
Comparative Analysis of Epicuticular Lipids in Locusta migratoria and Calliptamus italicus: A Possible Role in Susceptibility to Entomopathogenic Fungi

 ,
, {kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:Simple Summary
Abstract
1. Introduction
2. Methods
2.1. Insects and Fungi
2.2. Cuticular Lipid Extraction and Derivatization
2.3. GC-MS Analysis
2.4. Susceptibility to Fungal Infections
2.5. Adhesion and Germination Assays
2.6. Statistics
3. Results
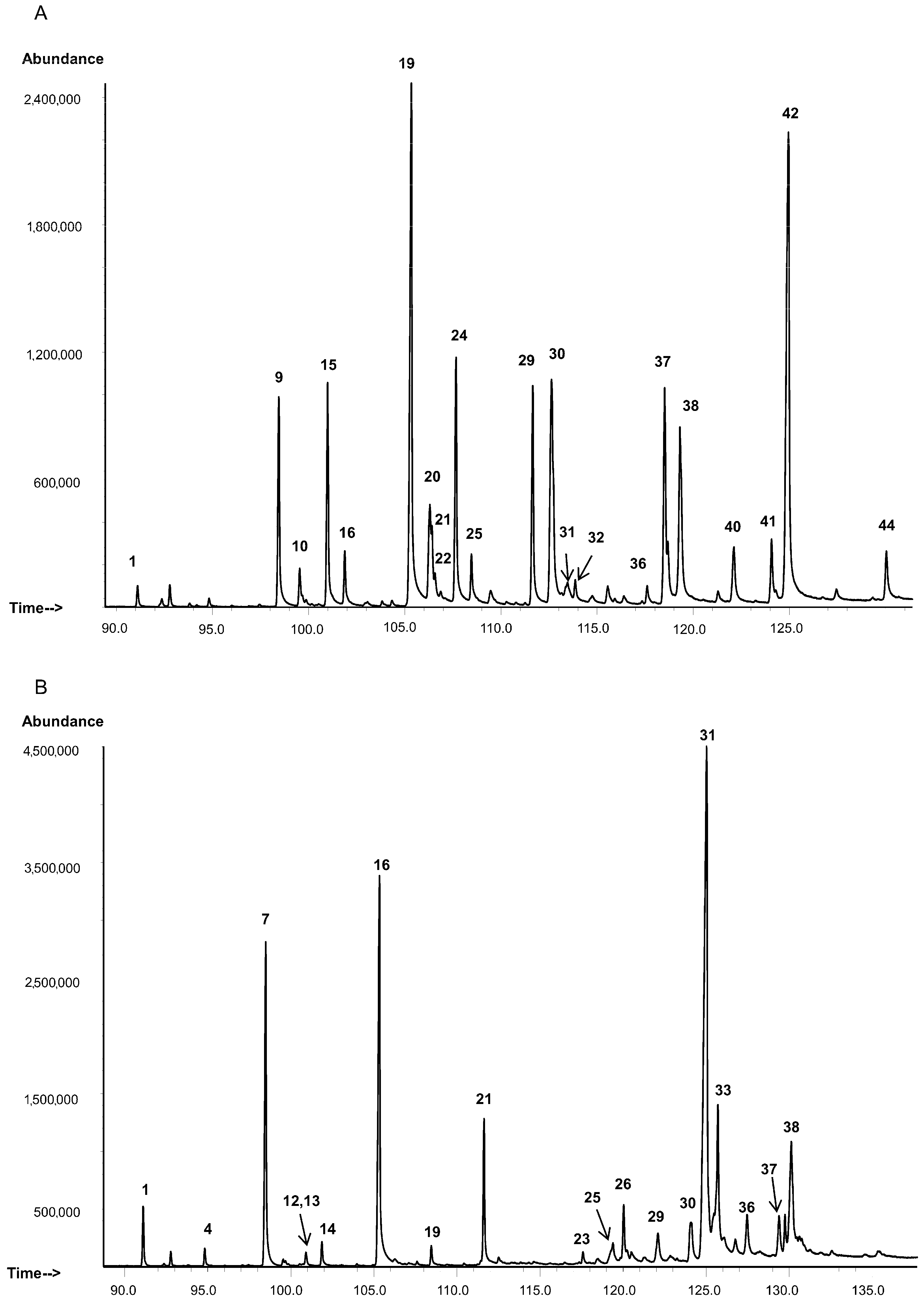
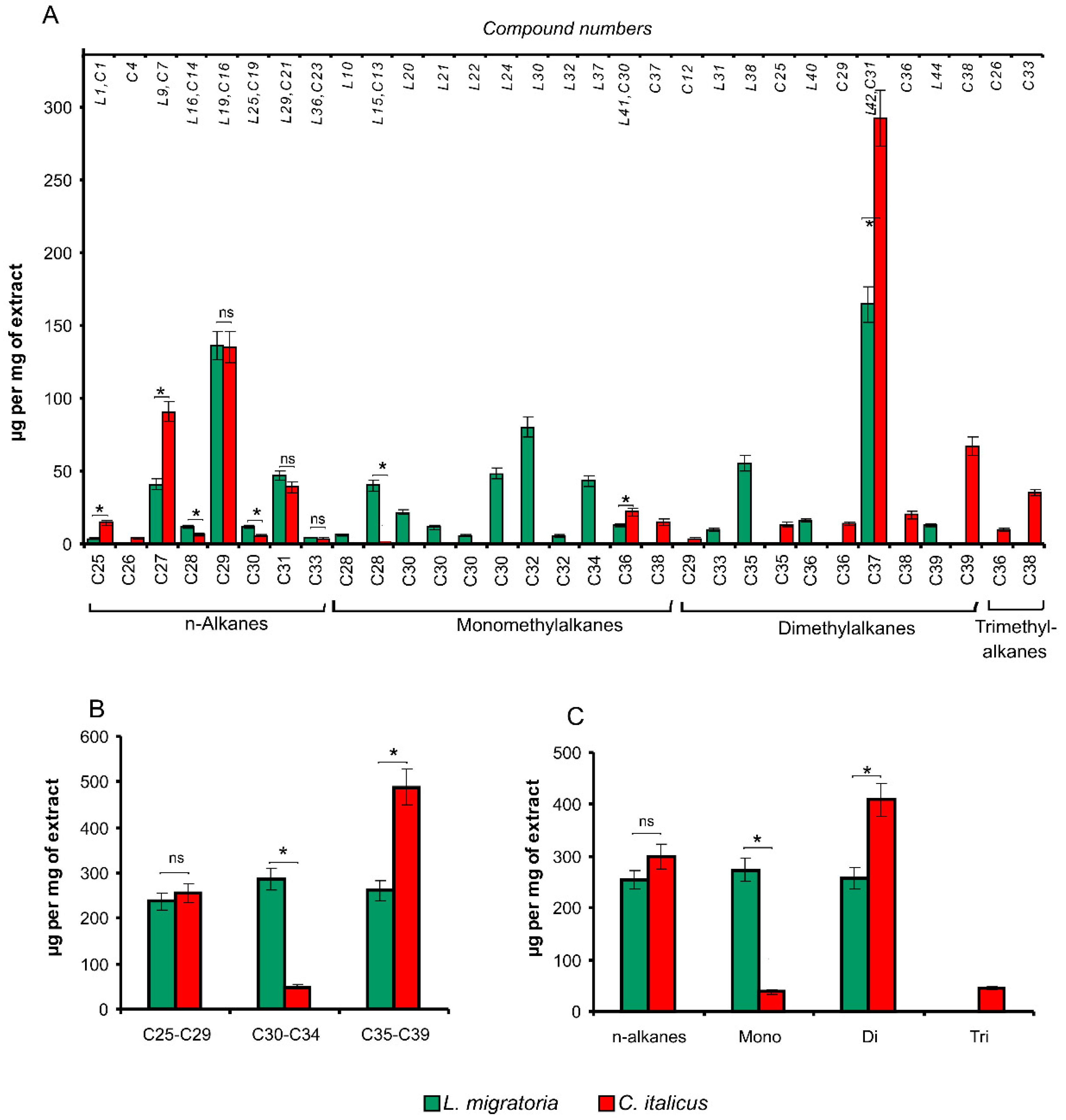
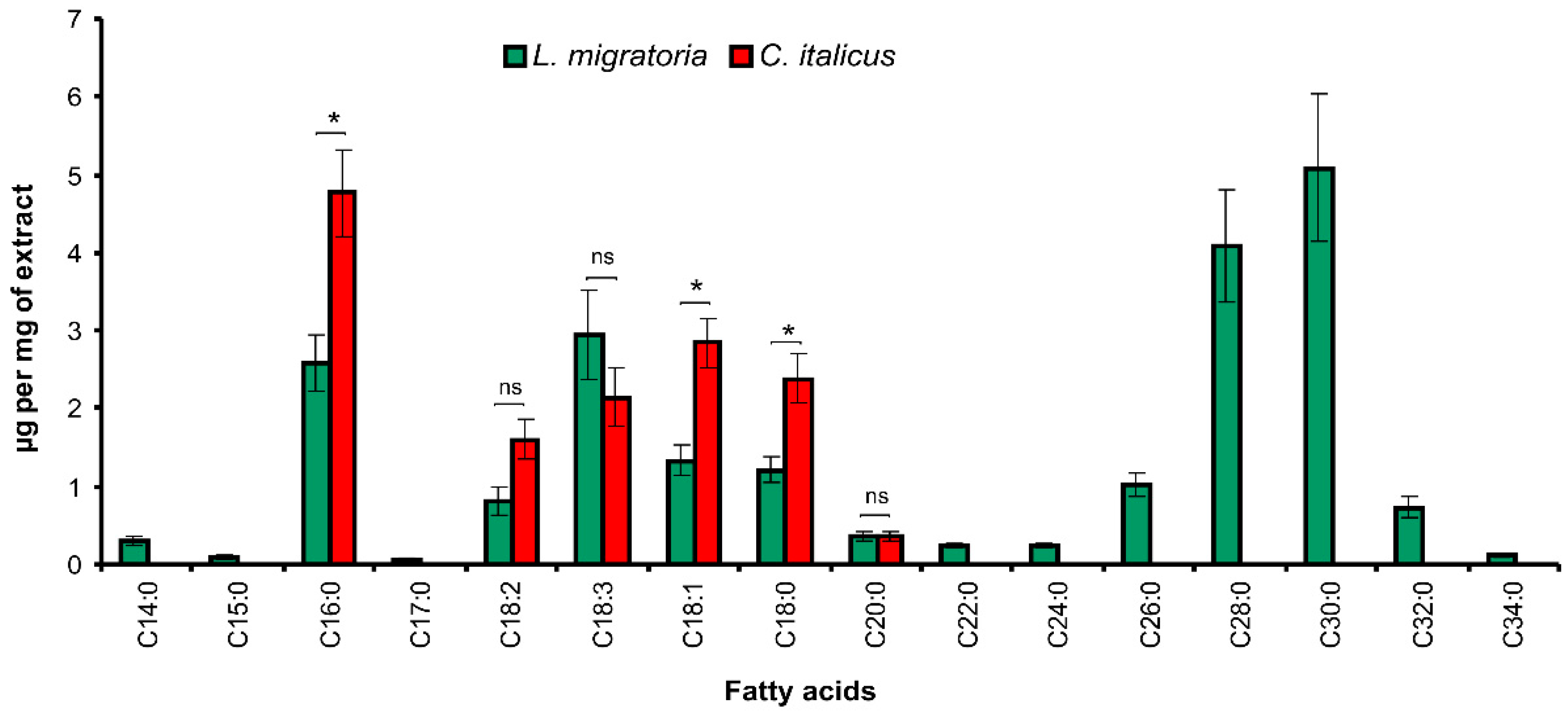
3.1. Cuticular Lipids
3.1.1. Hydrocarbons
3.1.2. Fatty Acids
3.1.3. Other Lipids
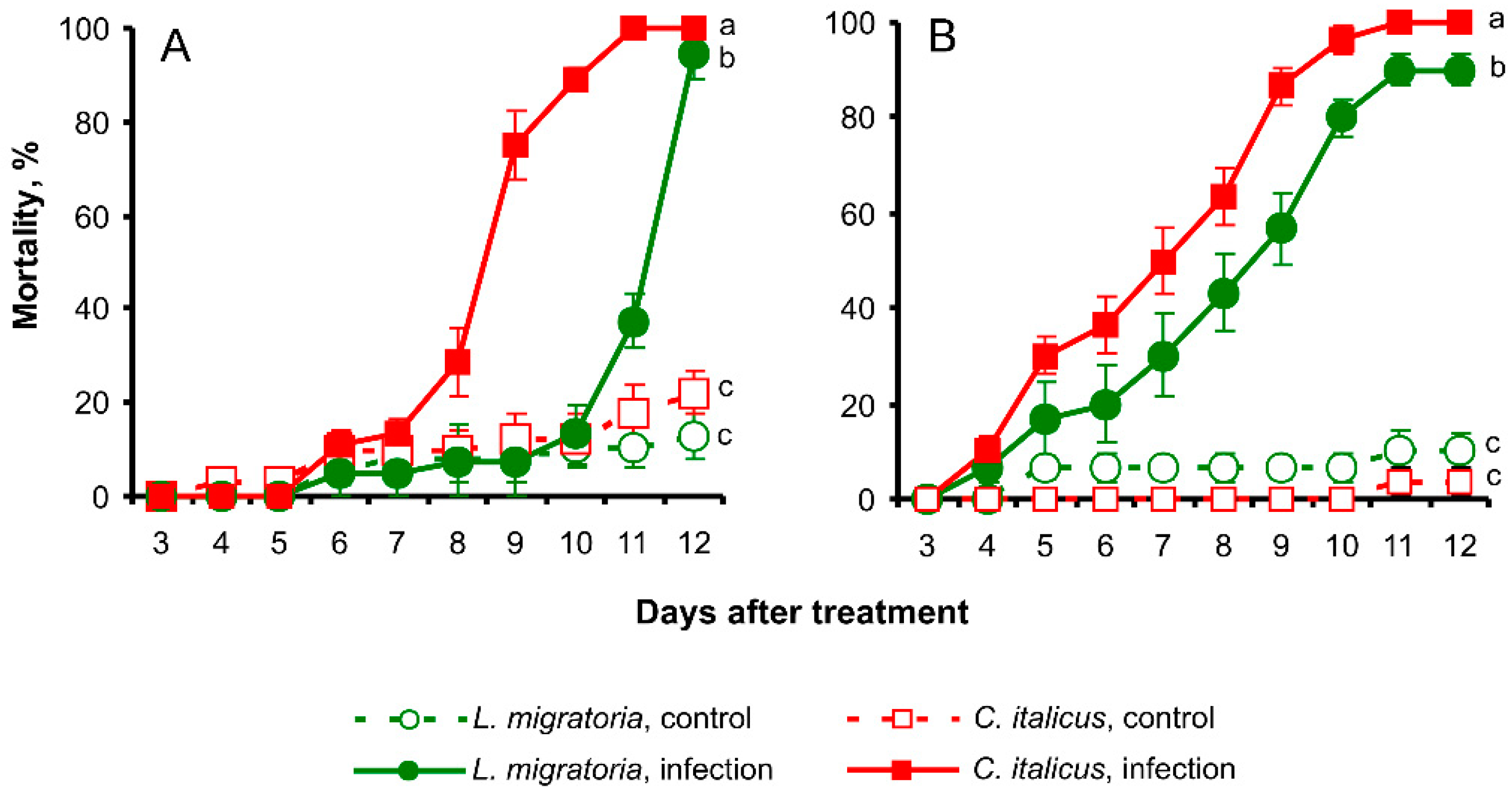
3.2. Susceptibility to Fungal Infections
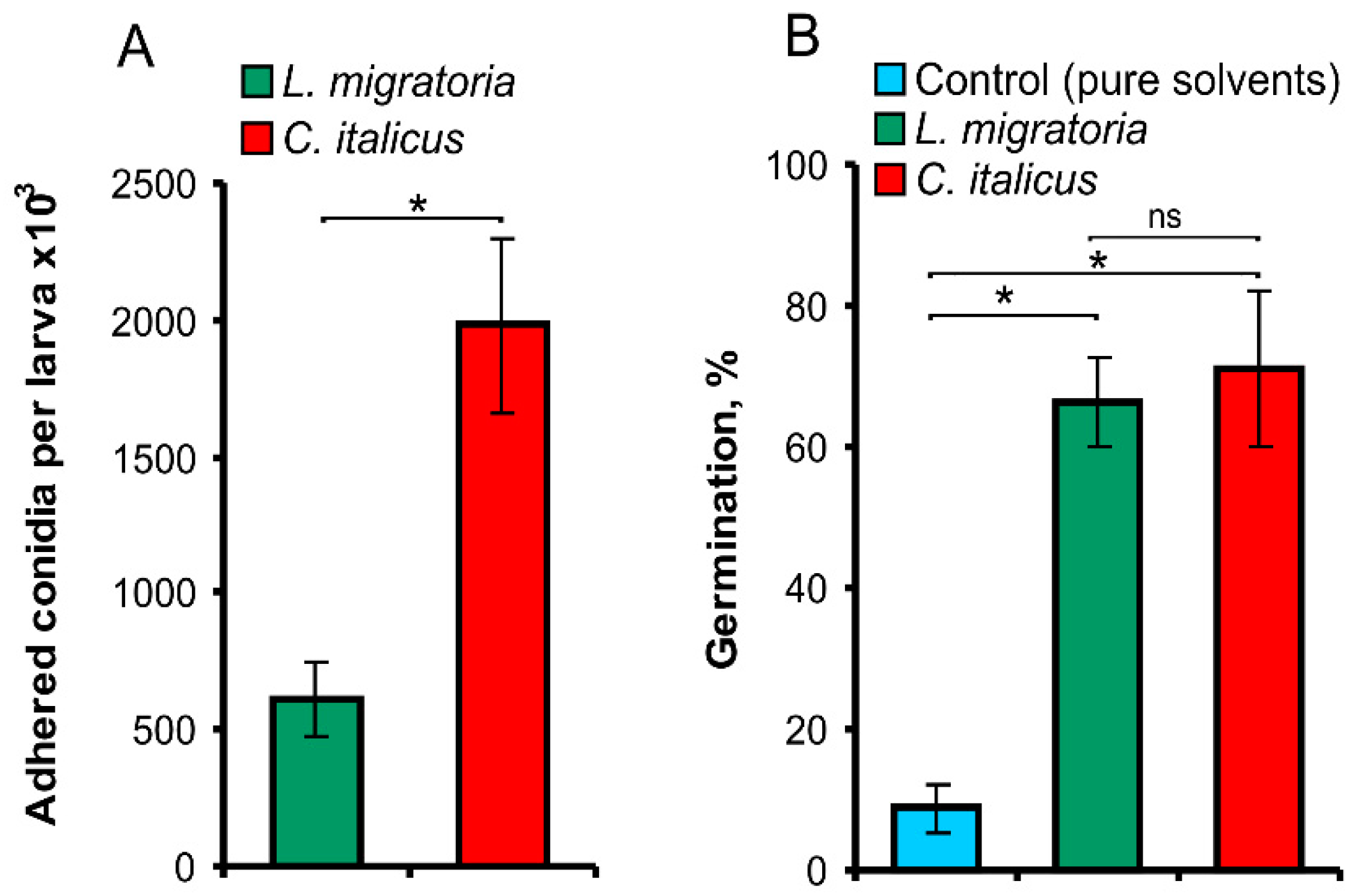
3.3. The Adhesion and Germination Assays
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Howard, R.W.; Blomquist, G.J. Ecological, behavioral, and biochemical aspects of insect hydrocarbons. Annu. Rev. Entomol. 2005, 50, 371–393. [Google Scholar] [CrossRef]
- Blomquist, G.J.; Bagnères, A.-G. (Eds.) Introduction: History and overview of insect hydrocarbons. In Insect Hydrocarbons Biology, Biochemistry, and Chemical Ecology, 1st ed.; Cambridge University Press: New York, NY, USA, 2010; pp. 3–18. [Google Scholar]
- Pedrini, N.; Crespo, R.; Juárez, M.P. Biochemistry of insect epicuticle degradation by entomopathogenic fungi. Comp. Biochem. Physiol. C Toxicol. Pharmacol. 2007, 146, 124–137. [Google Scholar] [CrossRef] [PubMed]
- Pedrini, N. Molecular interactions between entomopathogenic fungi (Hypocreales) and their insect host: Perspectives from stressful cuticle and hemolymph battlefields and the potential of dual RNA sequencing for future studies. Fungal Biol. 2018, 122, 538–545. [Google Scholar] [CrossRef] [PubMed]
- Keyhani, N.O. Lipid biology in fungal stress and virulence: Entomopathogenic fungi. Fungal Biol. 2018, 122, 420–429. [Google Scholar] [CrossRef] [PubMed]
- Lockey, K.H. Lipids of the insect cuticle: Origin, composition and function. Comp. Biochem. Physiol. B Comp. Biochem. 1988, 89, 595–645. [Google Scholar] [CrossRef]
