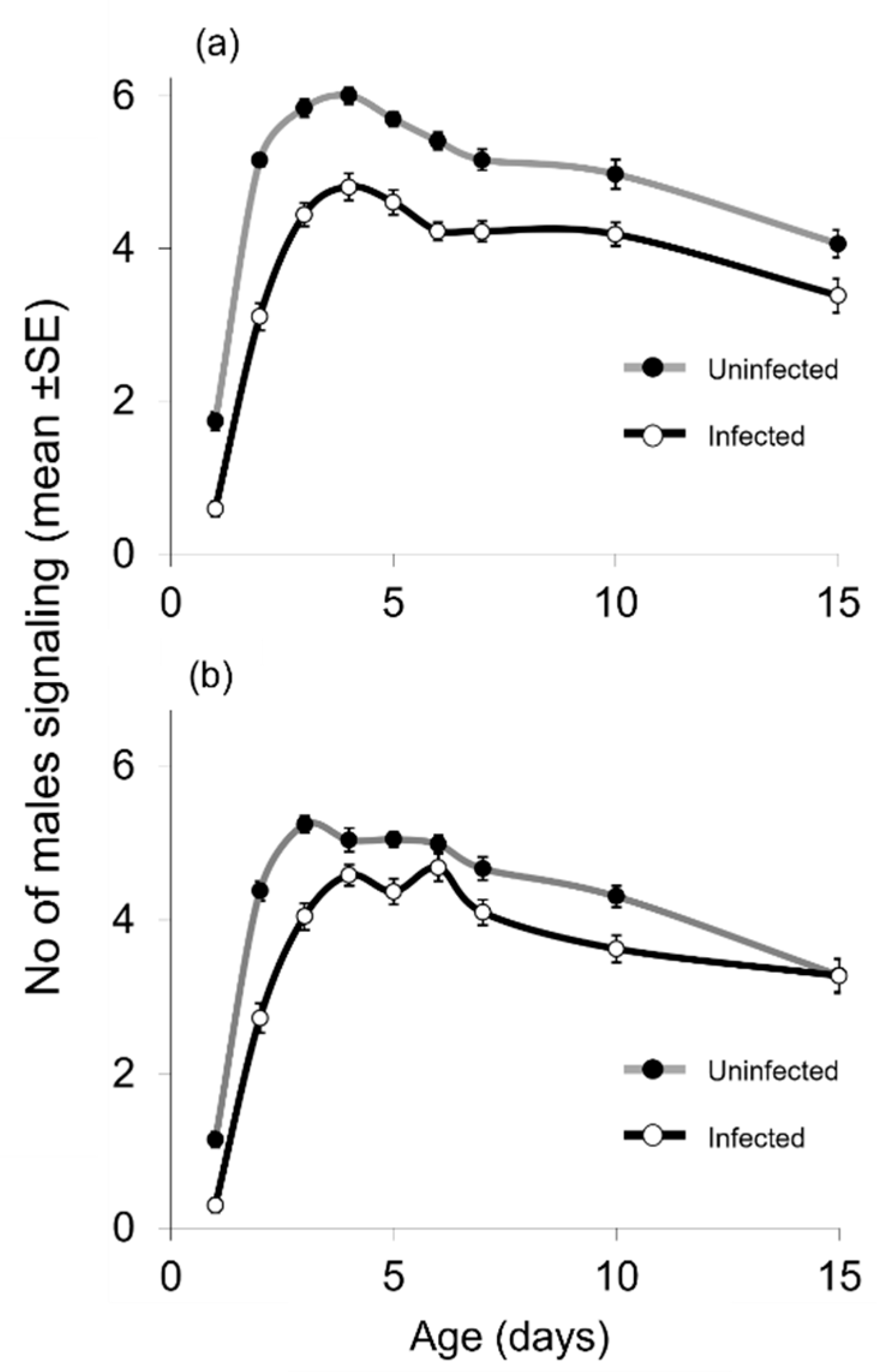
3.1. Effect of Infection, Age, and Adult Food
3.2. Effect of Infection, Food, and Time of Day
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Liquido, N.J.; Cunningham, R.T.; Nakagawa, S. Host Plants of Mediterranean Fruit Fly (Diptera: Tephritidae) on the Island of Hawaii (1949–1985 Survey). J. Econ. Entomol. 1990, 83, 1863–1878. [Google Scholar] [CrossRef]
- White, I.M.; Elson-Harris, M.M. Fruit Flies of Economic Significance: Their Identification and Bionomics; CAB International: Wallingford, UK, 1992; ISBN 0-85198-790-7. [Google Scholar]
- Aluja, M.; Mangan, R.L. Fruit Fly (Diptera: Tephritidae) Host Status Determination: Critical Conceptual, Methodological, and Regulatory Considerations. Annu. Rev. Entomol. 2008, 53, 473–502. [Google Scholar] [CrossRef]
- Vera, M.T.; Rodriguez, R.; Segura, D.F.; Cladera, J.L.; Sutherst, R.W. Potential Geographical Distribution of the Mediterranean Fruit Fly, Ceratitis capitata (Diptera: Tephritidae), with Emphasis on Argentina and Australia. Environ. Entomol. 2002, 31, 1009–1022. [Google Scholar] [CrossRef]
- Navarro-Llopis, V.; Vacas, S.; Zarzo, M.; Primo, J. Dispersal Ability of Ceratitis capitata (Diptera: Tephritidae): Edge Effect in Area-Wide Treatments. J. Appl. Entomol. 2014, 138, 403–408. [Google Scholar] [CrossRef]
- Diamantidis, A.D.; Carey, J.R.; Nakas, C.T.; Papadopoulos, N.T. Population-Specific Demography and Invasion Potential in Medfly. Ecol. Evol. 2011, 1, 479–488. [Google Scholar] [CrossRef] [PubMed]
- Dyck, V.A.; Hendrichs, J.; Robinson, A.S. Sterile Insect Technique: Principles and Practice in Area-Wide Integrated Pest Management, 2nd ed.; CRC Press: Boca Raton, FL, USA, 2021; ISBN 978-1-00-303557-2. [Google Scholar]
- Klassen, W.; Curtis, C.F.; Hendrichs, J. History of the Sterile Insect Technique. In Sterile Insect Technique: Principles and Practice in Area-Wide Integrated Pest Management; Dyck, V.A., Hendrichs, J., Robinson, A.S., Eds.; CRC Press: Boca Raton, FL, USA, 2021; pp. 1–44. ISBN 978-1-00-303557-2. [Google Scholar]
- Knipling, E.F. Possibilities of Insect Control or Eradication through the Use of Sexually Sterile Males. J. Econ. Entomol. 1955, 48, 459–462. [Google Scholar] [CrossRef]
- Bakri, A.; Mehta, K.; Lance, D.R. Sterilizing Insects with Ionizing Radiation. In Sterile Insect Technique: Principles and Practice in Area-Wide Integrated Pest Management; Dyck, V.A., Hendrichs, J., Robinson, A.S., Eds.; CRC Press: Boca Raton, FL, USA, 2021; pp. 355–398. ISBN 978-1-00-303557-2. [Google Scholar]
- Calkins, C.O. The Importance of Understanding Fruit Fly Mating Behavior in Sterile Male Release Programs (Diptera, Tephritidae). Folia Entomol. Mex. 1984, 61, 205–213. [Google Scholar]
- Arita, L.H.; Kaneshiro, K.Y. The Dynamics of the Lek System and Mating Success in Males of the Mediterranean Fruit Fly, Ceratitis capitata (Wiedemann). Proc. Hawaiian Entomol. Soc. 1985, 25, 39–48. [Google Scholar]
- Whittier, T.S.; Kaneshiro, K.Y.; Prescott, L.D. Mating Behavior of Mediterranean Fruit Flies (Diptera: Tephritidae) in a Natural Environment. Ann. Entomol. Soc. Am. 1992, 85, 214–218. [Google Scholar] [CrossRef]
- McInnis, D.O.; Lance, D.R.; Jackson, C.G. Behavioral Resistance to the Sterile Insect Technique by Mediterranean Fruit Fly (Diptera: Tephritidae) in Hawaii. Ann. Entomol. Soc. Am. 1996, 89, 739–744. [Google Scholar] [CrossRef]
- Lance, D.R.; McInnis, D.O.; Rendon, P.; Jackson, C.G. Courtship Among Sterile and Wild Ceratitis capitata (Diptera: Tephritidae) in Field Cages in Hawaii and Guatemala. Ann. Entomol. Soc. Am. 2000, 93, 1179–1185. [Google Scholar] [CrossRef]
- Cayol, J.-P. Changes in Sexual Behavior and Life History Traits of Tephritid Species Caused by Mass-Rearing Processes. In Fruit Flies (Tephritidae) Phylogeny and Evolution of Behavior; Aluja, M., Norrbom, A., Eds.; CRC Press: Boca Raton, FL, USA, 2001; pp. 843–860. ISBN 978-0-429-12467-9. [Google Scholar]
- Lux, S.A.; Vilardi, J.C.; Liedo, P.; Gaggl, K.; Calcagno, G.E.; Munyiri, F.N.; Vera, M.T.; Manso, F. Effects of Irradiation on the Courtship Behavior of Medfly (Diptera, Tephritidae) Mass Reared for the Sterile Insect Technique. Flen 2002, 85, 102–112. [Google Scholar] [CrossRef]
- Barry, J.D.; McInnis, D.O.; Gates, D.; Morse, J.G. Effects of Irradiation on Mediterranean Fruit Flies (Diptera: Tephritidae): Emergence, Survivorship, Lure Attraction, and Mating Competition. J. Econ. Entomol. 2003, 96, 615–622. [Google Scholar] [CrossRef] [PubMed]
- Hooper, G.H.S. Sterilization of the Mediterranean Fruit Fly with Gamma Radiation: Effect on Male Competitiveness and Change in Fertility of Females Alternately Mated with Irradiated and Untreated Males. J. Econ. Entomol. 1972, 65, 1–6. [Google Scholar] [CrossRef]
