
Heat Shock 70 kDa Protein Cognate 3 of Brown Planthopper Is Required for Survival and Suppresses Immune Response in Plants

 , and
, and 
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:Simple Summary
Abstract
1. Introduction
2. Materials and Methods
2.1. Plant Materials and Insects
2.2. Cloning and Sequencing of NlHSC70-3 Gene
2.3. Sequence Analysis of the NlHSC70-3 Gene and Its Encoding Protein
2.4. Expression Patterns of NlHSC70-3
2.5. dsRNA Synthesis and Injection to Trigger RNAi in BPH
2.6. Subcellular Localization of NlHSC70-3 in N. benthamiana and Rice Cells
2.7. Defense Gene Expression Analyses of N. benthamiana Leaves
2.8. ROS Burst in N. benthamiana
2.9. Phylogenetic Tree
2.10. Data Analysis
3. Results
3.1. Identification of NlHSC70-3 in the Rice Tissue after BPH Infestation
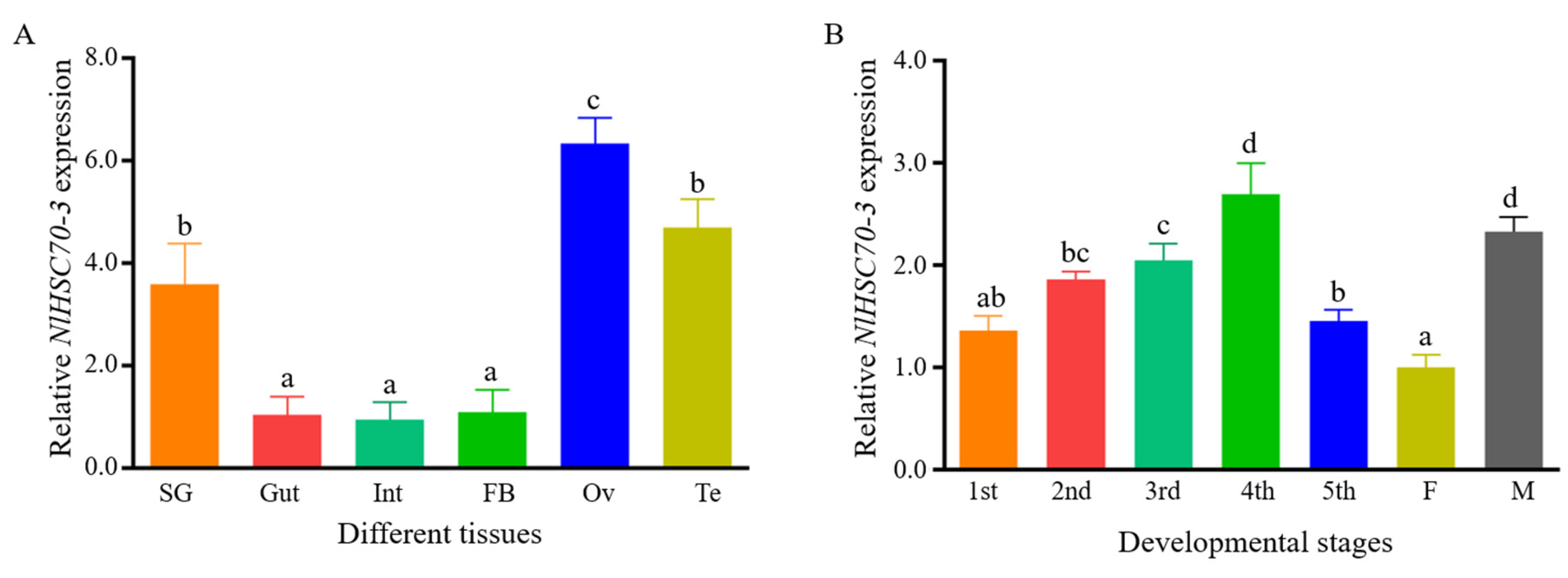
3.2. Expression Pattern of NlHSC70-3 in the BPH
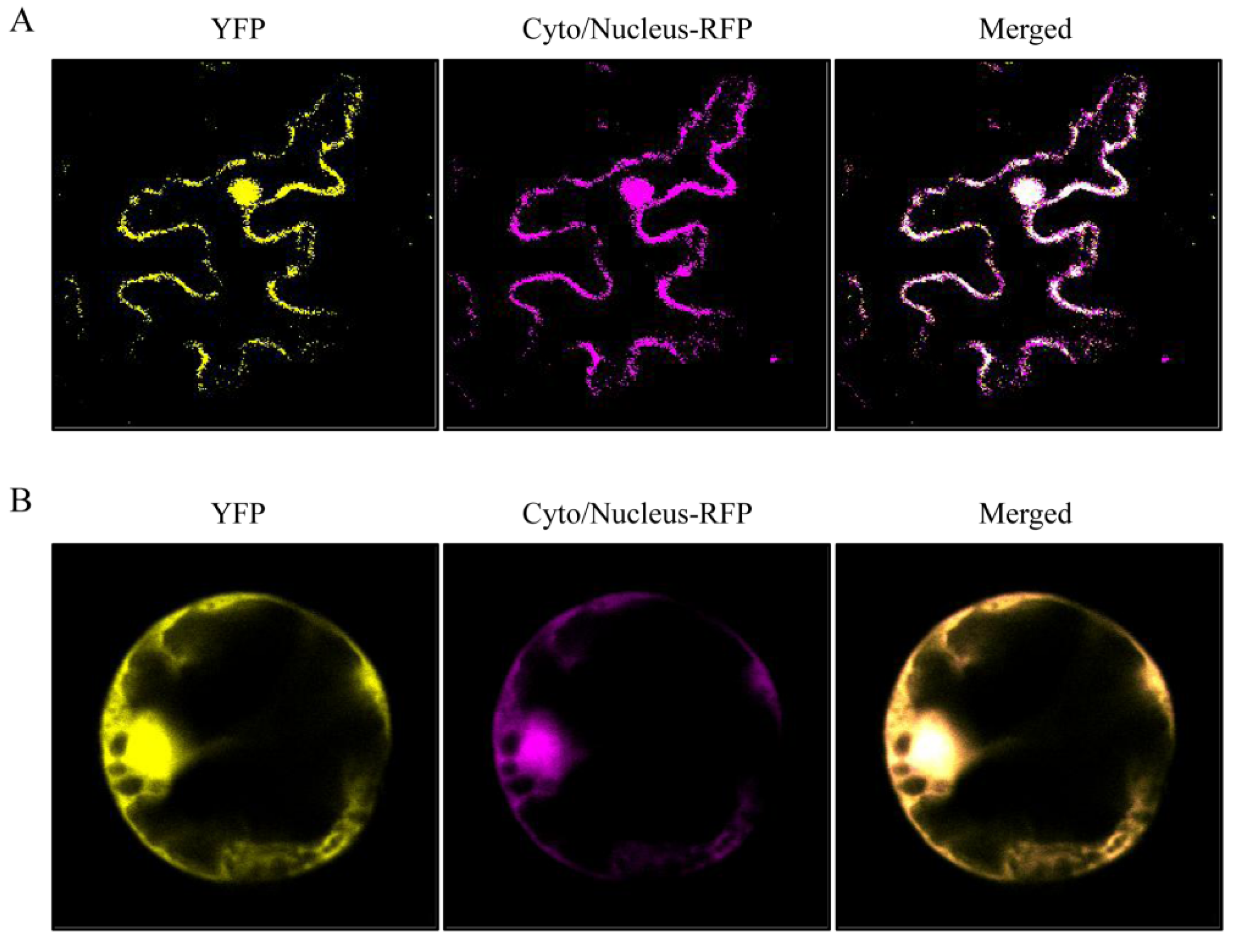
3.3. Subcellular Localization of NlHSC70-3 in Plant Cells
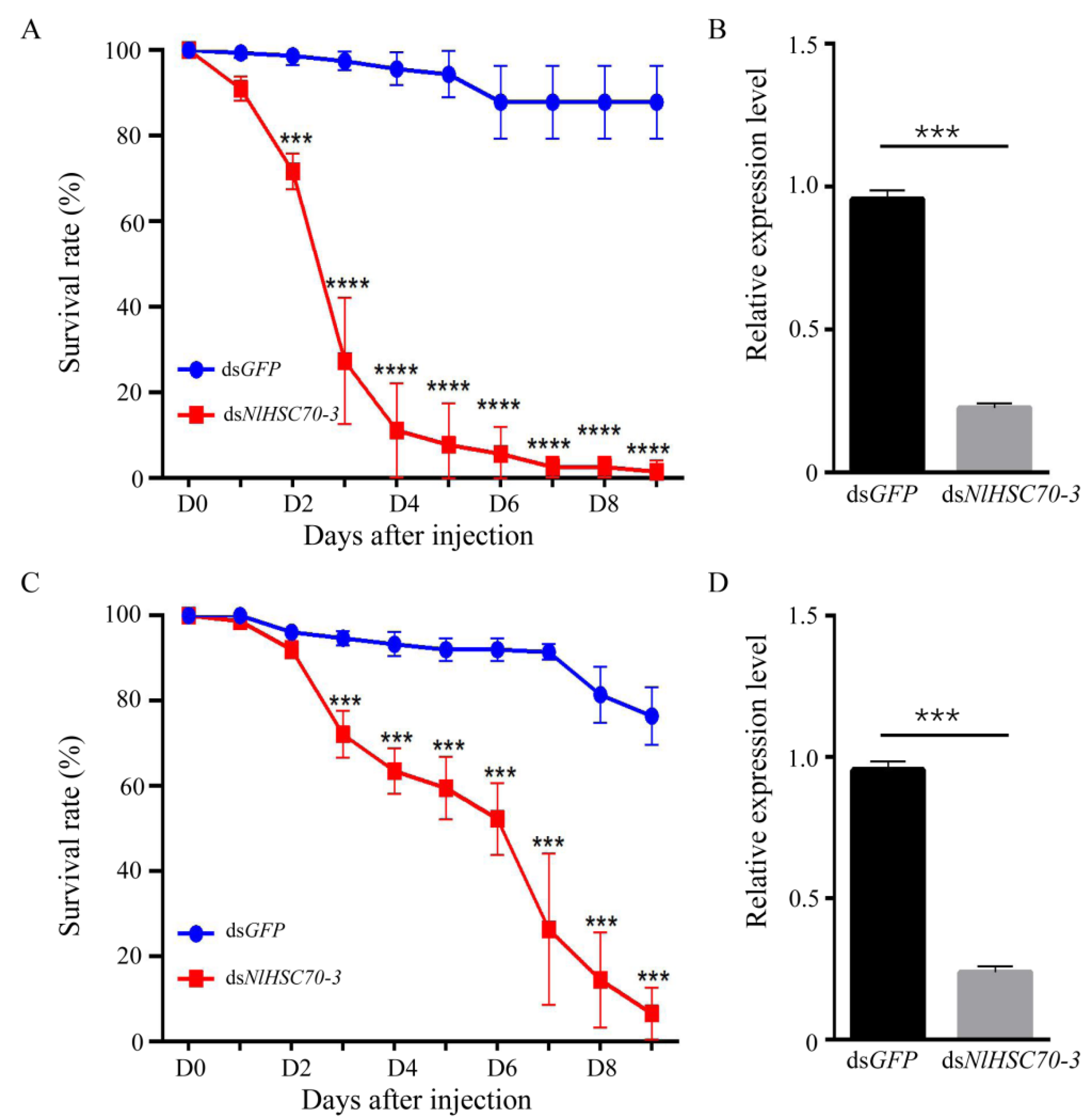
3.4. NlHSC70-3 Is Critical to Survival of BPH
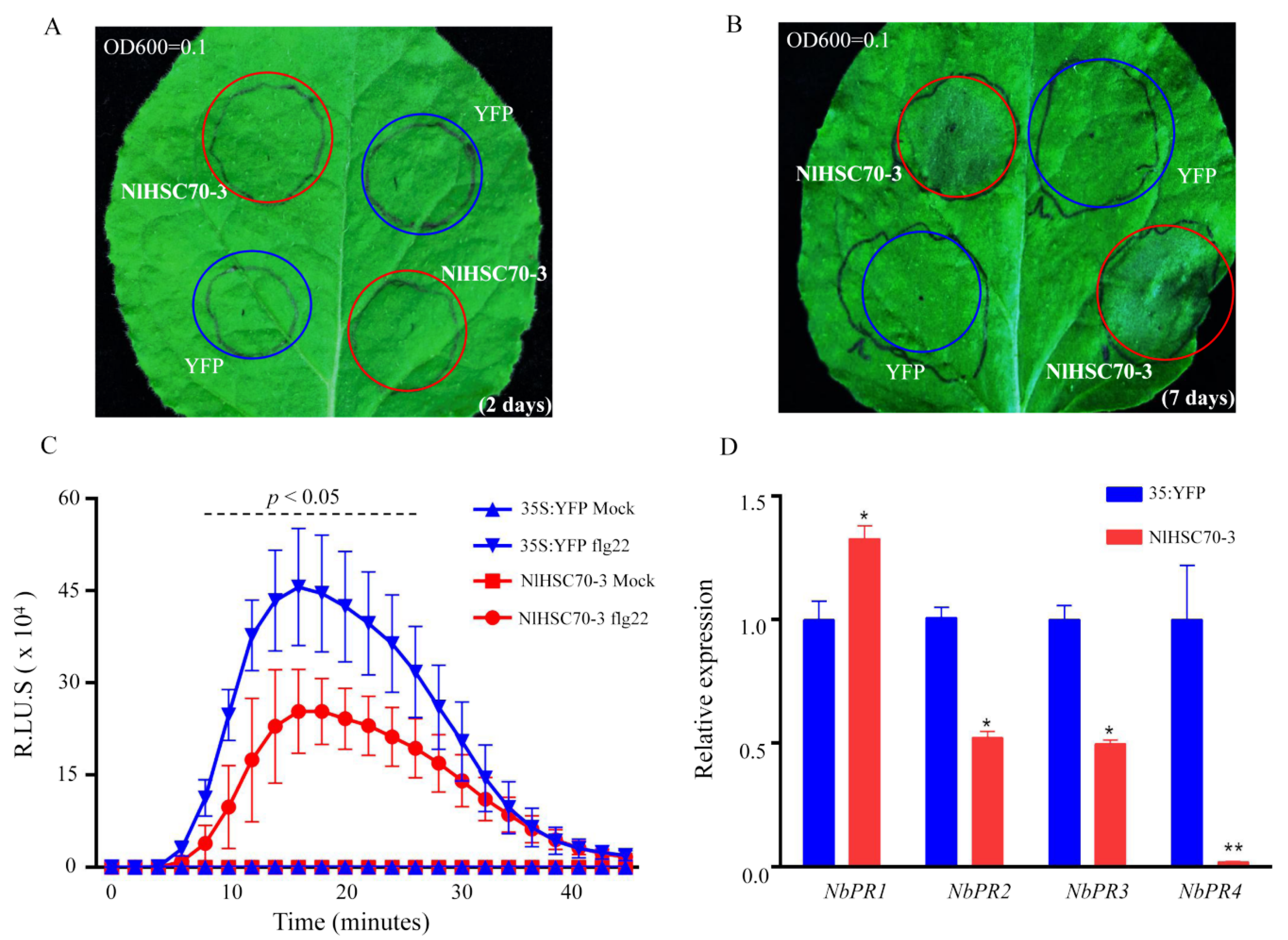
3.5. Defense Response Are Inhibited by NlHSC70-3
3.6. Sequence Analysis of NlHSC70-3 with Rice HSP70s
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Cheng, X.; Zhu, L.; He, G. Towards understanding of molecular interactions between rice and the BPH. Mol. Plant 2013, 6, 621–634. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Backus, E.A.; Serrano, M.S.; Ranger, C.M. Mechanisms of hopperburn: An overview of insect taxonomy, behavior, and physiology. Annu. Rev. Entomol. 