Identification and Characterization of CYP6 Family Genes from the Oriental Fruit Moth (Grapholita molesta) and Their Responses to Insecticides

 ,
, {kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:Simple Summary
Abstract
1. Introduction
2. Materials and Methods
2.1. Insect Samples
2.2. Bioinformatics Analysis
2.3. Induction of Expression by Three Kinds of Insecticides
2.4. Statistical Analysis
3. Results
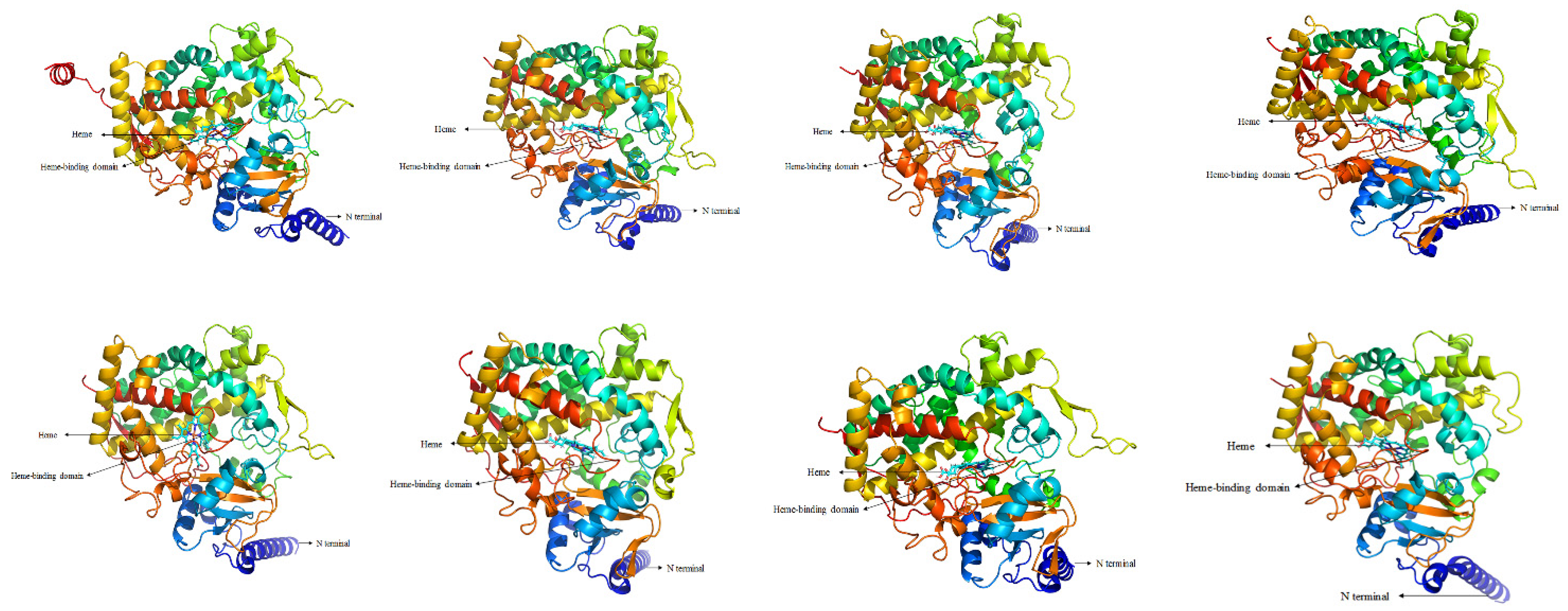
3.1. Identification and Characteristics of CYP6 Gene Members in G. molesta
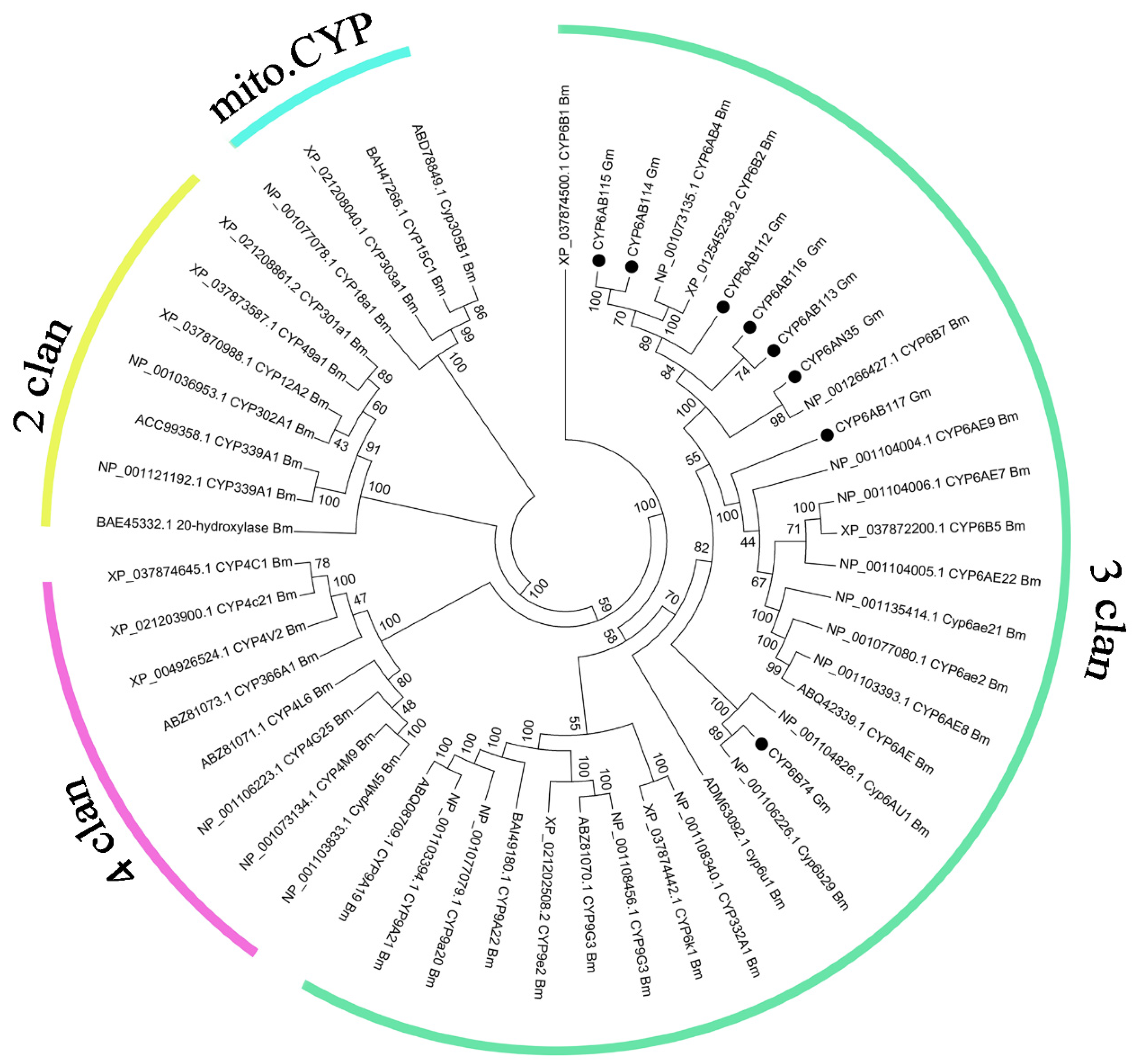
3.2. Phylogenetic Analysis
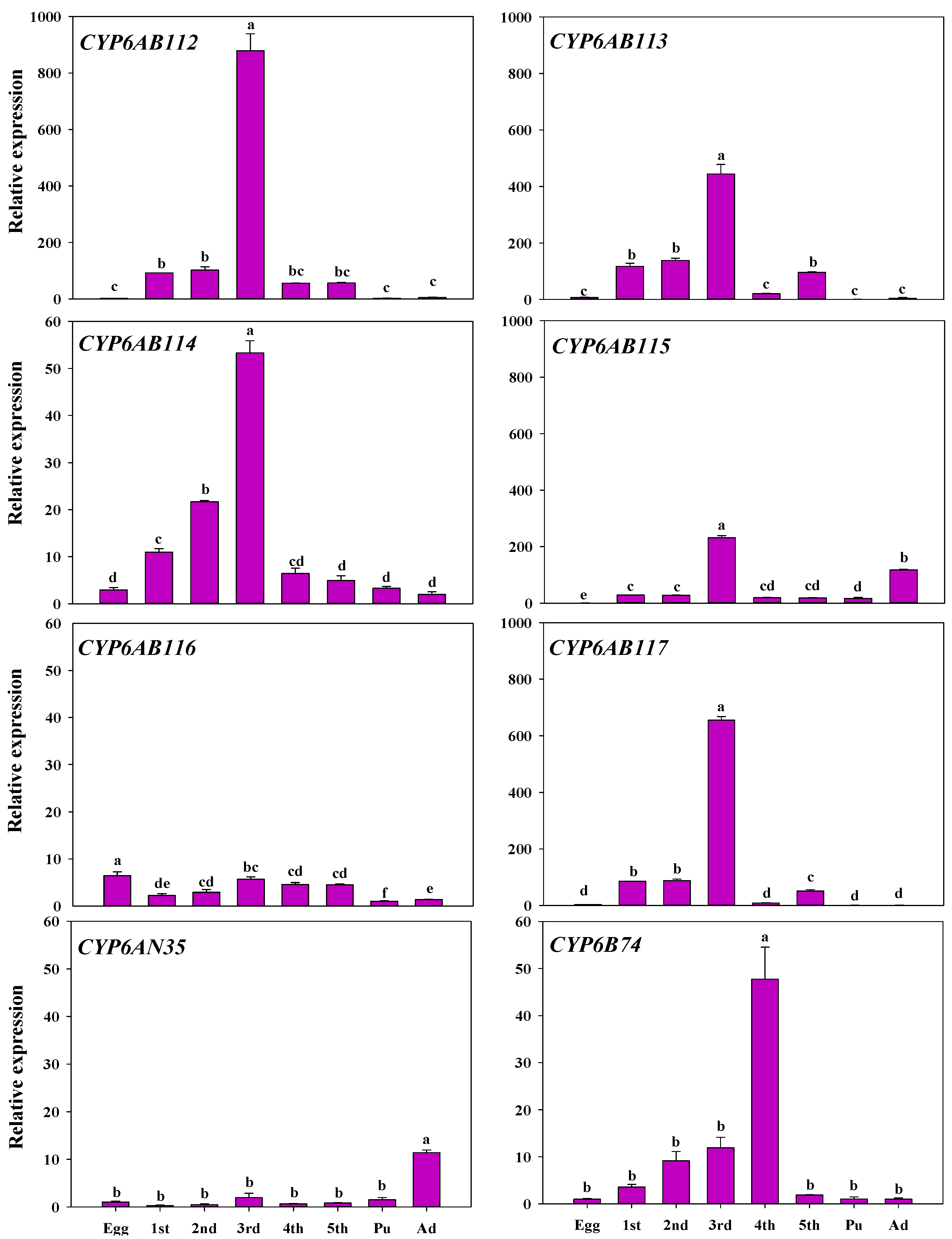
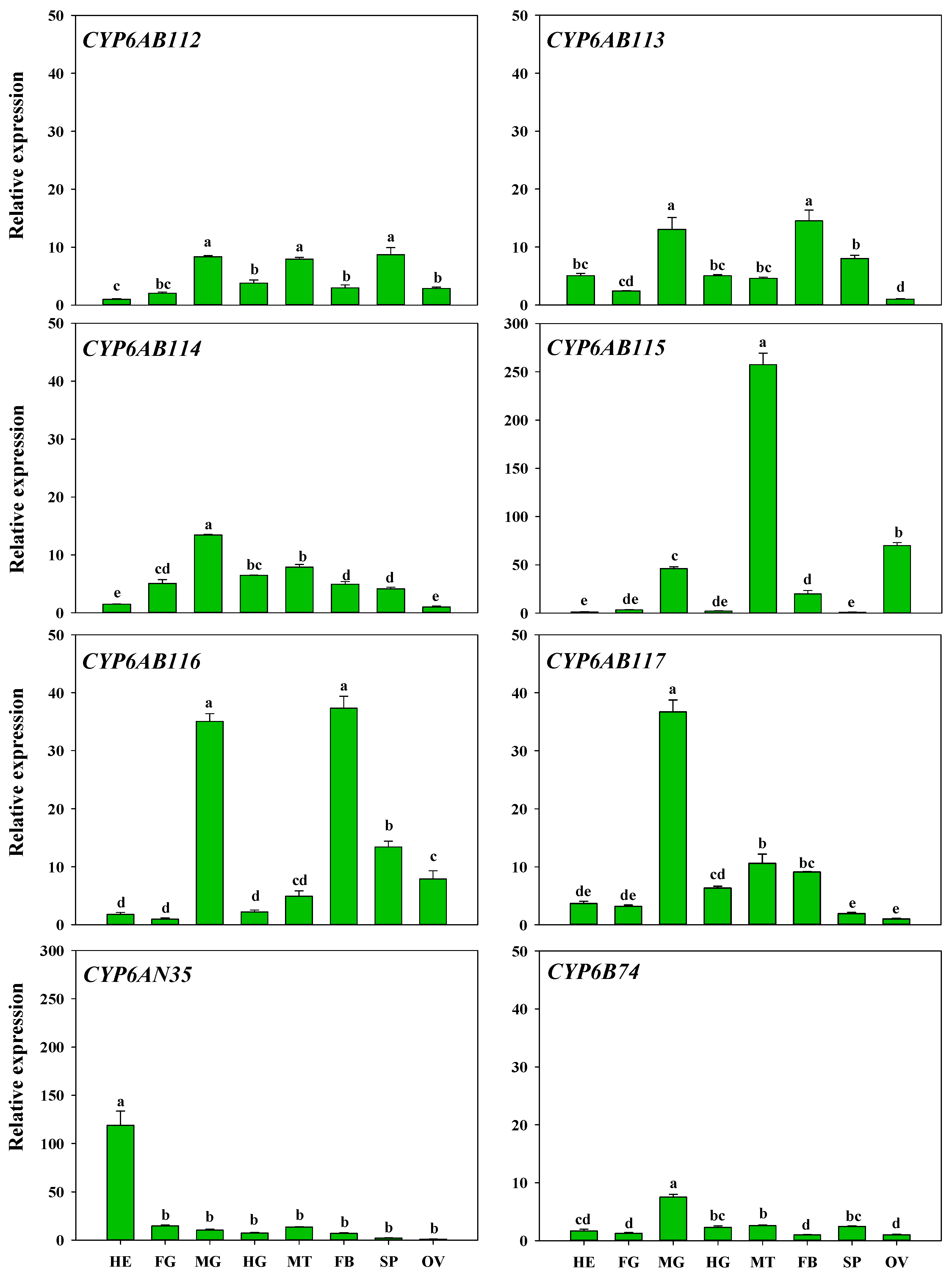
3.3. Developmental Stage and Tissue Specificity of P450 Gene Expression
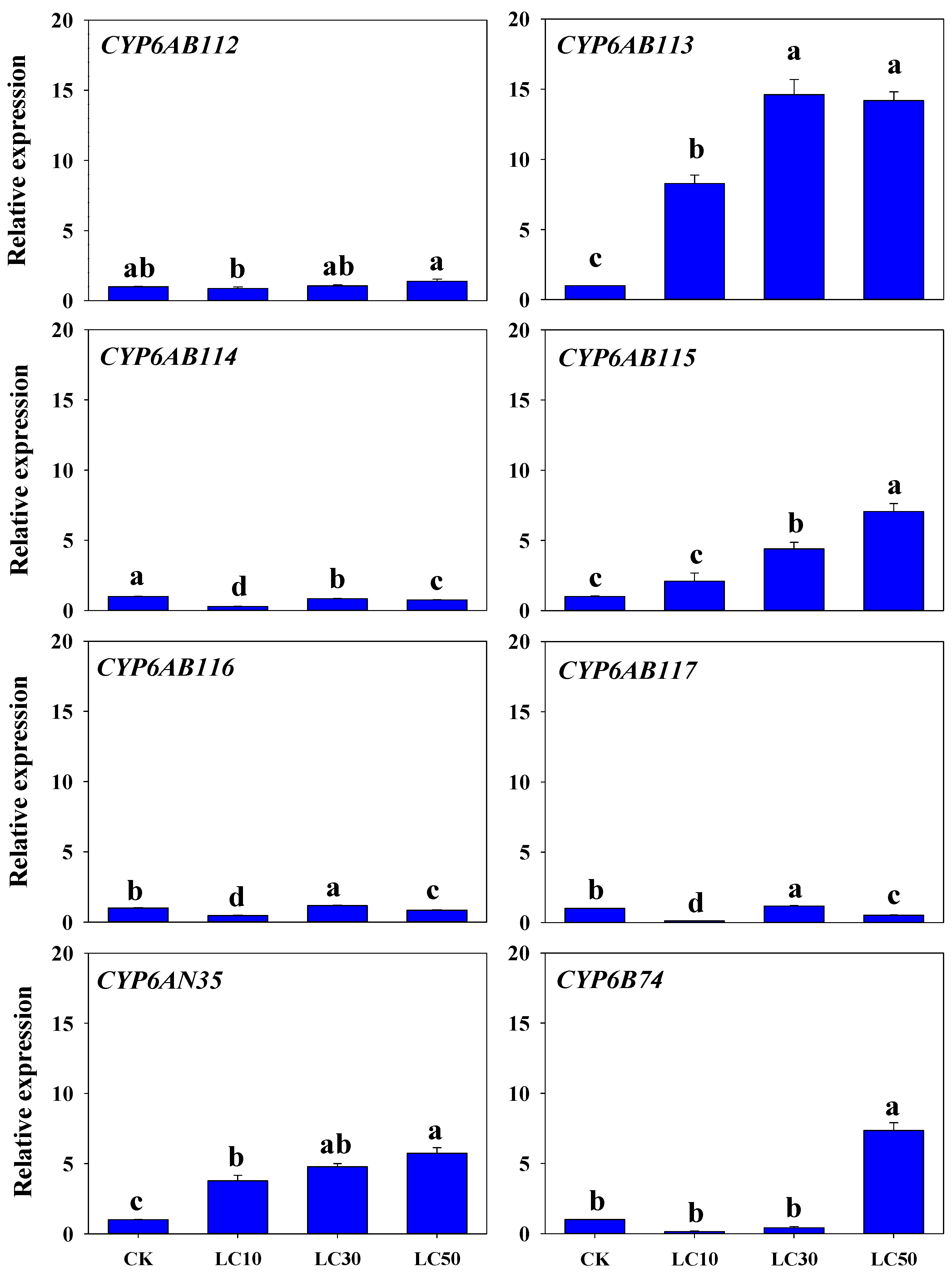
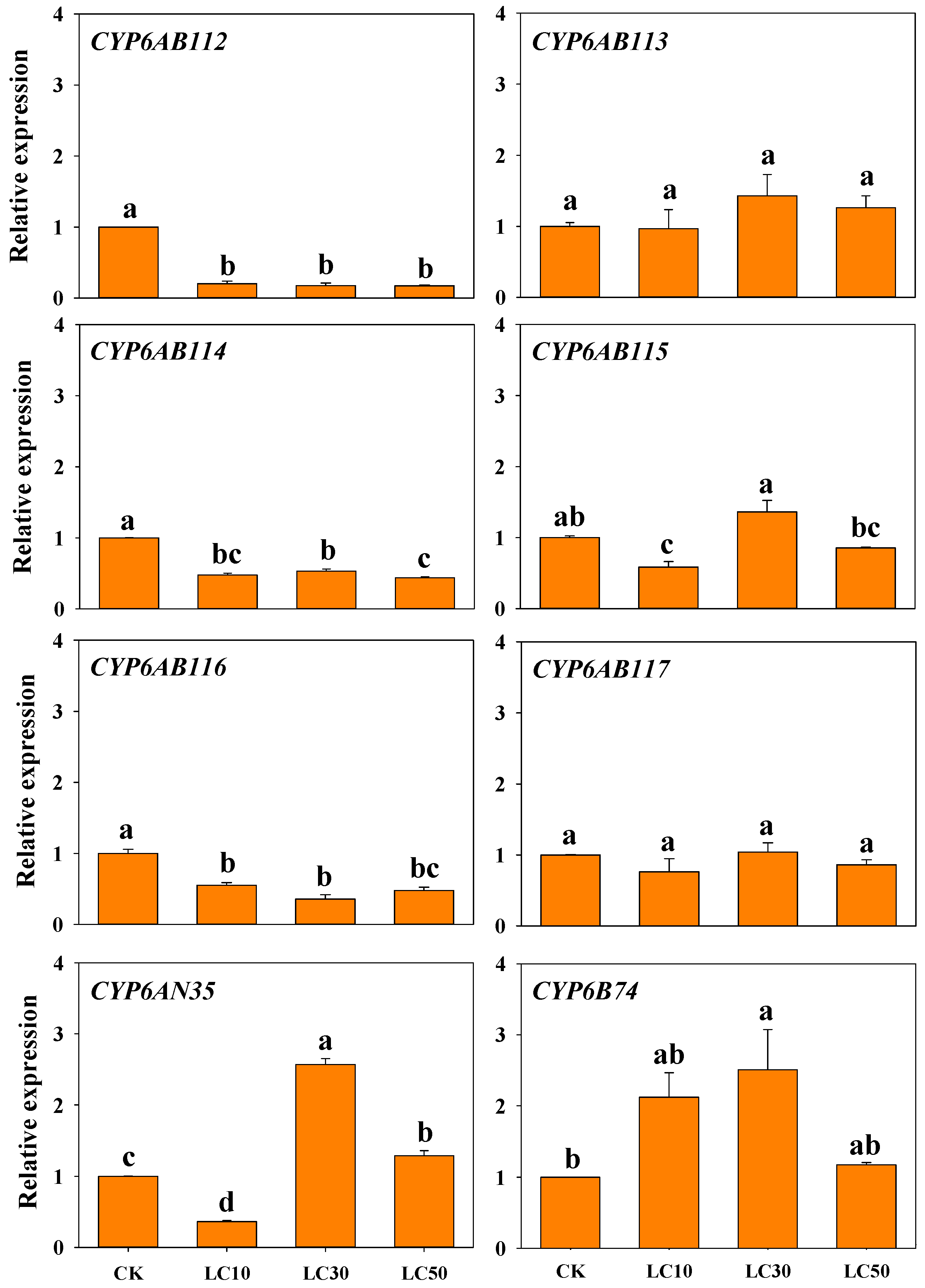
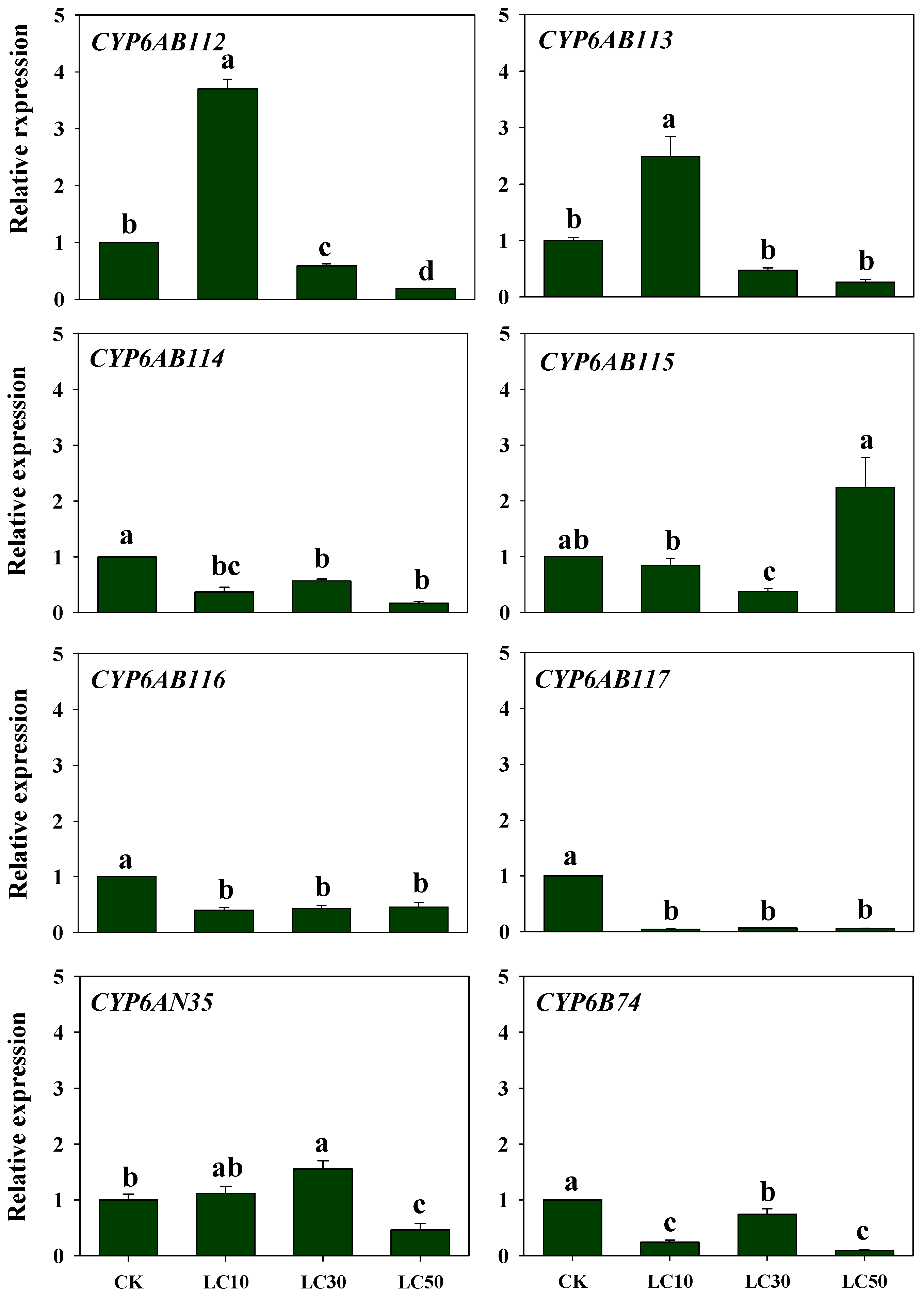
3.4. Response of CYP Genes Expression to Insecticide Exposure
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Liu, Y.Q.; Li, G.W. Zoology of China, Insecta, Volume 27, Lepidoptera, Torridae; Science Press: Beijing, China, 2002; p. 601. Available online: http://www.zoology.csdb.cn/taxon/%7B2AA1F11E-9939-4034-B926-29FBE1D192D9%7D (accessed on 25 January 2022).
- Myers, C.T.; Hull, L.A.; Krawczyk, G. Seasonal and cultivar-associated variation in oviposition preference of Oriental fruit moth (Lepidoptera: Tortricidae) adults and feeding behavior of neonate larvae in apples. J. Econ. Entomol. 2006, 99, 349–358. [Google Scholar] [CrossRef]
- Neven, L.G.; Kumar, S.; Yee, W.L.; Wakie, T. Current and future potential risk of establishment of Grapholita molesta (Lepidoptera: Tortricidae) in Washington State. Environ. Entomol. 2018, 47, 448–456. [Google Scholar] [CrossRef]
- Tian, Z.; Li, Y.; Xing, Y.; Li, R.; Liu, J. Structural insights into two representative conformations of the complex formed by Grapholita molesta (Busck) pheromone binding protein 2 and Z-8-dodecenyl acetate. J. Agric. Food Chem. 2019, 67, 4425–4434. [Google Scholar] [CrossRef]
- Amat, C.; Bosch-Serra, D.; Avilla, J.; Colomar, L.E. Different population phenologies of Grapholita molesta (Busck) in two hosts and two nearby regions in the NE of Spain. Insects 2021, 12, 612. [Google Scholar] [CrossRef] [PubMed]