- Parker, A.; Mehta, K. Sterile Insect Technique: A Model For Dose Optimization For Improved Sterile Insect Quality. Flen 2007, 90, 88–95. [Google Scholar] [CrossRef]
- Guerfali, M.M.; Parker, A.; Fadhl, S.; Hemdane, H.; Raies, A.; Chevrier, C. Fitness and Reproductive Potential of Irradiated Mass-Reared Mediterranean Fruit Fly Males Ceratitis capitata (Diptera: Tephritidae): Lowering Radiation Doses. Flen 2011, 94, 1042–1050. [Google Scholar] [CrossRef]
- Kraaijeveld, K.; Chapman, T. Effects of Male Sterility on Female Remating in the Mediterranean Fruitfly, Ceratitis capitata. Proc. R. Soc. London. Ser. B Biol. Sci. 2004, 271, S209–S211. [Google Scholar] [CrossRef]
- Leftwich, P.T.; Koukidou, M.; Rempoulakis, P.; Gong, H.-F.; Zacharopoulou, A.; Fu, G.; Chapman, T.; Economopoulos, A.; Vontas, J.; Alphey, L. Genetic Elimination of Field-Cage Populations of Mediterranean Fruit Flies. Proc. R. Soc. B Biol. Sci. 2014, 281, 20141372. [Google Scholar] [CrossRef]
- Meza, J.S.; Haq, I.-u.-; Vreysen, M.J.B.; Bourtzis, K.; Kyritsis, G.A.; Cáceres, C. Comparison of Classical and Transgenic Genetic Sexing Strains of Mediterranean Fruit Fly (Diptera: Tephritidae) for Application of the Sterile Insect Technique. PLoS ONE 2018, 13, e0208880. [Google Scholar] [CrossRef]
- Virginio, J.F.; Gómez, M.; Pinto, A.M.; Aniely, G.G.; Paranhos, B.J.; Gava, C.A.T.; Cáceres, C.; Walder, J.M.M. Male Sexual Competitiveness of Two Ceratitis capitata Strains, Tsl Vienna 8 and OX3864A Transgenics, in Field Cage Conditions. Entomol. Exp. Et Appl. 2017, 164, 318–326. [Google Scholar] [CrossRef]
- Häcker, I.; Bourtzis, K.; Schetelig, M.F. Applying Modern Molecular Technologies in Support of the Sterile Insect Technique. In Sterile Insect Technique; Dyck, V.A., Hendrichs, J., Robinson, A.S., Eds.; CRC Press: Boca Raton, FL, USA, 2021; pp. 657–702. ISBN 978-1-00-303557-2. [Google Scholar]
- Reeves, R.G.; Denton, J.A.; Santucci, F.; Bryk, J.; Reed, F.A. Scientific Standards and the Regulation of Genetically Modified Insects. PLoS Negl. Trop. Dis. 2012, 6, e1502. [Google Scholar] [CrossRef] [PubMed]
- Eberhard, W.G. Sexual Behavior and Sexual Selection in the Mediterranean Fruit Fly, Ceratitis capitata (Dacinae: Ceratitidini). In Fruit Flies (Tephritidae): Phylogeny and Evolution of Behavior; Aluja, M., Norrbom, A., Eds.; CRC Press: Boca Raton, FL, USA, 2001; pp. 459–489. ISBN 978-0-429-12467-9. [Google Scholar]
- Jang, E.B.; Light, D.M.; Flath, R.A.; Nagata, J.T.; Mon, T.R. Electroantennogram Responses of the Mediterranean Fruit Fly, Ceratitis capitata to Identified Volatile Constituents from Calling Males. Entomol. Exp. Et Appl. 1989, 50, 7–19. [Google Scholar] [CrossRef]
- Whittier, T.S.; Nam, F.Y.; Shelly, T.E.; Kaneshiro, K.Y. Male Courtship Success and Female Discrimination in the Mediterranean Fruit Fly (Diptera: Tephritidae). J. Insect Behav. 1994, 7, 159–170. [Google Scholar] [CrossRef]
- Shelly, T.E. Male Signalling and Lek Attractiveness in the Mediterranean Fruit Fly. Anim. Behav. 2000, 60, 245–251. [Google Scholar] [CrossRef]
- Shelly, T.E. Sexual Selection on Leks: A Fruit Fly Primer. J. Insect Sci. 2018, 18, 9. [Google Scholar] [CrossRef]
- Warburg, M.S.; Yuval, B. Effects of Energetic Reserves on Behavioral Patterns of Mediterranean Fruit Flies (Diptera: Tephritidae). Oecologia 1997, 112, 314–319. [Google Scholar] [CrossRef]
- Yuval, B.; Kaspi, R.; Shloush, S.; Warburg, M.S. Nutritional Reserves Regulate Male Participation in Mediterranean Fruit Fly Leks. Ecol. Entomol. 1998, 23, 211–215. [Google Scholar] [CrossRef]
- Kaspi, R.; Taylor, P.W.; Yuval, B. Diet and Size Influence Sexual Advertisement and Copulatory Success of Males in Mediterranean Fruit Fly Leks. Ecol. Entomol. 2000, 25, 279–284. [Google Scholar] [CrossRef]
- Marchini, D.; Del Bene, G.; Cappelli, L.; Dallai, R. Ultrastructure of the Male Reproductive Accessory Glands in the Medfly Ceratitis capitata (Diptera: Tephritidae) and Preliminary Characterization of Their Secretions. Arthropod Struct. Dev. 2003, 31, 313–327. [Google Scholar] [CrossRef]
- Epsky, N.D.; Heath, R.R. Food Availability and Pheromone Production by Males of Anastrepha suspensa (Diptera: Tephritidae). Environ. Entomol. 1993, 22, 942–947. [Google Scholar] [CrossRef]
- Landolt, P.J.; Sivinski, J. Effects of Time of Day, Adult Food, and Host Fruit on Incidence of Calling by Male Caribbean Fruit Flies (Diptera: Tephritidae). Environ. Entomol. 1992, 21, 382–387. [Google Scholar] [CrossRef]
- Aluja, M.; Jácome, I.; Macías-Ordóñez, R. Effect of Adult Nutrition on Male Sexual Performance in Four Neotropical Fruit Fly Species of the Genus Anastrepha (Diptera: Tephritidae). J. Insect Behav. 2001, 14, 759–775. [Google Scholar] [CrossRef]
- Liedo, P.; Orozco, D.; Cruz-López, L.; Quintero, J.L.; Becerra-Pérez, C.; del Refugio Hernández, M.; Oropeza, A.; Toledo, J. Effect of Post-Teneral Diets on the Performance of Sterile Anastrepha ludens and Anastrepha obliqua Fruit Flies. J. Appl. Entomol. 2013, 137, 49–60. [Google Scholar] [CrossRef]