- Downer, R.G.H.; Matthews, J.R. Patterns of Lipid Distribution and Utilisation in Insects. Am. Zool. 1976, 16, 733–745. [Google Scholar] [CrossRef]
- Blomquist, G. Biosynthesis of cuticular hydrocarbons. In Insect Hydrocarbons Biology, Biochemistry, and Chemical Ecology, 1st ed.; Blomquist, G.J., Bagnères, A.-G., Eds.; Cambridge University Press: New York, NY, USA, 2010; pp. 35–52. [Google Scholar] [CrossRef]
- Buckner, J. Oxygenated derivatives of hydrocarbons. In Insect Hydrocarbons Biology, Biochemistry, and Chemical Ecology, 1st ed.; Blomquist, G.J., Bagnères, A.-G., Eds.; Cambridge University Press: New York, NY, USA, 2010; pp. 187–204. [Google Scholar] [CrossRef]
- Menzel, F.; Blaimer, B.B.; Schmitt, T. How do cuticular hydrocarbons evolve? Physiological constraints and climatic and biotic selection pressures act on a complex functional trait. Proc. R. Soc. B Biol. Sci. 2017, 284, 20161727. [Google Scholar] [CrossRef]
- Hadley, N.F.; Schultz, T.D. Water loss in three species of tiger beetles (Cicindela): Correlations with epicuticular hydrocarbons. J. Insect Physiol. 1987, 33, 677–682. [Google Scholar] [CrossRef]
- McClain, E.; Seely, M.K.; Hadley, N.F.; Gray, V. Wax blooms in Tenebrionid beetles of the Namib desert: Correlations with environment. Ecology 1985, 66, 112–118. [Google Scholar] [CrossRef]
- Ramsay, J.A. The evaporation of water from the cockroach. J. Exp. Biol. 1935, 12, 373–383. [Google Scholar] [CrossRef]
- Gibbs, A.G. Lipid melting and cuticular permeability: New insights into an old problem. J. Insect Physiol. 2002, 48, 391–400. [Google Scholar] [CrossRef]
- Gibbs, A.; Pomonis, J.G. Physical properties of insect cuticular hydrocarbons: The effects of chain length, methyl-branching and unsaturation. Comp. Biochem. Physiol. B Biochem. Mol. Biol. 1995, 112, 243–249. [Google Scholar] [CrossRef]
- Patel, S.; Nelson, D.R.; Gibbs, A.G. Chemical and physical analyses of wax ester properties. J. Insect Sci. 2001, 1, 4. [Google Scholar] [CrossRef]