- de Lame, F.M.; Hong, J.J.; Shearer, P.W.; Brattsten, L.B. Sex-related differences in the tolerance of Oriental fruit moth (Grapholita molesta) to organophosphate insecticides. Pest Manag. Sci. Former. Pestic. Sci. 2001, 57, 827–832. [Google Scholar] [CrossRef] [PubMed]
- Borchert, D.M.; Stinner, R.E.; Walgenbach, J.F.; Kennedy, G.G. Oriental fruit moth (Lepidoptera: Tortricidae) phenology and management with methoxyfenozide in North Carolina apples. J. Econ. Entomol. 2004, 97, 1353–1364. [Google Scholar] [CrossRef] [PubMed]
- Usmani, K.A.; Shearer, P.W. Susceptibility of male oriental fruit moth (Lepidoptera: Tortricidae) populations from New Jersey apple orchards to azinphosmethyl. J. Econ. Entomol. 2001, 94, 233–239. [Google Scholar] [CrossRef] [PubMed]
- Pree, D.J.; Whitty, K.J.; Van Driel, L. Resistance to insecticides in oriental fruit moth populations (Grapholita molesta) from the Niagara peninsula of Ontario. Can. Entomol. 1998, 130, 245–256. [Google Scholar] [CrossRef]
- Atanasso, A.; Shearer, P.W.; Hamilton, G.; Polk, D. Development and implementation of a reduced risk peach arthropod management program in New Jersey. J. Econ. Entomol. 2002, 95, 803–812. [Google Scholar] [CrossRef] [PubMed]
- Liu, N.N. Insecticide Resistance in Mosquitoes: Impact, Mechanisms, and Research Directions. Annu. Rev. Entomol. 2015, 60, 537–559. [Google Scholar] [CrossRef]
- Feyereisen, R. Insect cytochrome P450. In Comprehensive Molecular Insect Science; Gilbert, L.I., Iatrou, K., Gill, S.S., Eds.; Elsevier: Oxford, UK, 2005; Volume 4, pp. 1–77. [Google Scholar]
- Huang, Y.; Shen, G.M.; Jiang, H.B.; Jiang, X.Z.; Dou, W.; Wang, J.J. Multiple P450 genes: Identification, tissue-specific expression and their responses to insecticide treatments in the oriental fruit fly, Bactrocera dorsalis (Hendel) (Diptera: Tephritidea). Pestic. Biochem. Physiol. 2013, 106, 1–7. [Google Scholar] [CrossRef]
- Liska, D.J. The detoxification enzyme systems. Altern. Med. Rev. 1998, 3, 187–198. [Google Scholar] [PubMed]