- Pereira, R.; Teal, P.E.A.; Conway, H.; Worley, J.; Sivinski, J. Influence of Methoprene and Dietary Protein on Maturation and Sexual Performance of Sterile Anastrepha ludens (Diptera:Tephritidae). J. Appl. Entomol. 2013, 137, 191–199. [Google Scholar] [CrossRef]
- Shelly, T.E.; Edu, J.; Pahio, E. Influence of Diet and Dmethyl Eugenol on the Mating Success of Males of the Oriental Fruit Fly, Bactrocera dorsalis (Diptera: Tephritidae). Flen 2005, 88, 307–313. [Google Scholar] [CrossRef]
- Perez-Staples, D.; Prabhu, V.; Taylor, P.W. Post-Teneral Protein Feeding Enhances Sexual Performance of Queensland Fruit Flies. Physiol. Entomol. 2007, 32, 225–232. [Google Scholar] [CrossRef]
- Wang, F.; Chambi, C.; Li, Z.; Huang, C.; Ma, Y.; Li, C.; Tian, X.; Sangija, F.; Ntambo, M.S.; Kankonda, O.M.; et al. Influence of Supplemental Protein on the Life Expectancy and Reproduction of the Chinese Citrus Fruit Fly, Bactrocera minax (Enderlein) (Tetradacus minax) (Diptera: Tephritidae). J. Insect Sci. 2018, 18, 25. [Google Scholar] [CrossRef] [PubMed]
- Webster, R.P.; Stoffolano, J.G., Jr. The Influence of Diet on the Maturation of the Reproductive System of the Apple Maggot, Rhagoletis pomonella. Ann. Entomol. Soc. Am. 1978, 71, 844–849. [Google Scholar] [CrossRef]
- Webster, R.P.; Stoffolano, J.G., Jr.; Prokopy, R.J. Long-Term Intake of Protein and Sucrose in Relation to Reproductive Behavior of Wild and Laboratory Cultured Rhagoletis pomonella. Ann. Entomol. Soc. Am. 1979, 72, 41–46. [Google Scholar] [CrossRef]
- Teal, P.E.A.; Gomez-Simuta, Y.; Dueben, B.D.; Holler, T.C.; Olson, S. Improving the Efficacy of the Sterile Insect Technique for Fruit Flies by Incorporation of Hormone and Dietary Supplements into Adult Holding Protocols. In Proceedings of the Area-Wide Control of Insect Pests; Vreysen, M.J.B., Robinson, A.S., Hendrichs, J., Eds.; Springer Netherlands: Dordrecht, The Netherlands, 2007; pp. 163–173. [Google Scholar]
- Haq, I.-u.-; Cáceres, C.; Hendrichs, J.; Teal, P.E.A.; Stauffer, C.; Robinson, A.S. Methoprene Modulates the Effect of Diet on Male Melon Fly, Bactrocera cucurbitae, Performance at Mating Aggregations. Entomol. Exp. Et Appl. 2010, 136, 21–30. [Google Scholar] [CrossRef]
- Haq, I.-u.-; Vreysen, M.J.B.; Teal, P.E.A.; Hendrichs, J. Methoprene Application and Diet Protein Supplementation to Male Melon Fly, Bactrocera cucurbitae, Modifies Female Remating Behavior. Insect Sci. 2014, 21, 637–646. [Google Scholar] [CrossRef] [PubMed]
- Papanastasiou, S.A.; Carey, J.R.; Papadopoulos, N.T. Effects of Early-Life Protein Starvation on Longevity and Sexual Performance of Male Medfly. PLoS ONE 2019, 14, e0219518. [Google Scholar] [CrossRef] [PubMed]
- Papadopoulos, N.T.; Katsoyannos, B.I.; Kouloussis, N.A.; Economopoulos, A.P.; Carrey, J.R. Effect of Adult Age, Food, and Time of Day on Sexual Calling Incidence of Wild and Mass-Reared Ceratitis capitata Males. Entomol. Exp. Et Appl. 1998, 89, 175–182. [Google Scholar] [CrossRef]
- Diamantidis, A.D.; Papadopoulos, N.T.; Carey, J.R. Medfly Populations Differ in Diel and Age Patterns of Sexual Signalling. Entomol. Exp. Appl. 2008, 128, 389–397. [Google Scholar] [CrossRef]
- Kaspi, R.; Yuval, B. Post-Teneral Protein Feeding Improves Sexual Competitiveness But Reduces Longevity of Mass-Reared Sterile Male Mediterranean Fruit Flies (Diptera: Tephritidae). Ann. Entomol. Soc. Am. 2000, 93, 949–955. [Google Scholar] [CrossRef]
- Shelly, T.E.; McInnis, D.O. Influence of Adult Diet on the Mating Success and Survival of Male Mediterranean Fruit Flies (Diptera: Tephritidae) from Two Mass-Rearing Strains on Field-Caged Hst Trees. Flen 2003, 86, 340–344. [Google Scholar] [CrossRef]
- Shelly, T.E.; Kennelly, S.S. Starvation and the Mating Success of Wild Male Mediterranean Fruit Flies (Diptera: Tephritidae). J. Insect Behav. 2003, 16, 171–179. [Google Scholar] [CrossRef]
- Shelly, T.E.; Rendon, P.; Hernandez, E.; Salgado, S.; McInnis, D.; Villalobos, E.; Liedo, P. Effects of Diet, Ginger Root Oil, and Elevation on the Mating Competitiveness of Male Mediterranean Fruit Flies (Diptera: Tephritidae) from a Mass-Reared, Genetic Sexing Strain in Guatemala. J. Econ. Entomol. 2003, 96, 1132–1141. [Google Scholar] [CrossRef]
- Blay, S.; Yuval, B. Nutritional Correlates of Reproductive Success of Male Mediterranean Fruit Flies (Diptera: Tephritidae). Anim. Behav. 1997, 54, 59–66. [Google Scholar] [CrossRef]
- Taylor, P.W.; Yuval, B. Postcopulatory Sexual Selection in Mediterranean Fruit Flies: Advantages for Large and Protein-Fed Males. Anim. Behav. 1999, 58, 247–254. [Google Scholar] [CrossRef]