- Lockey, K.H.; Oraha, V.S. Cuticular lipids of adult Locusta migratoria migratoriodes (R and F), Schistocerca gregaria (Forskål) (Acrididae) and other orthopteran species—II. Hydrocarbons. Comp. Biochem. Physiol. B Biochem. Mol. Biol. 1990, 95, 721–744. [Google Scholar] [CrossRef]
- Chung, H.; Carroll, S.B. Wax, sex and the origin of species: Dual roles of insect cuticular hydrocarbons in adaptation and mating. BioEssays 2015, 37, 822–830. [Google Scholar] [CrossRef] [PubMed]
- Gibbs, A.G.; Louie, A.K.; Ayala, J.A. Effects of temperature on cuticular lipids and water balance in a desert Drosophila: Is thermal acclimation beneficial? J. Exp. Biol. 1998, 201, 71–80. [Google Scholar] [CrossRef]
- Rourke, B.C. Geographic and altitudinal variation in water balance and metabolic rate in a California grasshopper, Melanoplus sanguinipes. J. Exp. Biol. 2000, 203, 2699–2712. [Google Scholar] [CrossRef] [PubMed]
- Sergeev, M.G. Distribution patterns of grasshoppers and their kin over the Eurasian steppes. Insects 2021, 12, 77. [Google Scholar] [CrossRef] [PubMed]
- Sergeev, M.G. Ups and downs of the Italian locust (Calliptamus italicus L.) Populations in the Siberian steppes: On the horns of dilemmas. Agronomy 2021, 11, 746. [Google Scholar] [CrossRef]
- Genin, E.; Jullien, R.; Perez, F.; Fuzeau-Braesch, S. Cuticular hydrocarbons of gregarious and solitary locusts Locusta migratoria cinerascens. J. Chem. Ecol. 1986, 12, 1213–1238. [Google Scholar] [CrossRef]
- Lockey, K.H. Cuticular hydrocarbons of Locusta, Schistocerca, and Periplaneta, and their role in waterproofing. Insect Biochem. 1976, 6, 457–472. [Google Scholar] [CrossRef]
- Moore, D. A plague on locusts—The Lubilosa story. Outlooks Pest Manag. 2008, 19, 14–17. [Google Scholar] [CrossRef]
- Ortiz-Urquiza, A.; Keyhani, N.O. Action on the surface: Entomopathogenic fungi versus the insect cuticle. Insects 2013, 4, 357–374. [Google Scholar] [CrossRef]
- Sosa-Gomez, D.R.; Boucias, D.G.; Nation, J.L. Attachment of Metarhizium anisopliae to the southern green stink bug Nezara viridula cuticle and fungistatic effect of cuticular lipids and aldehydes. J. Invertebr. Pathol. 1997, 69, 31–39. [Google Scholar] [CrossRef]