2005, 50, 125–151. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Brar, D.S.; Virk, P.S.; Jena, K.K.; Khush, G.S. Breeding for Resistance to Planthoppers in Rice; International Rice Research Institute: Los Baños, Philippines, 2009; pp. 401–427. [Google Scholar]
- Qiu, Y.; Guo, J.; Jing, S. Development and characterization of japonica rice lines carrying the brown planthopper-resistance gene BPH12 and BPH6. Theor. Appl. Genet. 2012, 124, 485–494. [Google Scholar] [CrossRef] [PubMed]
- Hu, J.; Xiao, C.; Qing, Y.; He, Y. Recent progress on the genetics and molecular breeding of BPH resistance in rice. Rice 2016, 9, 30. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Huang, H.J.; Liu, C.W.; Huang, X.H.; Zhou, X.; Zhou, J.C.; Zhang, C.X. Screening and functional analyses of Nilaparvata lugens salivary proteome. J. Proteome Res. 2016, 15, 1883–1896. [Google Scholar] [CrossRef] [PubMed]
- Konishi, H.; Noda, H.; Tamura, Y.; Hattori, M. Proteomic analysis of the salivary glands of the rice BPH, Nilaparvata lugens (Stål) (Homoptera: Delphacidae). Appl. Entomol. Zool. 2009, 44, 525–534. [Google Scholar] [CrossRef] [Green Version]
- Sharma, A.; Khan, A.N.; Subrahmanyam, S.; Raman, A.; Taylor, G.S.; Fletcher, M.J. Salivary proteins of plant-feeding hemipteroids-implication in phytophagy. Bull. Entomol. Res. 2014, 104, 117–136. [Google Scholar] [CrossRef]
- Huang, H.J.; Zhang, C.X.; Hong, X.Y. How does saliva function in planthopper-host interactions? Arch. Insect Biochem. Physiol. 2019, 100, e21537. [Google Scholar] [CrossRef]