- Leftwich, P.T.; Nash, W.J.; Friend, L.A.; Chapman, T. Adaptation to Divergent Larval Diets in the Medfly, Ceratitis capitata. Evolution 2017, 71, 289–303. [Google Scholar] [CrossRef] [PubMed]
- Chan, H.T., Jr.; Hansen, J.D.; Tam, S.Y.T. Larval Diets from Different Protein Sources for Mediterranean Fruit Flies (Diptera: Tephritidae). J. Econ. Entomol. 1990, 83, 1954–1958. [Google Scholar] [CrossRef]
- Leftwich, P.T.; Nash, W.J.; Friend, L.A.; Chapman, T. Contribution of Maternal Effects to Dietary Selection in Mediterranean Fruit Flies. Evolution 2019, 73, 278–292. [Google Scholar] [CrossRef] [PubMed]
- Roriz, A.K.P.; Joachim-Bravo, I.S. The Relevance of Age and Nutritional Status on the Mating Competitiveness of Medfly Males (Diptera: Teprhitidae). Zoologia (Curitiba) 2013, 30, 506–512. [Google Scholar] [CrossRef]
- Shelly, T.E.; Edu, J.; Pahio, E. Female Medflies Mate Selectively with Young Males but Gain No Apparent Fitness Benefits. J. Insect Behav. 2011, 24, 55–66. [Google Scholar] [CrossRef]
- Werren, J.H. Biology of Wolbachia. Annu. Rev. Entomol. 1997, 42, 587–609. [Google Scholar] [CrossRef]
- Zug, R.; Hammerstein, P. Still a Host of Hosts for Wolbachia: Analysis of Recent Data Suggests That 40% of Terrestrial Arthropod Species Are Infected. PLoS ONE 2012, 7, e38544. [Google Scholar] [CrossRef]
- Zhang, K.-J.; Han, X.; Hong, X.-Y. Various Infection Status and Molecular Evidence for Horizontal Transmission and Recombination of Wolbachia and Cardinium among Rice Planthoppers and Related Species. Insect Sci. 2013, 20, 329–344. [Google Scholar] [CrossRef]
- Turelli, M.; Cooper, B.S.; Richardson, K.M.; Ginsberg, P.S.; Peckenpaugh, B.; Antelope, C.X.; Kim, K.J.; May, M.R.; Abrieux, A.; Wilson, D.A.; et al. Rapid Global Spread of wRi-like Wolbachia across Multiple Drosophila. Curr. Biol. 2018, 28, 963–971.e8. [Google Scholar] [CrossRef]
- Weeks, A.R.; Breeuwer, J.A.J. Wolbachia–Induced Parthenogenesis in a Genus of Phytophagous Mites. Proc. R. Soc. London. Ser. B Biol. Sci. 2001, 268, 2245–2251. [Google Scholar] [CrossRef]
- Stouthamer, R.; Kazmer, D.J. Cytogenetics of Microbe-Associated Parthenogenesis and Its Consequences for Gene Flow in Trichogramma Wasps. Heredity 1994, 73, 317–327. [Google Scholar] [CrossRef]
- Hiroki, M.; Kato, Y.; Kamito, T.; Miura, K. Feminization of Genetic Males by a Symbiotic Bacterium in a Butterfly, Eurema hecabe (Lepidoptera: Pieridae). Naturwissenschaften 2002, 89, 167–170. [Google Scholar] [CrossRef]
- Negri, I.; Pellecchia, M.; Mazzoglio, P.J.; Patetta, A.; Alma, A. Feminizing Wolbachia in Zyginidia pullula (Insecta, Hemiptera), a Leafhopper with an XX/X0 Sex-Determination System. Proc. R. Soc. B Biol. Sci. 2006, 273, 2409–2416. [Google Scholar] [CrossRef] [PubMed]
- Hurst, G.D.; Johnson, A.P.; Schulenburg, J.H.G.; Fuyama, Y. Male-Killing Wolbachia in Drosophila: A Temperature-Sensitive Trait with a Threshold Bacterial Density. Genetics 2000, 156, 699–709. [Google Scholar] [CrossRef] [PubMed]
- Fialho, R.F.; Stevens, L. Male-Killing Wolbachia in a Flour Beetle. Proc. R. Soc. London. Ser. B Biol. Sci. 2000, 267, 1469–1473. [Google Scholar] [CrossRef] [PubMed]
- Jiggins, F.M.; Hurst, G.D.D.; Schulenburg, J.H.G.V.D.; Majerus, M.E.N. Two Male-Killing Wolbachia Strains Coexist within a Population of the Butterfly Acraea encedon. Heredity 2001, 86, 161–166. [Google Scholar] [CrossRef]
- Zeh, D.W.; Zeh, J.A.; Bonilla, M.M. Wolbachia, Sex Ratio Bias and Apparent Male Killing in the Harlequin Beetle Riding Pseudoscorpion. Heredity 2005, 95, 41–49. [Google Scholar] [CrossRef]
- Bourtzis, K.; O’Neill, S. Wolbachia Infections and Arthropod Reproduction: Wolbachia Can Cause Cytoplasmic Incompatibility, Parthenogenesis, and Feminization in Many Arthropods. BioScience 1998, 48, 287–293. [Google Scholar] [CrossRef]
- Werren, J.H.; Baldo, L.; Clark, M.E. Wolbachia: Master Manipulators of Invertebrate Biology. Nat. Rev. Microbiol. 2008, 6, 741–751. [Google Scholar] [CrossRef]
- Yen, J.H.; Barr, A.R. The Etiological Agent of Cytoplasmic Incompatibility in Culex pipiens. J. Invertebr. Pathol. 1973, 22, 242–250. [Google Scholar] [CrossRef]
- O’Neill, S.L.; Karr, T.L. Bidirectional Incompatibility between Conspecific Populations of Drosophila simulans. Nature 1990, 348, 178–180. [Google Scholar] [CrossRef] [PubMed]
- Zabalou, S.; Apostolaki, A.; Livadaras, I.; Franz, G.; Robinson, A.S.; Savakis, C.; Bourtzis, K. Incompatible Insect Technique: Incompatible Males from a Ceratitis capitata Genetic Sexing Strain. Entomol. Exp. Et Appl. 2009, 132, 232–240. [Google Scholar] [CrossRef]
- Pietri, J.E.; DeBruhl, H.; Sullivan, W. The Rich Somatic Life of Wolbachia. MicrobiologyOpen 2016, 5, 923–936. [Google Scholar] [CrossRef] [PubMed]
- Strunov, A.; Schneider, D.I.; Albertson, R.; Miller, W.J. Restricted Distribution and Lateralization of Mutualistic Wolbachia in the Drosophila Brain. Cell. Microbiol. 2017, 19, e12639. [Google Scholar] [CrossRef] [PubMed]