- Jarrold, S.L.; Moore, D.; Potter, U.; Charnley, A.K. The contribution of surface waxes to pre-penetration growth of an entomopathogenic fungus on host cuticle. Mycol. Res. 2007, 111, 240–249. [Google Scholar] [CrossRef]
- Tomilova, O.G.; Yaroslavtseva, O.N.; Ganina, M.D.; Tyurin, M.V.; Chernyak, E.I.; Senderskiy, I.V.; Noskov, Y.A.; Polenogova, O.V.; Akhanaev, Y.B.; Kryukov, V.Y.; et al. Changes in antifungal defence systems during the intermoult period in the Colorado potato beetle. J. Insect Physiol. 2019, 116, 106–117. [Google Scholar] [CrossRef]
- Napolitano, R.; Juárez, M.P. Entomopathogenous fungi degrade epicuticular hydrocarbons of Triatoma infestans. Arch. Biochem. Biophys. 1997, 344, 208–214. [Google Scholar] [CrossRef]
- Pedrini, N.; Ortiz-Urquiza, A.; Zhang, S.; Keyhani, N.O. Targeting of insect epicuticular lipids by the entomopathogenic fungus Beauveria bassiana: Hydrocarbon oxidation within the context of a host-pathogen interaction. Front. Microbiol. 2013, 4, 24. [Google Scholar] [CrossRef]
- Lecuona, R.; Riba, G.; Cassier, P.; Clement, J.L. Alterations of insect epicuticular hydrocarbons during infection with Beauveria bassiana or B. brongniartii. J. Invertebr. Pathol. 1991, 58, 10–18. [Google Scholar] [CrossRef]
- Gołębiowski, M.; Cerkowniak, M.; Ostachowska, A.; Naczk, A.M.; Boguś, M.I.; Stepnowski, P. Effect of Conidiobolus coronatus on the cuticular and internal lipid composition of Tettigonia viridissima males. Chem. Biodivers. 2016, 13, 982–989. [Google Scholar] [CrossRef]
- Paszkiewicz, M.; Gołębiowski, M.; Sychowska, J.; Boguś, M.I.; Włóka, E.; Stepnowski, P. The effect of the entomopathogenic fungus Conidiobolus coronatus on the composition of cuticular and internal lipids of Blatta orientalis females. Physiol. Entomol. 2016, 41, 111–120. [Google Scholar] [CrossRef]
- Gołębiowski, M.; Bojke, A.; Tkaczuk, C. Effects of the entomopathogenic fungi Metarhizium robertsii, Metarhizium flavoviride, and Isaria fumosorosea on the lipid composition of Galleria mellonella larvae. Mycologia 2021, 113, 525–535. [Google Scholar] [CrossRef]