- Adams, M.D.; Celniker, S.E.; Holt, R.A.; Evans, C.A.; Gocayne, J.D.; Amanatides, P.G.; Scherer, S.E.; Li, P.W.; Hoskins, R.A.; Galle, R.F.; et al. The genome sequence of Drosophila melanogaster. Science 2000, 287, 2185–2195. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Strode, C.; Wondj, C.S.; David, J.P.; Hawkes, N.J.; Lumjuan, N.; Nelson, D.R.; Drane, D.R.; Karunaratne, S.P.; Hemingway, J.; Black, W.C., 4th; et al. Genomic analysis of detoxification genes in the mosquito Aedes aegypti. Insect Biochem. Mol. Biol. 2008, 38, 113–123. [Google Scholar] [CrossRef] [PubMed]
- Xia, Q.; Zhou, Z.; Lu, C.; Cheng, D.; Dai, F.; Li, B.; Zhao, P.; Zha, X.; Cheng, T.; Chai, C.; et al. A draft sequence for the genome of the domesticated silkworm (Bombyx mori). Science 2004, 306, 1937–1940. [Google Scholar] [PubMed] [Green Version]
- Wan, F.; Yin, C.; Tang, R.; Chen, M.; Wu, Q.; Huang, C.; Qian, W.; Rota-Stabelli, O.; Yang, N.; Wang, S.; et al. A chromosome-level genome assembly of Cydia pomonella provides insights into chemical ecology and insecticide resistance. Nat. Commun. 2019, 10, 4237. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zhou, G.L.; Huang, J.L. Diversity and evolution of CYP6 family in insects. Chin. Bull. Entomol. 2002, 39, 246–251. [Google Scholar]
- Guo, Y.; Zhang, J.; Yang, M.; Yan, M.; Zhu, K.Y.; Guo, Y.; Ma, E. Comparative analysis of cytochrome P450-like genes from Locusta migratoria manilensis: Expression profiling and response to insecticide exposure. Insect Sci. 2012, 19, 75–85. [Google Scholar] [CrossRef]
- Bergé, J.; Feyereisen, R.; Amichot, M. Cytochrome P450 monooxygenases and insecticide resistance in insects. Philos. Trans. R. Soc. Lond. Ser. B Biol. Sci. 1998, 353, 1701–1705. [Google Scholar] [CrossRef] [Green Version]
- Zhang, X.Y.; Dong, J.; Wu, H.H.; Zhang, H.H.; Zhang, J.Z.; Ma, E.B. Knockdown of cytochrome P450 CYP6 family genes increases susceptibility to carbamates and pyrethroids in the migratory locust, Locusta migratoria. Chemosphere 2019, 223, 48–57. [Google Scholar] [CrossRef]
- Wang, H.D.; Shi, Y.; Wang, L.; Liu, S.; Wu, S.W.; Yang, Y.; Feyereisen, R.; Wu, Y. CYP6AE gene cluster knockout in Helicoverpa armigera reveals role in detoxification of phytochemicals and insecticides. Nat. Commun. 2018, 9, 4820. [Google Scholar] [CrossRef] [Green Version]