- Vale, P.F.; Jardine, M.D. Sex-Specific Behavioural Symptoms of Viral Gut Infection and Wolbachia in Drosophila melanogaster. J. Insect Physiol. 2015, 82, 28–32. [Google Scholar] [CrossRef]
- Bi, J.; Sehgal, A.; Williams, J.A.; Wang, Y.-F. Wolbachia Affects Sleep Behavior in Drosophila melanogaster. J. Insect Physiol. 2018, 107, 81–88. [Google Scholar] [CrossRef]
- Morioka, E.; Oida, M.; Tsuchida, T.; Ikeda, M. Nighttime Activities and Peripheral Clock Oscillations Depend on Wolbachia Endosymbionts in Flies. Sci. Rep. 2018, 8, 15432. [Google Scholar] [CrossRef]
- Turley, A.P.; Moreira, L.A.; O’Neill, S.L.; McGraw, E.A. Wolbachia Infection Reduces Blood-Feeding Success in the Dengue Fever Mosquito, Aedes aegypti. PLoS Negl. Trop. Dis. 2009, 3, e516. [Google Scholar] [CrossRef]
- Evans, O.; Caragata, E.P.; McMeniman, C.J.; Woolfit, M.; Green, D.C.; Williams, C.R.; Franklin, C.E.; O’Neill, S.L.; McGraw, E.A. Increased Locomotor Activity and Metabolism of Aedes aegypti Infected with a Life-Shortening Strain of Wolbachia pipientis. J. Exp. Biol. 2009, 212, 1436–1441. [Google Scholar] [CrossRef]
- Hague, M.T.J.; Woods, H.A.; Cooper, B.S. Pervasive Effects of Wolbachia on Host Activity. Biol. Lett. 2021, 17, 20210052. [Google Scholar] [CrossRef]
- Bi, J.; Zheng, Y.; Wang, R.-F.; Ai, H.; Haynes, P.R.; Brownlie, J.C.; Yu, X.-Q.; Wang, Y.-F. Wolbachia Infection May Improve Learning and Memory Capacity of Drosophila by Altering Host Gene Expression through MicroRNA. Insect Biochem. Mol. Biol. 2019, 106, 47–54. [Google Scholar] [CrossRef] [PubMed]
- Templé, N.; Richard, F.-J. Intra-Cellular Bacterial Infections Affect Learning and Memory Capacities of an Invertebrate. Front. Zool. 2015, 12, 36. [Google Scholar] [CrossRef] [PubMed]
- Hedges, L.M.; Brownlie, J.C.; O’Neill, S.L.; Johnson, K.N. Wolbachia and Virus Protection in Insects. Science 2008, 322, 702. [Google Scholar] [CrossRef] [PubMed]
- Teixeira, L.; Ferreira, Á.; Ashburner, M. The Bacterial Symbiont Wolbachia Induces Resistance to RNA Viral Infections in Drosophila melanogaster. PLoS Biol. 2008, 6, e1000002. [Google Scholar] [CrossRef]
- Kambris, Z.; Cook, P.E.; Phuc, H.K.; Sinkins, S.P. Immune Activation by Life-Shortening Wolbachia and Reduced Filarial Competence in Mosquitoes. Science 2009, 326, 134–136. [Google Scholar] [CrossRef]
- Moreira, L.A.; Iturbe-Ormaetxe, I.; Jeffery, J.A.; Lu, G.; Pyke, A.T.; Hedges, L.M.; Rocha, B.C.; Hall-Mendelin, S.; Day, A.; Riegler, M.; et al. A Wolbachia Symbiont in Aedes aegypti Limits Infection with Dengue, Chikungunya, and Plasmodium. Cell 2009, 139, 1268–1278. [Google Scholar] [CrossRef]
- Hughes, G.L.; Koga, R.; Xue, P.; Fukatsu, T.; Rasgon, J.L. Wolbachia Infections Are Virulent and Inhibit the Human Malaria Parasite Plasmodium falciparum in Anopheles gambiae. PLoS Pathog. 2011, 7, e1002043. [Google Scholar] [CrossRef]
- Ye, Y.H.; Woolfit, M.; Rancès, E.; O’Neill, S.L.; McGraw, E.A. Wolbachia-Associated Bacterial Protection in the Mosquito Aedes aegypti. PLoS Negl. Trop. Dis. 2013, 7, e2362. [Google Scholar] [CrossRef]
- Martinez, J.; Longdon, B.; Bauer, S.; Chan, Y.-S.; Miller, W.J.; Bourtzis, K.; Teixeira, L.; Jiggins, F.M. Symbionts Commonly Provide Broad Spectrum Resistance to Viruses in Insects: A Comparative Analysis of Wolbachia Strains. PLoS Pathog. 2014, 10, e1004369. [Google Scholar] [CrossRef]
- Peng, Y.; Nielsen, J.E.; Cunningham, J.P.; McGraw, E.A. Wolbachia Infection Alters Olfactory-Cued Locomotion in Drosophila spp. Appl. Environ. Microbiol. 2008, 74, 3943–3948. [Google Scholar] [CrossRef]
- Bi, J.; Wang, Y.-F. The Effect of the Endosymbiont Wolbachia on the Behavior of Insect Hosts. Insect Science 2020, 27, 846–858. [Google Scholar] [CrossRef] [PubMed]
- Koukou, K.; Pavlikaki, H.; Kilias, G.; Werren, J.H.; Bourtzis, K.; Alahiotis, S.N. Influence of Antibiotic Treatment and Wolbachia Curing on Sexual Isolation Among Drosophila melanogaster Cage Populations. Evolution 2006, 60, 87–96. [Google Scholar] [CrossRef] [PubMed]
- Miller, W.J.; Ehrman, L.; Schneider, D. Infectious Speciation Revisited: Impact of Symbiont-Depletion on Female Fitness and Mating Behavior of Drosophila paulistorum. PLoS Pathog. 2010, 6, e1001214. [Google Scholar] [CrossRef] [PubMed]
- Zhao, D.-X.; Zhang, X.-F.; Chen, D.-S.; Zhang, Y.-K.; Hong, X.-Y. Wolbachia-Host Interactions: Host Mating Patterns Affect Wolbachia Density Dynamics. PLoS ONE 2013, 8, e66373. [Google Scholar] [CrossRef] [PubMed]
- Rohrscheib, C.E.; Bondy, E.; Josh, P.; Riegler, M.; Eyles, D.; van Swinderen, B.; Weible, M.W.; Brownlie, J.C. Wolbachia Influences the Production of Octopamine and Affects Drosophila Male Aggression. Appl. Environ. Microbiol. 2015, 81, 4573–4580. [Google Scholar] [CrossRef]