- Ji, R.; Yu, H.; Fu, Q.; Chen, H.; Ye, W.; Li, S.; Lou, Y. Comparative transcriptome analysis of salivary glands of two populations of rice BPH, Nilaparvata lugens, that differ in virulence. PLoS ONE 2013, 8, e79612. [Google Scholar] [CrossRef]
- Liu, X.; Zhou, H.; Zhao, J.; Hua, H.; He, Y. Identification of the secreted watery saliva proteins of the rice BPH, Nilaparvata lugens (Stål) by transcriptome and Shotgun LC–MS/MS approach. J. Insect Physiol. 2016, 89, 60–69. [Google Scholar] [CrossRef] [Green Version]
- Ji, R.; Ye, W.; Chen, H.; Zeng, J.; Li, H.; Yu, H.; Li, J.; Lou, Y. A salivary endo-β-1,4-glucanase acts as an effector that enables the BPH to feed on rice. Plant Physiol. 2017, 173, 1920–1932. [Google Scholar] [CrossRef] [PubMed]
- Ye, W.; Yu, H.; Jian, Y.; Zeng, J.; Ji, R.; Chen, H.D.; Lou, Y. A salivary EF-hand calcium-binding protein of the BPH Nilaparvata lugens functions as an effector for defense responses in rice. Sci. Rep. 2017, 7, 40498. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Shangguan, X.; Zhang, J.; Liu, B.; Zhao, Y.; Wang, H.; Wang, Z.; Guo, J.; Rao, W.; Jing, S.; Guang, W. A mucin-like protein of planthopper is required for feeding and induces immunity response in plants. Plant Physiol. 2018, 176, 552–565. [Google Scholar] [CrossRef] [Green Version]
- Huang, J.; Zhang, N.; Shan, J.H.; Peng, Y.X.; Chen, R. Salivary protein 1 of BPH is required for survival and induces immunity response in plants. Front. Plant Sci. 2020, 11, 1334. [Google Scholar] [CrossRef] [PubMed]