- Li, J.J.; Li, X.M.; Bai, R.; Shi, Y.; Tang, Q.B.; An, S.H.; Song, Q.H.; Yan, F.M. RNA interference of the P450 CYP6CM1 gene has different efficacy in B and Q biotypes of Bemisia tabaci. Pest Manag. Sci. 2015, 71, 1175–1181. [Google Scholar] [CrossRef]
- Wang, Y.; Tian, J.; Han, Q.; Zhang, Y.; Liu, Z. Genomic organization and expression pattern of cytochrome P450 genes in the wolf spider Pardosa pseudoannulata. Comp. Biochem. Physiol. Part C Toxicol. Pharmacol. 2021, 248, 109–118. [Google Scholar] [CrossRef]
- Li, L.; Gao, X.; Lan, M.; Yuan, Y.; Guo, Z.; Tang, P.; Li, M.; Liao, X.; Zhu, J.; Li, Z.; et al. De novo transcriptome analysis and identification of genes associated with immunity, detoxification and energy metabolism from the fat body of the tephritid gall fly, Procecidochares utilis. PLoS ONE 2019, 14, e0226039. [Google Scholar] [CrossRef]
- Guo, Y.Q.; Chai, Y.P.; Zhang, L.J.; Zhao, Z.G.; Gao, L.L.; Ma, R. Transcriptome analysis and identification of major detoxification gene families and insecticide targets in Grapholita molesta (Busck) (Lepidoptera: Tortricidae). J. Insect Sci. 2017, 17, 43. [Google Scholar] [CrossRef]
- Yu, Q.; Feng, Y.T.; Guo, X.J.; Du, E.Q.; Guo, G.M.; Zhang, R.X. Determination of the efficacy of the newly hatched larvae of Grapholitha molesta (Busck). Int. J. Fruit Sci. 2017, 34, 1483–1489. [Google Scholar]
- Du, J. Biological Characteristics and Artificial Rearing Techniques of Grapholitha molesta (Busck). Master’s Thesis, Northwest A&F University, Xianyang, China, 2009. [Google Scholar]
- Kumar, S.; Stecher, G.; Tamura, K. MEGA7: Molecular evolutionary genetics analysis version 7.0 for bigger datasets. Mol. Biol. Evol. 2016, 33, 1870–1874. [Google Scholar] [CrossRef] [Green Version]
- Han, H.; Mao, R.; Zhao, Z.; Zhang, L.; Gao, L.; Guo, Y. Indoor toxicity of three types of insecticides against Grapholita molesta Busck. J. Shanxi Agric. Sci. 2019, 47, 1470–1473. [Google Scholar]
- Li, S.; Su, L.; Li, B.; Li, Y.; Li, G.; Wu, J. Gene cloning, prokaryotic expression and enzymatic characterization of the glutathione S-transferase GmolGST6 of the pear carnivora. Acta Entomol. Sin. 2018, 61, 398–409. [Google Scholar]