- De Crespigny, F.E.C.; Pitt, T.D.; Wedell, N. Increased Male Mating Rate in Drosophila Is Associated with Wolbachia Infection. J. Evol. Biol. 2006, 19, 1964–1972. [Google Scholar] [CrossRef]
- Panteleev, D.I.; Goriacheva, I.I.; Andrianov, B.V.; Reznik, N.L.; Lazebnyĭ, O.E.; Kulikov, A.M. The endosymbiotic bacterium Wolbachia enhances the nonspecific resistance to insect pathogens and alters behavior of Drosophila melanogaster. Genetika 2007, 43, 1277–1280. [Google Scholar] [CrossRef]
- Schneider, D.I.; Ehrman, L.; Engl, T.; Kaltenpoth, M.; Hua-Van, A.; Le Rouzic, A.; Miller, W.J. Symbiont-Driven Male Mating Success in the Neotropical Drosophila paulistorum Superspecies. Behav. Genet. 2019, 49, 83–98. [Google Scholar] [CrossRef]
- Liu, C.; Wang, J.-L.; Zheng, Y.; Xiong, E.-J.; Li, J.-J.; Yuan, L.-L.; Yu, X.-Q.; Wang, Y.-F. Wolbachia-Induced Paternal Defect in Drosophila Is Likely by Interaction with the Juvenile Hormone Pathway. Insect Biochem. Mol. Biol. 2014, 49, 49–58. [Google Scholar] [CrossRef]
- He, Z.; Zhang, H.-B.; Li, S.-T.; Yu, W.-J.; Biwot, J.; Yu, X.-Q.; Peng, Y.; Wang, Y.-F. Effects of Wolbachia Infection on the Postmating Response in Drosophila melanogaster. Behav. Ecol. Sociobiol. 2018, 72, 146. [Google Scholar] [CrossRef]
- Blomquist, G.J.; Bagnères, A.-G. Insect Hydrocarbons: Biology, Biochemistry, and Chemical Ecology; Cambridge University Press: Cambridge, UK, 2010; ISBN 978-0-521-89814-0. [Google Scholar]
- Chung, H.; Carroll, S.B. Wax, Sex and the Origin of Species: Dual Roles of Insect Cuticular Hydrocarbons in Adaptation and Mating. BioEssays 2015, 37, 822–830. [Google Scholar] [CrossRef] [PubMed]
- Dembeck, L.M.; Böröczky, K.; Huang, W.; Schal, C.; Anholt, R.R.H.; Mackay, T.F.C. Genetic Architecture of Natural Variation in Cuticular Hydrocarbon Composition in Drosophila melanogaster. eLife 2015, 4, e09861. [Google Scholar] [CrossRef] [PubMed]
- Baião, G.C.; Schneider, D.I.; Miller, W.J.; Klasson, L. The Effect of Wolbachia on Gene Expression in Drosophila paulistorum and Its Implications for Symbiont-Induced Host Speciation. BMC Genom. 2019, 20, 465. [Google Scholar] [CrossRef]
- Richard, F.-J. Symbiotic Bacteria Influence the Odor and Mating Preference of Their Hosts. Front. Ecol. Evol. 2017, 5, 143. [Google Scholar] [CrossRef]
- Fortin, M.; Debenest, C.; Souty-Grosset, C.; Richard, F.-J. Males Prefer Virgin Females, Even If Parasitized, in the Terrestrial Isopod Armadillidium vulgare. Ecol. Evol. 2018, 8, 3341–3353. [Google Scholar] [CrossRef] [PubMed]
- Boller, E.F.; Russ, K.; Vallo, V.; Bush, G.L. Incompatible Races of European Cherry Fruit Fly, Rhagoletis cerasi (Dipter a: Tephritidae), Their Origin and Potential Use in Biological Control. Entomol. Exp. Et Appl. 1976, 20, 237–247. [Google Scholar] [CrossRef]
- Bourtzis, K.; Nirgianaki, A.; Onyango, P.; Savakis, C. A Prokaryotic DnaA Sequence in Drosophila melanogasten Wolbachia Infection and Cytoplasmic Incompatibility among Laboratory Strains. Insect Mol. Biol. 1994, 3, 131–142. [Google Scholar] [CrossRef]
- Rocha, L.S.; Mascarenhas, R.O.; Perondini, A.L.P.; Selivon, D. Occurrence of Wolbachia in Brazilian Samples of Ceratitis capitata (Wiedemann) (Diptera: Tephritidae). Neotrop. Entomol. 2005, 34, 1013–1015. [Google Scholar] [CrossRef]
- Zabalou, S.; Riegler, M.; Theodorakopoulou, M.; Stauffer, C.; Savakis, C.; Bourtzis, K. Wolbachia-Induced Cytoplasmic Incompatibility as a Means for Insect Pest Population Control. Proc. Natl. Acad. Sci. USA 2004, 101, 15042–15045. [Google Scholar] [CrossRef]
- Bourtzis, K.; Robinson, A.S. Insect Pest Control Using Wolbachia and/or Radiation. In Insect Symbiosis, Volume 2; Bourtzis, K., Miller, T.A., Eds.; CRC Press (Taylor and Francis Group, LLC): Boca Raton, FL, USA, 2006; pp. 225–246. ISBN 978-0-429-12721-2. [Google Scholar]
- Sarakatsanou, A.; Diamantidis, A.D.; Papanastasiou, S.A.; Bourtzis, K.; Papadopoulos, N.T. Effects of Wolbachia on Fitness of the Mediterranean Fruit Fly (Diptera: Tephritidae). J. Appl. Entomol. 2011, 135, 554–563. [Google Scholar] [CrossRef]
- Kyritsis, G.A. Effect of Endosymbiotic Microorganisms on the Behavior and Biology of the Mediterranean Fruit Fly (Diptera: Tephritidae). Ph.D. Thesis, University of Thessaly, Volos, Greek, 2016. [Google Scholar]
- Kyritsis, G.A.; Augustinos, A.A.; Livadaras, I.; Cáceres, C.; Bourtzis, K.; Papadopoulos, N.T. Medfly-Wolbachia Symbiosis: Genotype x Genotype Interactions Determine Host’s Life History Traits under Mass Rearing Conditions. BMC Biotechnol. 2019, 19, 96. [Google Scholar] [CrossRef] [PubMed]
- Dionysopoulou, N.K.; Papanastasiou, S.A.; Kyritsis, G.A.; Papadopoulos, N.T. Effect of Host Fruit, Temperature and Wolbachia Infection on Survival and Development of Ceratitis capitata Immature Stages. PLoS ONE 2020, 15, e0229727. [Google Scholar] [CrossRef] [PubMed]