- Huang, H.J.; Liu, C.W.; Xu, H.J.; Bao, Y.Y.; Zhang, C.X. Mucin-like protein, a saliva component involved in brown planthopper virulence and host adaptation. J. Insect Physiol. 2017, 98, 223–230. [Google Scholar] [CrossRef]
- Ritossa, F.M. A new puffing pattern induced by temperature shock and DNP in Drosophila. Experientia 1962, 15, 571–573. [Google Scholar] [CrossRef]
- Mayer, M.P.; Bukau, B. Hsp70 chaperones: Cellular functions and molecular mechanism. Cell. Mol. Life Sci. 2005, 62, 670–684. [Google Scholar] [CrossRef] [Green Version]
- Jian, X.; Bao, Y.Y.; Li, B.L.; Cheng, Y.B.; Peng, Z.Y.; Hang, L.; Xu, H.J.; Zhu, Z.R.; Lou, Y.G.; Cheng, J.A. Transcriptome analysis of the BPH Nilaparvata lugens. PLoS ONE 2010, 5, e14233. [Google Scholar] [CrossRef] [Green Version]
- Feder, M.E.; Hofmann, G.E. Heat shock proteins, molecular chaperones, and the stress response: Evolutionary and ecological physiology. Annu. Rev. Physiol. 1999, 61, 243–282. [Google Scholar] [CrossRef] [Green Version]
- Vladimír, K.; Borovanská, T.; Hermes, L.M. The 70 kDa heat shock protein assists during the repair of chilling injury in the insect Pyrrhocoris apterus. PLoS ONE 2009, 4, e4546. [Google Scholar] [CrossRef] [Green Version]
- Dong, B.; Liu, X.Y.; Li, B.; Li, M.Y.; Li, S.G.; Liu, S. A heat shock protein protects against oxidative stress induced by lambda-cyhalothrin in the green peach aphid Myzus persicae. Pestic. Biochem. Physiol. 2021, 181, 104995. [Google Scholar] [CrossRef] [PubMed]
- Lu, K.; Chen, X.; Liu, W.T.; Zhou, Q. Characterization of heat shock cognate protein 70 gene and its differential expression in response to thermal stress between two wing morphs of Nilaparvata lugens (Stål). Comp. Biochem. Physiol. Part A Mol. Integr. Physiol. 2016, 199, 47–53. [Google Scholar] [CrossRef] [PubMed]