- Livak, K.J.; Schmittgen, T.D. Analysis of relative gene expression data using real-time quantitative PCR and the 2(-Delta Delta C (T)) Method. Methods 2001, 25, 402–408. [Google Scholar] [CrossRef]
- Ai, J.; Zhu, Y.; Duan, J.; Yu, Q.; Zhang, G.; Wan, F.; Xiang, Z.H. Genome-wide analysis of cytochrome P450 monooxygenase genes in the silkworm, Bombyx mori. Gene 2011, 480, 42–50. [Google Scholar] [CrossRef] [PubMed]
- Feyereisen, R. Insect P450 enzymes. Annu. Rev. Entomol. 1999, 44, 507–533. [Google Scholar] [CrossRef] [PubMed]
- Li, X.; Schuler, M.A.; Berenbaum, M.R. Molecular mechanisms of metabolic resistance to synthetic and natural xenobiotics. Annu. Rev. Entomol. 2007, 52, 231–253. [Google Scholar] [CrossRef] [PubMed]
- Guo, Y.; Wu, H.; Zhang, X.; Ma, E.; Guo, Y.; Zhu, K.Y.; Zhang, J. RNA interference of cytochrome P450 CYP6F subfamily genes affects susceptibility to different insecticides in Locusta migratoria. Pest Manag. Sci. 2016, 72, 2154–2165. [Google Scholar] [CrossRef] [PubMed]
- Mittapelly, P.; Bansal, R.; Michel, A. Differential expression of cytochrome P450 CYP6 genes in the brown marmorated stink bug, Halyomorpha halys (Hemiptera: Pentatomidae). J. Econ. Entomol. 2019, 112, 1403–1410. [Google Scholar] [CrossRef] [PubMed]
- Pauchet, Y.; Wilkinson, P.; Vogel, H.; Nelson, D.R.; Reynolds, S.E.; Heckel, D.G.; French-Constant, R.H. Pyrosequencing the Manduca sexta larval midgut transcriptome: Messages for digestion, detoxification and defence. Insect Mol. Biol. 2010, 1, 61–75. [Google Scholar] [CrossRef] [PubMed]
- Li, H.S.; Dai, H.G.; Wang, J. Cloning and bioinformatics analysis of cytochrome P450 Gene from Plutella xylostella. Biotechnology 2005, 15, 8–11. [Google Scholar]
- Liu, Y.; Wang, N.; Zhang, Z.; Li, F. Transcriptomics analysis of drug resistance-related genes in five Lepidopteran pests. Chin. J. Appl. Entomol. 2012, 49, 317–323. [Google Scholar]
- Waterhouse, A.; Bertoni, M.; Bienert, S.; Studer, G.; Tauriello, G.; Gumienny, R.; Heer, F.T.; de Beer, T.A.P.; Rempfer, C.; Bordoli, L.; et al. SWISS-MODEL: Homology modelling of protein structures and complexes. Nucleic Acids Res. 2018, 46, W296–W303. [Google Scholar] [CrossRef] [Green Version]