- Boller, E.F. Rhagoletis cerasi and Ceratitis capitata. In Handbook of Insect Rearing; Sing, P., Moore, R., Eds.; Elsevier: Amsterdam, The Netherlands, 1985; pp. 135–144. [Google Scholar]
- Arita, L.H.; Kaneshiro, K.Y. Structure and Function of the Rectal Epithelium and Anal Glands During Mating Behavior in the Mediterranean Fruit Fly Male. Proc. Hawaiian Entomol. Soc. 1986, 26, 27–30. [Google Scholar]
- Arita, L.H.; Kaneshiro, K.Y. Sexual Selection and Lek Behavior in the Mediterranean Fruit Fly, Ceratitis capitata (Diptera: Tephritidae). Pac. Sci. 1989, 43, 135–143. [Google Scholar]
- Sokal, R.R.; Rohlf, F.J. Biometry: The Principles and Practice of Statistics in Biological Research, 3rd ed.; W.H. Freeman: New York, NY, USA, 1995; ISBN 978-0-7167-2411-7. [Google Scholar]
- Shelly, T.E.; Kennelly, S.S.; McInnis, D.O. Effect of Adult Diet on Signalling Activity, Mate Attraction, and Mating Success in Male Mediterranean Fruit Flies (Diptera: Tephritidae). Flen 2002, 85, 150–155. [Google Scholar] [CrossRef]
- Joachim-Bravo, I.S.; Anjos, C.S.; Costa, A.M. The Role of Protein in the Sexual Behaviour of Males of Ceratitis capitata (Diptera: Tephritidae): Mating Success, Copula Duration and Number of Copulations. Zoologia (Curitiba) 2009, 26, 407–412. [Google Scholar] [CrossRef]
- Newton, I.L.G.; Rice, D.W. The Jekyll and Hyde Symbiont: Could Wolbachia Be a Nutritional Mutualist? J. Bacteriol. 2020, 202, e00589-19. [Google Scholar] [CrossRef]
- Hosokawa, T.; Koga, R.; Kikuchi, Y.; Meng, X.-Y.; Fukatsu, T. Wolbachia as a Bacteriocyte-Associated Nutritional Mutualist. Proc. Natl. Acad. Sci. USA 2010, 107, 769–774. [Google Scholar] [CrossRef]
- Brownlie, J.C.; Adamski, M.; Slatko, B.; McGraw, E.A. Diversifying Selection and Host Adaptation in Two Endosymbiont Genomes. BMC Evol. Biol. 2007, 7, 68. [Google Scholar] [CrossRef]
- Foster, J.; Ganatra, M.; Kamal, I.; Ware, J.; Makarova, K.; Ivanova, N.; Bhattacharyya, A.; Kapatral, V.; Kumar, S.; Posfai, J.; et al. The Wolbachia Genome of Brugia malayi: Endosymbiont Evolution within a Human Pathogenic Nematode. PLoS Biol. 2005, 3, e121. [Google Scholar] [CrossRef]
- Wu, B.; Novelli, J.; Foster, J.; Vaisvila, R.; Conway, L.; Ingram, J.; Ganatra, M.; Rao, A.U.; Hamza, I.; Slatko, B. The Heme Biosynthetic Pathway of the Obligate Wolbachia Endosymbiont of Brugia malayi as a Potential Anti-Filarial Drug Target. PLoS Negl. Trop. Dis. 2009, 3, e475. [Google Scholar] [CrossRef] [PubMed]
- Brownlie, J.C.; Cass, B.N.; Riegler, M.; Witsenburg, J.J.; Iturbe-Ormaetxe, I.; McGraw, E.A.; O’Neill, S.L. Evidence for Metabolic Provisioning by a Common Invertebrate Endosymbiont, Wolbachia pipientis, during Periods of Nutritional Stress. PLoS Pathog. 2009, 5, e1000368. [Google Scholar] [CrossRef] [PubMed]
- Kremer, N.; Voronin, D.; Charif, D.; Mavingui, P.; Mollereau, B.; Vavre, F. Wolbachia Interferes with Ferritin Expression and Iron Metabolism in Insects. PLoS Pathog. 2009, 5, e1000630. [Google Scholar] [CrossRef] [PubMed]
- Geoghegan, V.; Stainton, K.; Rainey, S.M.; Ant, T.H.; Dowle, A.A.; Larson, T.; Hester, S.; Charles, P.D.; Thomas, B.; Sinkins, S.P. Perturbed Cholesterol and Vesicular Trafficking Associated with Dengue Blocking in Wolbachia-Infected Aedes aegypti Cells. Nat. Commun. 2017, 8, 526. [Google Scholar] [CrossRef] [PubMed]
- Caragata, E.P.; Rancès, E.; O’Neill, S.L.; McGraw, E.A. Competition for Amino Acids Between Wolbachia and the Mosquito Host, Aedes aegypti. Microb. Ecol. 2014, 67, 205–218. [Google Scholar] [CrossRef]
- Behar, A.; Ben-Yosef, M.; Lauzon, C.R.; Yuval, B.; Jurkevitch, E. Structure and Function of the Bacterial Community Associated with the Mediterranean Fruit Fly. In Insect Symbiosis, Volume 3; Bourtzis, K., Miller, T.A., Eds.; CRC Press: Boca Raton, FL, USA, 2008; pp. 1–24. ISBN 978-0-429-14977-1. [Google Scholar]
- Audsley, M.D.; Seleznev, A.; Joubert, D.A.; Woolfit, M.; O’Neill, S.L.; McGraw, E.A. Wolbachia Infection Alters the Relative Abundance of Resident Bacteria in Adult Aedes aegypti Mosquitoes, but Not Larvae. Mol. Ecol. 2018, 27, 297–309. [Google Scholar] [CrossRef]
- Simhadri, R.K.; Fast, E.M.; Guo, R.; Schultz, M.J.; Vaisman, N.; Ortiz, L.; Bybee, J.; Slatko, B.E.; Frydman, H.M. The Gut Commensal Microbiome of Drosophila melanogaster Is Modified by the Endosymbiont Wolbachia. mSphere 2017, 2, e00287-17. [Google Scholar] [CrossRef]
- Li, T.-P.; Zhou, C.-Y.; Gong, J.-T.; Xi, Z.; Hong, X.-Y. Recently Introduced Wolbachia Reduces Bacterial Species Richness and Reshapes Bacterial Community Structure in Nilaparvata lugens. Pest Manag. Sci. 2022, 78, 1881–1894. [Google Scholar] [CrossRef]
- Duan, X.-Z.; Sun, J.-T.; Wang, L.-T.; Shu, X.-H.; Guo, Y.; Keiichiro, M.; Zhu, Y.-X.; Bing, X.-L.; Hoffmann, A.A.; Hong, X.-Y. Recent Infection by Wolbachia Alters Microbial Communities in Wild Laodelphax striatellus Populations. Microbiome 2020, 8, 104. [Google Scholar] [CrossRef]