- Lu, K.; Chen, X.; Liu, W.; Zhang, Z.; Wang, Y.; You, K.; Li, Y.; Zhang, R.; Zhou, Q. Characterization of heat shock protein 70 transcript from Nilaparvata lugens (Stål): Its response to temperature and insecticide stresses. Biochem. Physiol. 2017, 142, 102–110. [Google Scholar] [CrossRef] [PubMed]
- Chen, X.; Li, Z.D.; Li, D.T.; Jiang, M.X.; Zhang, C.X. Hsp70/DNAJ family of genes in the brown planthopper, Nilaparvata lugens: Diversity and function. Genes 2021, 12, 394. [Google Scholar] [CrossRef] [PubMed]
- Ye, Y.X.; Zhang, H.H.; Li, D.T.; Zhuo, J.C.; Shen, Y.; Hu, Q.L.; Zhang, C.X. Chromosome-level assembly of the brown planthopper genome with a characterized Y chromosome. Mol. Ecol. Resour. 2021, 21, 1287–1298. [Google Scholar] [CrossRef] [PubMed]
- Keller, H.; Blein, J.P.; Bonnet, P. Physiological and molecular characteristics of elicitin-induced systemic acquired resistance in tobacco. Plant Physiol. 1996, 110, 365–376. [Google Scholar] [CrossRef]
- Maimbo, M.; Ohnishi, K.; Hikichi, Y. S-glycoprotein-like protein regulates defense responses in Nicotiana plants against Ralstonia solanacearum. Plant Physiol. 2010, 152, 2023–2035. [Google Scholar] [CrossRef] [Green Version]
- Han, X.; Zhang, M.H.; Yang, M.L.; Hu, Y.R. Arabidopsis JAZ proteins interact with and suppress RHD6 transcription factor to regulate jasmonate-stimulated root hair development. Plant Cell 2020, 32, 1049–1062. [Google Scholar] [CrossRef]