- Su, C.; Liu, Y.; Li, S.; Yang, J.; Yen, Y. Mutations in the CYP1B1 gene may contribute to juvenile-onset open-angle glaucoma. Eye 2012, 26, 1369–1377. [Google Scholar] [CrossRef]
- Ziniel, P.D.; Karumudi, B.; Barnard, A.H.; Fisher, E.M.; Thatcher, G.R.; Podust, L.M.; Williams, D.L. The Schistosoma mansoni cytochrome P450 (CYP3050A1) is essential for worm survival and egg development. PLOS Negl. Trop. Dis. 2015, 9, e0004279. [Google Scholar] [CrossRef] [PubMed]
- Chen, C.; Shan, T.; Liu, Y.; Shi, X.; Gao, X. Identification of a novel cytochrome P450 CYP3356A1 linked with insecticide detoxification in Bradysia odoriphaga. Pest Manag. Sci. 2019, 75, 1006–1013. [Google Scholar] [CrossRef] [PubMed]
- Zimmer, C.T.; Bass, C.; Williamson, M.S.; Kaussmann, M.; Wölfel, K.; Gutbrod, O.; Nauen, R. Molecular and functional characterization of CYP6BQ23, a cytochrome P450 conferring resistance to pyrethroids in European populations of pollen beetle, Meligethes aeneus. Insect Biochem. Mol. Biol. 2014, 45, 18–29. [Google Scholar] [CrossRef] [PubMed]
- Xiong, W.; Gao, S.; Mao, J.; Wei, L.; Xie, J.; Liu, J.; Bi, J.; Song, X.; Li, B. CYP4BN6 and CYP6BQ11 mediate insecticide susceptibility and their expression is regulated by latrophilin in Tribolium castaneum. Pest Manag. Sci. 2019, 75, 2744–2755. [Google Scholar] [CrossRef] [PubMed]
- Pottier, M.A.; Bozzolan, F.; Chertemps, T.; Jacquin-Joly, E.; Lalouette, L.; Siaussat, D.; Maïbèche-Coisne, M. Cytochrome P450s and cytochrome P450 reductase in the olfactory organ of the cotton leafworm Spodoptera littoralis. Insect Mol. Biol. 2012, 21, 568–580. [Google Scholar] [CrossRef] [PubMed]
- Liu, J.; Wu, H.; Zhang, X.; Ma, W.; Zhu, W.; Silver, K.; Ma, E.; Zhang, J.; Zhu, K.Y. Metabolism of selected model substrates and insecticides by recombinant CYP6FD encoded by its gene predominately expressed in the brain of Locusta migratoria. Pestic. Biochem. Physiol. 2019, 159, 154–162. [Google Scholar] [CrossRef] [PubMed]