- Ami, E.B.; Yuval, B.; Jurkevitch, E. Manipulation of the Microbiota of Mass-Reared Mediterranean Fruit Flies Ceratitis capitata (Diptera: Tephritidae) Improves Sterile Male Sexual Performance. ISME J. 2010, 4, 28–37. [Google Scholar] [CrossRef]
- Ben-Yosef, M.; Jurkevitch, E.; Yuval, B. Effect of Bacteria on Nutritional Status and Reproductive Success of the Mediterranean Fruit Fly Ceratitis capitata. Physiol. Entomol. 2008, 33, 145–154. [Google Scholar] [CrossRef]
- Darrington, M.; Leftwich, P.T.; Holmes, N.A.; Friend, L.A.; Clarke, N.V.E.; Worsley, S.F.; Margaritopolous, J.T.; Hogenhout, S.A.; Hutchings, M.I.; Chapman, T. Characterisation of the Symbionts in the Mediterranean Fruit Fly Gut. Microb. Genom. 2022, 8, 000801. [Google Scholar] [CrossRef] [PubMed]
- Cappelli, A.; Petrelli, D.; Gasperi, G.; Serrao, A.G.M.; Ricci, I.; Damiani, C.; Favia, G. Bacterial Symbionts in Ceratitis capitata. Insects 2022, 13, 474. [Google Scholar] [CrossRef]
- Bel Mokhtar, N.; Catalá-Oltra, M.; Stathopoulou, P.; Asimakis, E.; Remmal, I.; Remmas, N.; Maurady, A.; Britel, M.R.; García de Oteyza, J.; Tsiamis, G.; et al. Dynamics of the Gut Bacteriome During a Laboratory Adaptation Process of the Mediterranean Fruit Fly, Ceratitis capitata. Front. Microbiol. 2022, 13, 919760. [Google Scholar] [CrossRef]
- Kittayapong, P.; Kaeothaisong, N.; Ninphanomchai, S.; Limohpasmanee, W. Combined Sterile Insect Technique and Incompatible Insect Technique: Sex Separation and Quality of Sterile Aedes aegypti Male Mosquitoes Released in a Pilot Population Suppression Trial in Thailand. Parasites Vectors 2018, 11, 657. [Google Scholar] [CrossRef] [PubMed]
- Kittayapong, P.; Ninphanomchai, S.; Limohpasmanee, W.; Chansang, C.; Chansang, U.; Mongkalangoon, P. Combined Sterile Insect Technique and Incompatible Insect Technique: The First Proof-of-Concept to Suppress Aedes aegypti Vector Populations in Semi-Rural Settings in Thailand. PLoS Negl. Trop. Dis. 2019, 13, e0007771. [Google Scholar] [CrossRef]
- Zheng, X.; Zhang, D.; Li, Y.; Yang, C.; Wu, Y.; Liang, X.; Liang, Y.; Pan, X.; Hu, L.; Sun, Q.; et al. Incompatible and Sterile Insect Techniques Combined Eliminate Mosquitoes. Nature 2019, 572, 56–61. [Google Scholar] [CrossRef]
- Nikolouli, K.; Sassù, F.; Mouton, L.; Stauffer, C.; Bourtzis, K. Combining Sterile and Incompatible Insect Techniques for the Population Suppression of Drosophila suzukii. J. Pest. Sci. 2020, 93, 647–661. [Google Scholar] [CrossRef]
- Crawford, J.E.; Clarke, D.W.; Criswell, V.; Desnoyer, M.; Cornel, D.; Deegan, B.; Gong, K.; Hopkins, K.C.; Howell, P.; Hyde, J.S.; et al. Efficient Production of Male Wolbachia-Infected Aedes aegypti Mosquitoes Enables Large-Scale Suppression of Wild Populations. Nat. Biotechnol. 2020, 38, 482–492. [Google Scholar] [CrossRef]



{kind=link}
{kind=link}
| Source of Variation | df | MS | F | p | Partial Eta Squared |
|---|---|---|---|---|---|
| Infection | 1 | 79.0 | 110.8 | <0.001 | 0.755 |
| Food | 1 | 17.2 | 24.1 | <0.001 | 0.401 |
| Infection × Food | 1 | 4.6 | 6.5 | <0.001 | 0.152 |
| Error (between subjects) | 36 | 0.714 | |||
| Age | 8 | 67.4 | 402.8 | <0.001 | 0.918 |
| Age × Infection | 8 | 1.8 | 10.8 | <0.001 | 0.231 |
| Age × Food | 8 | 0.4 | 2.3 | 0.024 | 0.059 |
| Age × Infection × Food | 8 | 0.2 | 1.0 | 0.449 | 0.027 |
| Error (Age) | 288 | 0.167 |
| Source of Variation | df | MS | F | p | Partial Eta Squared |
|---|---|---|---|---|---|
| Infection | 1 | 100.9 | 31.7 | <0.001 | 0.469 |
| Food | 1 | 27.0 | 8.5 | 0.006 | 0.191 |
| Infection × Food | 1 | 4.44 | 1.4 | 0.245 | 0.037 |
| Error (between subjects) | 36 | 3.182 | |||
| Time of day | 8 | 194.9 | 133.5 | <0.001 | 0.788 |
| Time of day × Infection | 8 | 5.7 | 3.9 | <0.001 | 0.098 |
| Time of day × Food | 8 | 16.9 | 11.6 | <0.001 | 0.243 |
| Time of day × Infection × Food | 8 | 0.8 | 0.6 | 0.790 | 0.016 |
| Error (Time of day) | 288 | 1.460 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Kyritsis, G.A.; Koskinioti, P.; Bourtzis, K.; Papadopoulos, N.T. Effect of Wolbachia Infection and Adult Food on the Sexual Signaling of Males of the Mediterranean Fruit Fly Ceratitis capitata. Insects 2022, 13, 737. https://doi.org/10.3390/insects13080737
Kyritsis GA, Koskinioti P, Bourtzis K, Papadopoulos NT. Effect of Wolbachia Infection and Adult Food on the Sexual Signaling of Males of the Mediterranean Fruit Fly Ceratitis capitata. Insects. 2022; 13(8):737. https://doi.org/10.3390/insects13080737
Chicago/Turabian StyleKyritsis, Georgios A., Panagiota Koskinioti, Kostas Bourtzis, and Nikos T. Papadopoulos. 2022. "Effect of Wolbachia Infection and Adult Food on the Sexual Signaling of Males of the Mediterranean Fruit Fly Ceratitis capitata" Insects 13, no. 8: 737. https://doi.org/10.3390/insects13080737