- Rao, W.W.; Zheng, X.H.; Liu, B.F.; Guo, Q.; Guo, J.P.; Wu, Y.; Shangguan, X.X.; Wang, H.Y.; Wang, Z.; Hu, L.; et al. Secretome analysis and in planta expression of salivary proteins identify candidate effectors from the BPH Nilaparvata lugens. Mol. Plant-Microbe Interact. 2019, 32, 227–239. [Google Scholar] [CrossRef] [Green Version]
- Wahab, M.M.S.; Akkareddy, S.; Shanthi, P.; Latha, P. Identification of differentially expressed genes under heat stress conditions in rice (Oryza sativa L.). Mol. Biol. Rep. 2020, 47, 1935–1948. [Google Scholar] [CrossRef]
- Liu, A.; Zhou, Z.; Yi, Y.; Chen, G. Transcriptome analysis reveals the roles of stem nodes in cadmium transport to rice grain. BMC Genom. 2020, 21, 127. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zhao, L.; Vecchi, G.; Vendruscolo, M.; Krner, R.; Hartl, F.U. The Hsp70 chaperone system stabilizes a thermosensitive subproteome in E. coli. Cell Rep. 2019, 28, 1335–1345. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- King, A.M.; MacRae, T.H. Insect heat shock proteins during stress and diapause. Annu. Rev. Entomol. 2015, 60, 59–75. [Google Scholar] [CrossRef] [PubMed]
- Morammazi, S.; Shokrollahi, B. The pattern of HSP70 gene expression, flight activity and temperature in Apis mellifera colonies. J. Therm. Biol. 2020, 91, 102647. [Google Scholar] [CrossRef] [PubMed]
- Chang, Y.W.; Zhang, X.X.; Chen, J.Y.; Lu, M.X.; Gong, W.R.; Du, Y.Z. Characterization of three heat shock protein 70 genes from Liriomyza trifolii and expression during thermal stress and insect development. Bull. Entomol. Res. 2018, 109, 150–159. [Google Scholar] [CrossRef] [PubMed]