- Cohen, M.B.; Schuler, M.A.; Berenbaum, M.R. A host-inducible cytochrome P-450 from a host-specific caterpillar: Molecular cloning and evolution. Proc. Natl. Acad. Sci. USA 1992, 89, 10920–10924. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Snyder, M.J.; Stevens, J.L.; Andersen, J.F.; Feyereisen, R. Expression of cytochrome P450 genes of the CYP4 family in midgut and fat body of the tobacco hornworm, Manduca sexta. Arch. Biochem. Biophys. 1995, 321, 13–20. [Google Scholar] [CrossRef] [PubMed]
- Esquivel, C.J.; Cassone, B.J.; Piermarini, P.M. A de novo transcriptome of the Malpighian tubules in non-blood-fed and blood-fed Asian tiger mosquitoes Aedes albopictus: Insights into diuresis, detoxification, and blood meal processing. PeerJ 2016, 4, e1784. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zhou, X.; Fan, X.; Gao, Y.; Yang, J.; Qian, J.; Fan, D. Identification of two novel P450 genes and their responses to deltamethrin in the cabbage moth, Mamestra brassicae Linnaeus. Pestic. Biochem. Physiol. 2017, 141, 76–83. [Google Scholar] [CrossRef] [PubMed]
- Chung, H.; Sztal, T.; Pasricha, S.; Sridhar, M.; Batterham, P.; Daborn, P.J. Characterization of Drosophila melanogaster cytochrome P450 genes. Proc. Natl. Acad. Sci. USA 2009, 106, 5731–5736. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zhang, B.; Zhang, L.; Cui, R.; Zeng, X.; Gao, X. Cloning and expression of multiple cytochrome P450 genes: Induction by fipronil in workers of the red imported fire ant (Solenopsis invicta Buren). PLoS ONE 2016, 11, e0150915. [Google Scholar] [CrossRef] [PubMed]
- Sun, L.N.; Qiu, G.S.; Cui, L.; Ma, C.S.; Yuan, H.Z. Molecular characterization of a ryanodine receptor gene from Spodoptera exigua and its upregulation by chlorantraniliprole. Pestic. Biochem. Physiol. 2015, 123, 56–63. [Google Scholar] [CrossRef] [PubMed]
- Riddick, D.S.; Lee, C.; Bhathena, A.; Timsit, Y.E.; Cheng, P.Y.; Morgan, E.T.; Prough, R.A.; Ripp, S.L.; Miller, K.K.; Jahan, A.; et al. Transcriptional suppression of cytochrome P450 genes by endogenous and exogenous chemicals. Drug Metab. Dispos. 2004, 32, 367–375. [Google Scholar] [CrossRef]