- Saito, T.; Aoki, J. Toxicity of free fatty acids on the larval surfaces of two Lepidopterous insects towards Beauveria bassiana (Bals) Vuill and Paecilomyces fumosoroseus (Wize) Brown Et Smith (Deuteromycetes, Moniliales). Appl. Entomol. Zool. 1983, 18, 225–233. [Google Scholar] [CrossRef]
- Ment, D.; Gindin, G.; Rot, A.; Eshel, D.; Teper-Bamnolker, P.; Ben-Ze’ev, I.; Glazer, I.; Samish, M. Role of cuticular lipids and water-soluble compounds in tick susceptibility to Metarhizium infection. Biocontr. Sci. Technol. 2013, 23, 956–967. [Google Scholar] [CrossRef]
- Wrońska, A.K.; Boguś, M.I.; WøoÂka, E.; Kazek, M.; Kaczmarek, A.; Zalewska, K. Cuticular fatty acids of Galleria mellonella (Lepidoptera) inhibit fungal enzymatic activities of pathogenic Conidiobolus coronatus. PLoS ONE 2018, 13, e0192715. [Google Scholar] [CrossRef]
- Kaczmarek, A.; Boguś, M.I.; Włóka, E.; Wrońska, A.K.; Krawiel, A.; Kazek, M.; Zalewska, K.; Kłocińska-Biały, K.; Sobocińska, M.; Gliniewicz, A.; et al. The interaction between cuticle free fatty acids (FFAs) of the cockroaches Blattella germanica and Blatta orientalis and hydrolases produced by the entomopathogenic fungus Conidiobolus coronatus. PLoS ONE 2020, 15, e0235785. [Google Scholar] [CrossRef]
- Kryukov, V.Y.; Lednev, G.R.; Dubovskiy, I.M.; Serebrov, V.V.; Levchenko, M.V.; Khodyrev, V.P.; Sagitov, A.O.; Glupov, V.V. New approaches to the biological control of insects by entomopathogenic Hyphomycetes (Deuteromycota, Hyphomycetes). Euroasian Entomol. J. 2007, 6, 195–204. (In Russian) [Google Scholar]
- Koukos, D.; Meletiou-Christou, M.-S.; Rhizopoulou, S. Leaf surface wettability and fatty acid composition of Arbutus unedo and Arbutus andrachne grown under ambient conditions in a natural macchia. Acta Bot. Gall. 2015, 162, 225–232. [Google Scholar] [CrossRef]
- Nelson, D.R.; Adams, T.S.; Fatland, C.L. Hydrocarbons in the surface wax of eggs and adults of the Colorado potato beetle, Leptinotarsa decemlineata. Comp. Biochem. Physiol. B Biochem. Mol. Biol 2003, 134, 447–466. [Google Scholar] [CrossRef]