- Kaloshian, I.; Walling, L.L. Hemipteran and dipteran pests: Effectors and plant host immune regulators. Integr. Plant Biol. 2016, 58, 350–361. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Jiang, Y.; Zhang, C.X.; Chen, R.; He, S.Y. Challenging battles of plants with phloem-feeding insects and prokaryotic pathogens. Proc. Natl. Acad. Sci. USA 2019, 116, 23390–23397. [Google Scholar] [CrossRef]
- Li, B.; Li, Z.; Lu, C.; Zhao, D.; Shen, G.; Kusakabe, T.; Xia, Q.; Zhao, P. Heat shock cognate 70 functions as a chaperone for the stability of kinetochore protein CENP-N in holocentric insect silkworms. Int. J. Mol. Sci. 2019, 20, 5823. [Google Scholar] [CrossRef] [Green Version]






Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Yang, H.; Zhang, X.; Li, H.; Ye, Y.; Li, Z.; Han, X.; Hu, Y.; Zhang, C.; Jiang, Y. Heat Shock 70 kDa Protein Cognate 3 of Brown Planthopper Is Required for Survival and Suppresses Immune Response in Plants. Insects 2022, 13, 299. https://doi.org/10.3390/insects13030299
Yang H, Zhang X, Li H, Ye Y, Li Z, Han X, Hu Y, Zhang C, Jiang Y. Heat Shock 70 kDa Protein Cognate 3 of Brown Planthopper Is Required for Survival and Suppresses Immune Response in Plants. Insects. 2022; 13(3):299. https://doi.org/10.3390/insects13030299
Chicago/Turabian StyleYang, Houhong, Xiaoya Zhang, Hanjing Li, Yuxuan Ye, Zhipeng Li, Xiao Han, Yanru Hu, Chuanxi Zhang, and Yanjuan Jiang. 2022. "Heat Shock 70 kDa Protein Cognate 3 of Brown Planthopper Is Required for Survival and Suppresses Immune Response in Plants" Insects 13, no. 3: 299. https://doi.org/10.3390/insects13030299