- Li, X.; Baudry, J.; Berenbaum, M.R.; Schuler, M.A. Structural and functional divergence of insect CYP6B proteins: From specialist to generalist cytochrome P450. Proc. Natl. Acad. Sci. USA 2004, 101, 2939–2944. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Marcombe, S.; Fustec, B.; Cattel, J.; Chonephetsarath, S.; Thammavong, P.; Phommavanh, N.; David, J.P.; Corbel, V.; Sutherland, I.W.; Hertz, J.C.; et al. Distribution of insecticide resistance and mechanisms involved in the arbovirus vector Aedes aegypti in Laos and implication for vector control. PLoS Negl. Trop. Dis. 2019, 13, e0007852. [Google Scholar] [CrossRef] [Green Version]
- Ffrench-Constant, R.H. The molecular genetics of insecticide resistance. Genetics 2013, 194, 807–815. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Guo, Y.; Zhang, X.; Wu, H.; Yu, R.; Zhang, J.; Zhu, K.Y.; Guo, Y.; Ma, E. Identification and functional analysis of a cytochrome P450 gene CYP9AQ2 involved in deltamethrin detoxification from Locusta migratoria. Pestic. Biochem. Physiol. 2015, 122, 1–7. [Google Scholar] [CrossRef] [PubMed]
- Terriere, L.C. Induction of detoxication enzymes in insects. Annu. Rev. Entomol. 1984, 29, 71–88. [Google Scholar] [CrossRef] [PubMed]
- Lu, K.; Song, Y.; Zeng, R. The role of cytochrome P450-mediated detoxification in insect adaptation to xenobiotics. Curr. Opin. Insect Sci. 2021, 43, 103–107. [Google Scholar] [CrossRef] [PubMed]







Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Han, H.; Yang, Y.; Hu, J.; Wang, Y.; Zhao, Z.; Ma, R.; Gao, L.; Guo, Y. Identification and Characterization of CYP6 Family Genes from the Oriental Fruit Moth (Grapholita molesta) and Their Responses to Insecticides. Insects 2022, 13, 300. https://doi.org/10.3390/insects13030300
Han H, Yang Y, Hu J, Wang Y, Zhao Z, Ma R, Gao L, Guo Y. Identification and Characterization of CYP6 Family Genes from the Oriental Fruit Moth (Grapholita molesta) and Their Responses to Insecticides. Insects. 2022; 13(3):300. https://doi.org/10.3390/insects13030300
Chicago/Turabian StyleHan, Hui, Yanyu Yang, Jun Hu, Yuanxin Wang, Zhiguo Zhao, Ruiyan Ma, Lingling Gao, and Yanqiong Guo. 2022. "Identification and Characterization of CYP6 Family Genes from the Oriental Fruit Moth (Grapholita molesta) and Their Responses to Insecticides" Insects 13, no. 3: 300. https://doi.org/10.3390/insects13030300