- Ruther, J.; Döring, M.; Steiner, S. Cuticular hydrocarbons as contact sex pheromone in the parasitoid Dibrachys cavus. Entomol. Exp. Appl. 2011, 140, 59–68. [Google Scholar] [CrossRef]
- Ment, D.; Gindin, G.; Rot, A.; Soroker, V.; Glazer, I.; Barel, S.; Samish, M. Novel technique for quantifying adhesion of Metarhizium anisopliae conidia to the tick cuticle. Appl. Environ. Microbiol. 2010, 76, 3521–3528. [Google Scholar] [CrossRef]
- Hammer, Ø.; Harper, D.A.T.; Ryan, P.D. PAST: Paleontological statistics software package for education and data analysis. Palaeontol. Electron. 2001, 4, 1–9. [Google Scholar]
- Taylor, A.R.; Roubal, W.T.; Varanasi, U. Effects of structural variation in β-monoglycerides and other lipids on ordering in synthetic membranes. Lipids 1975, 10, 535–541. [Google Scholar] [CrossRef]
- Duranton, J.-F.; Lecoq, M. Le Criquet Pèlerin au Sahel; Ministère des Affaires Etrangères des Rayas-Bas et CIRAD/PRIFAS: Montpellier, France, 1990; pp. 1–183. [Google Scholar]
- Hamilton, A.G. The relation of humidity and temperature to the development of three species of african locusts—Locusta migratoria migratorioides (R. & F.), Schistocerca gregaria (FORSK.), Nomadacris septemfasciata (SERV.). Trans. R. Ent. Soc. Lond. 1936, 85, 1–60. [Google Scholar] [CrossRef]
- Hamilton, A.G. Further studies on the relation of humidity and temperature to the development of two species of african locusts—Locusta migratoria migratorioides (R. & F.) and Schistocerca gregaria (forsk.). Trans. R. Ent. Soc. Lond. 1950, 101, 1–58. [Google Scholar] [CrossRef]
- Sergeev, M.G.; Childebaev, M.K.; Van’kova, I.A.; Gapparov, F.A.; Kambulin, V.E.; Kokanova, E.O.; Latchininskiy, A.V.; Pshenitsyna, L.B.; Temreshev, I.I.; Cernijakhovskij, M.E.; et al. Italian locust Calliptamus italicus (Linnaeus, 1758). Morphology, Ecology, Distribution, Population Menegment; FAO: Rome, Italy, 2022; pp. 1–333, (In Russian). [Google Scholar] [CrossRef]
- Latchininsky, A.V.; Sergeev, M.G.; Childebaev, M.K.; Chernijakhovskij, M.E.; Lockwood, J.A.; Kambulin, V.E.; Gapparov, F.A. Grasshoppers of Kazakhstan, Middle Asia, and the Adjacent Territories; Association for Applied Acridology International; Wyoming University: Laramie, WY, USA, 2002; pp. 1–387. (In Russian) [Google Scholar]
- Grunshaw, J.P.; Guermouche, H.; Guermouche, S.; Jago, N.D.; Jullien, R.; Knowles, E.; Perez, F. Chemical taxonomic studies of cuticular hydrocarbons in locusts of the Schistocerca americana complex (Acrididae: Cyrtacanthacridinae): Chemical relationships between new world and old world species. J. Chem. Ecol. 1990, 16, 2835–2858. [Google Scholar] [CrossRef]
- Vötsch, W.; Nicholson, G.; Müller, R.; Stierhof, Y.-D.; Gorb, S.; Schwarz, U. Chemical composition of the attachment pad secretion of the locust Locusta migratoria. Insect Biochem. Mol. Biol. 2002, 32, 1605–1613. [Google Scholar] [CrossRef]
- Oraha, V.S.; Lockey, K.H. Cuticular lipids of Locusta migratoria migratorioides (R and F), Schistocerca gregaria (Forskål)(Acrididae) and other orthopteran species-I. Polar components. Comp. Biochem. Physiol. B Biochem. Mol. Biol. 1990, 95, 603–608. [Google Scholar] [CrossRef]
- Verbruggen, E.M.; Hermens, J.L.; Tolls, J. Physicochemical properties of higher nonaromatic hydrocarbons: A literature study. J. Phys. Chem. Ref. Data 2000, 29, 1435–1446. [Google Scholar] [CrossRef]
- Butt, T.M.; Coates, C.J.; Dubovskiy, I.M.; Ratcliffe, N.A. Entomopathogenic fungi: New insights into host–pathogen interactions. Adv. Genet. 2016, 94, 307–364. [Google Scholar]
- Alkhaibari, A.M.; Lord, A.M.; Maffeis, T.; Bull, J.C.; Olivares, F.L.; Samuels, R.I.; Butt, T.M. Highly specific host-pathogen interactions influence Metarhizium brunneum blastospore virulence against Culex quinquefasciatus larvae. Virulence 2018, 9, 1449–1467. [Google Scholar] [CrossRef] [PubMed]
- Tyurin, M.V.; Kryukov, V.Y.; Yaroslavtseva, O.N.; Elisaphenko, E.A.; Dubovskiy, I.M.; Glupov, V.V. Comparative analysis of the immune reactions in Colorado potato beetle larvae under development of mycoses caused by Metarhizium robertsii, M. brunneum and M. pemphigi. J. Evol. Biochem. Physiol. 2016, 52, 252–260. [Google Scholar] [CrossRef]
- Levchenko, M.V.; Kononchuk, A.G.; Gerus, A.V.; Lednev, G.R. Differential susceptibility of Locusta migratoria and Schistocerca gregaria (Orthoptera: Acrididae) to infection with entomopathogenic fungi. Plant Prot. News 2020, 103, 150–152. [Google Scholar] [CrossRef]
- Kassa, A. Development and Testing of Mycoinsecticides Based on Submerged Spores and Aerial Conidia of the Entomopathogenic Fungi Beauveria Bassiana and Metarhizium Anisopliae (Deuteromycotina: Hyphomycetes) for Control of Locusts, Grasshoppers and Storage Pests. Ph.D. Thesis, Georg-August-Universität Göttingen, Göttingen, Germany, 2003. [Google Scholar] [CrossRef]
- Heuskin, S.; Vanderplanck, M.; Bacquet, P.; Holveck, M.-J.; Kaltenpoth, M.; Engl, T.; Pels, C.; Taverne, C.; Lognay, G.; Nieberding, C.M. The composition of cuticular compounds indicates body parts, sex and age in the model butterfly Bicyclus anynana (Lepidoptera). Front. Ecol. Evol. 2014, 2, 37. [Google Scholar] [CrossRef]
- Hajek, A.E.; Eastburn, C.C. Attachment and germination of Entomophaga maimaiga conidia on host and non-host larval cuticle. J. Invertebr. Pathol. 2003, 82, 12–22. [Google Scholar] [CrossRef]





Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Ganina, M.D.; Tyurin, M.V.; Zhumatayeva, U.T.; Lednev, G.R.; Morozov, S.V.; Kryukov, V.Y. Comparative Analysis of Epicuticular Lipids in Locusta migratoria and Calliptamus italicus: A Possible Role in Susceptibility to Entomopathogenic Fungi. Insects 2022, 13, 736. https://doi.org/10.3390/insects13080736
Ganina MD, Tyurin MV, Zhumatayeva UT, Lednev GR, Morozov SV, Kryukov VY. Comparative Analysis of Epicuticular Lipids in Locusta migratoria and Calliptamus italicus: A Possible Role in Susceptibility to Entomopathogenic Fungi. Insects. 2022; 13(8):736. https://doi.org/10.3390/insects13080736
Chicago/Turabian StyleGanina, Mariya D., Maksim V. Tyurin, Ulzhalgas T. Zhumatayeva, Georgy R. Lednev, Sergey V. Morozov, and Vadim Yu. Kryukov. 2022. "Comparative Analysis of Epicuticular Lipids in Locusta migratoria and Calliptamus italicus: A Possible Role in Susceptibility to Entomopathogenic Fungi" Insects 13, no. 8: 736. https://doi.org/10.3390/insects13080736