Larval Morphological Adaptations of Leiodes cinnamomea (Panzer, 1793) (Coleoptera: Leiodidae: Leiodinae)—Obligatory Feeder of Tuber Species


Abstract
:Simple Summary
Abstract
1. Introduction
2. Materials and Methods
2.1. Materials
2.2. Breeding
2.3. Preparation
2.4. Light Photography and Image Processing
2.5. SEM Preparation
2.6. Terminology and Measurements
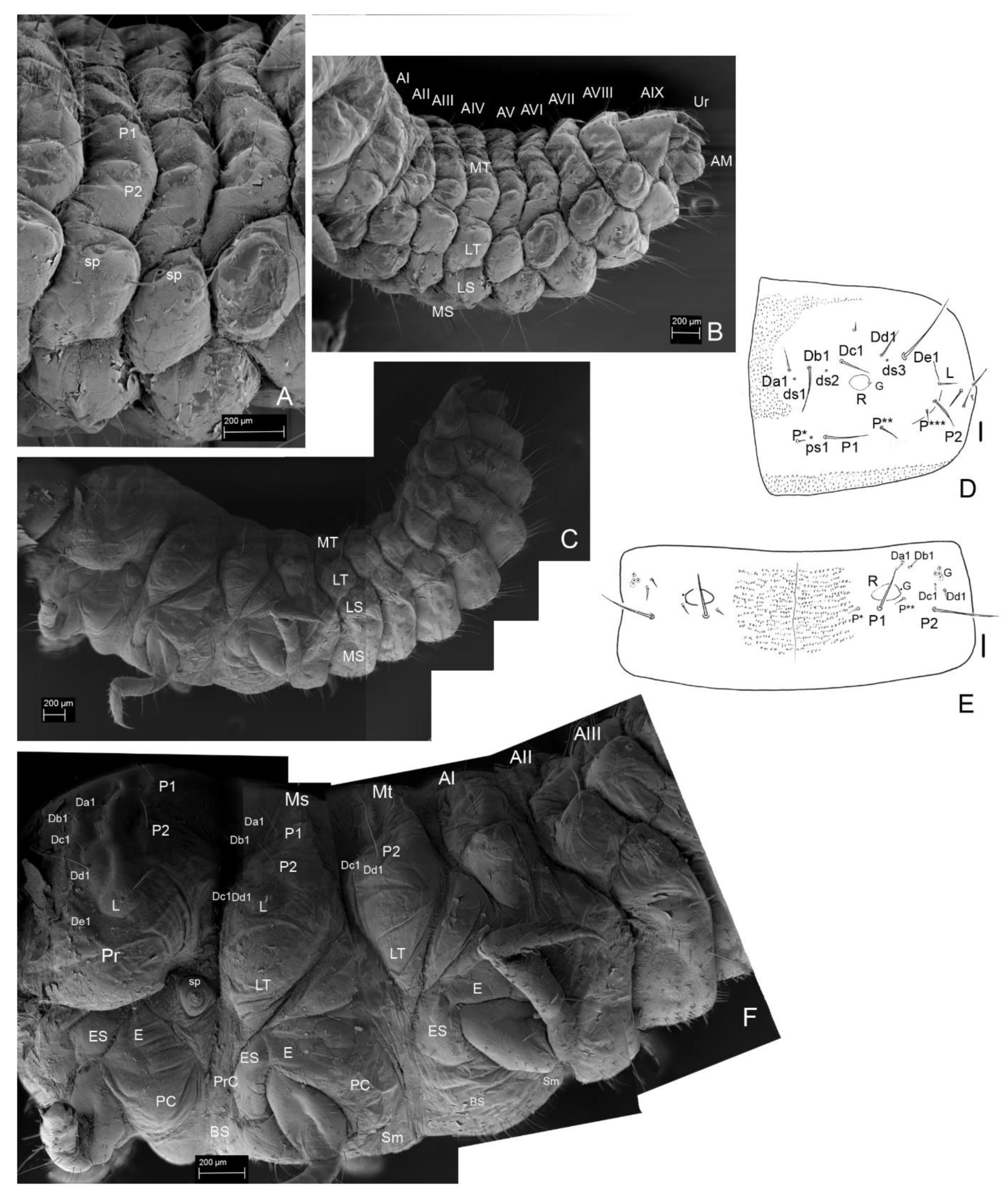
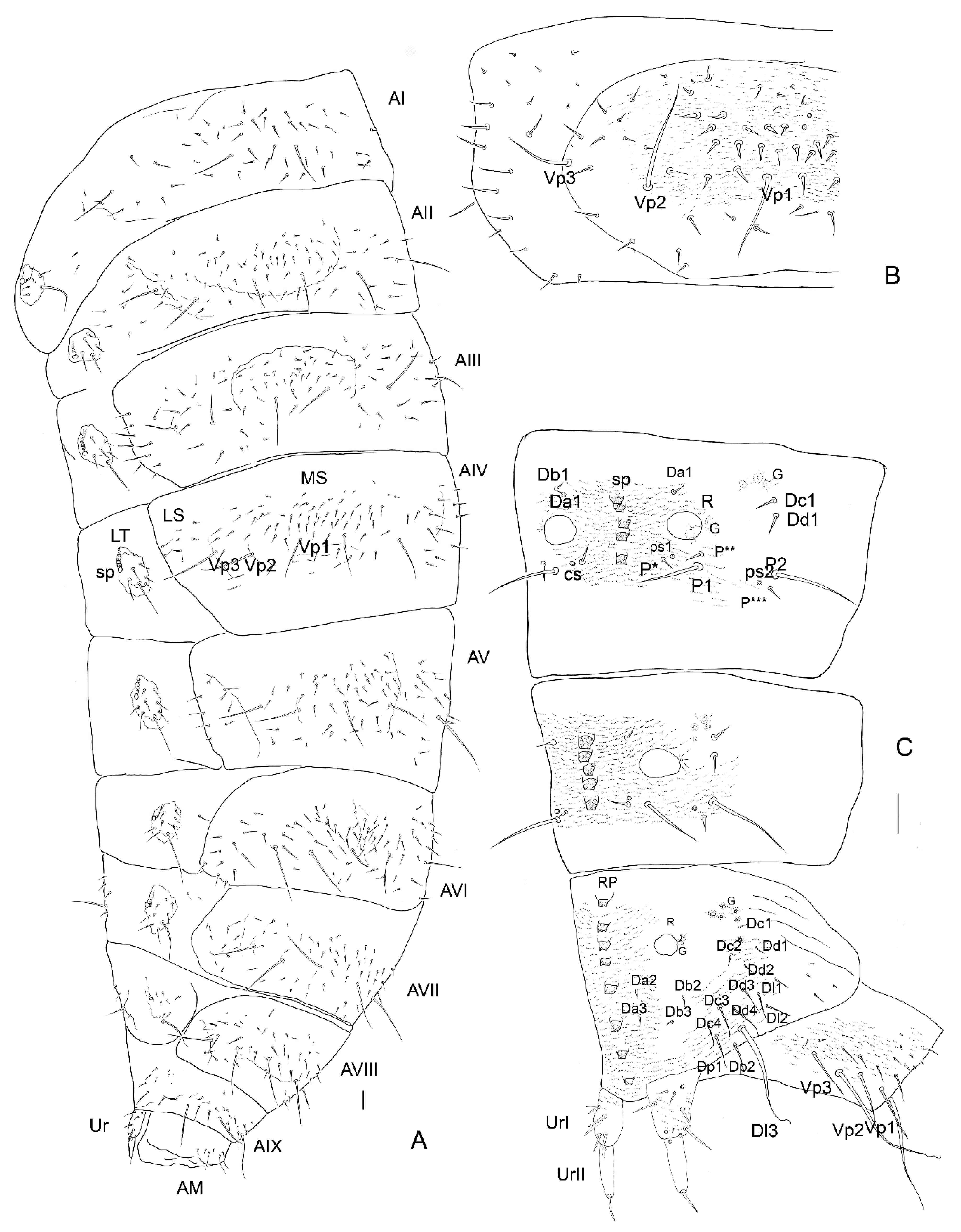
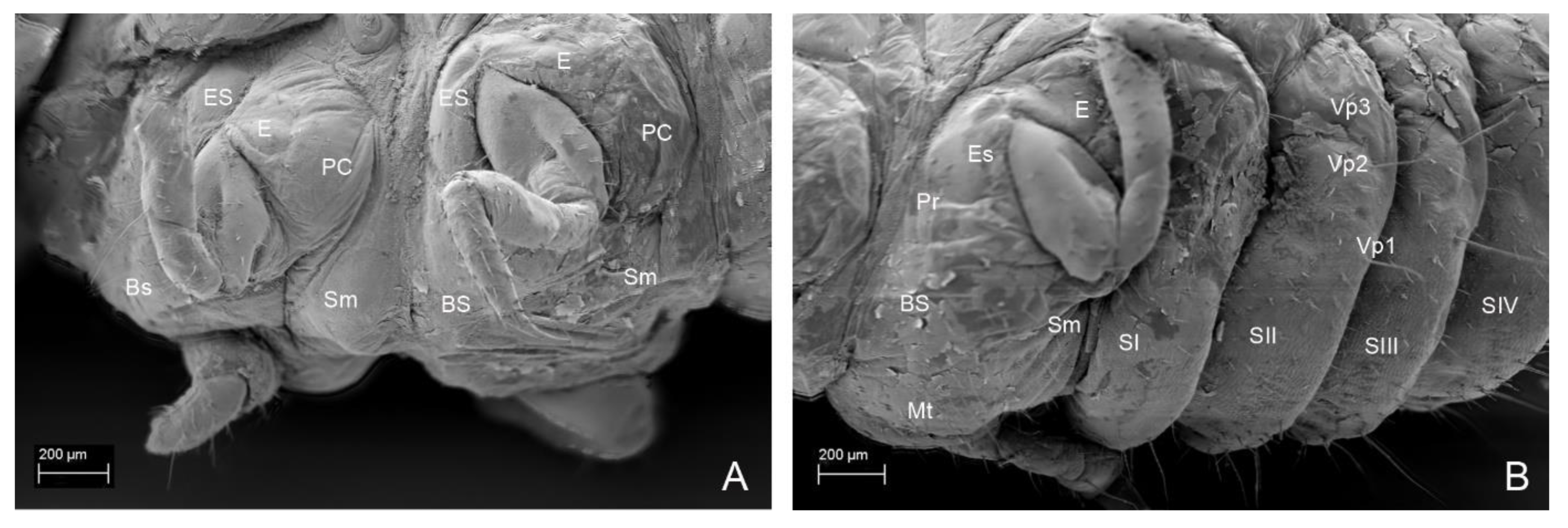
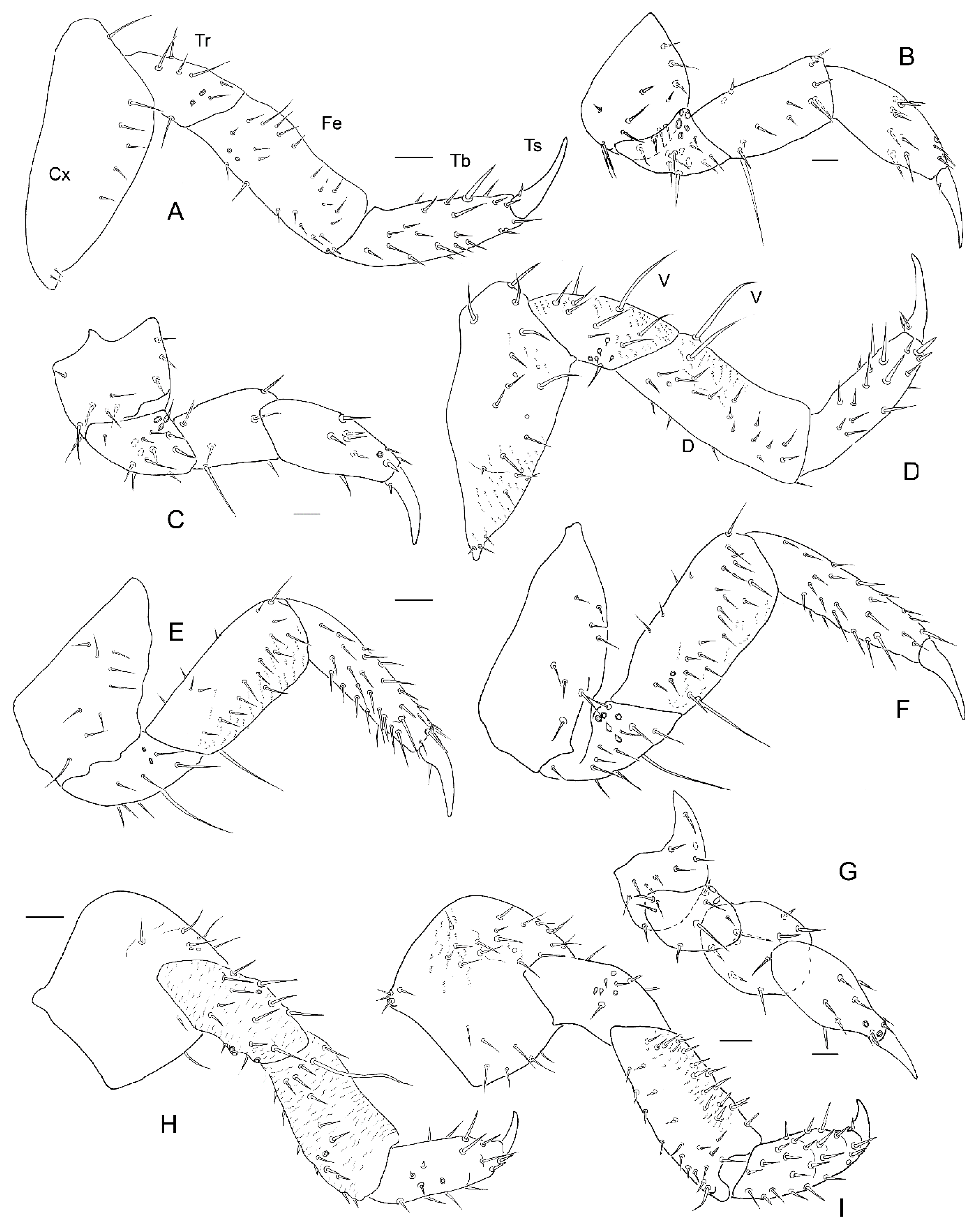
3. Results
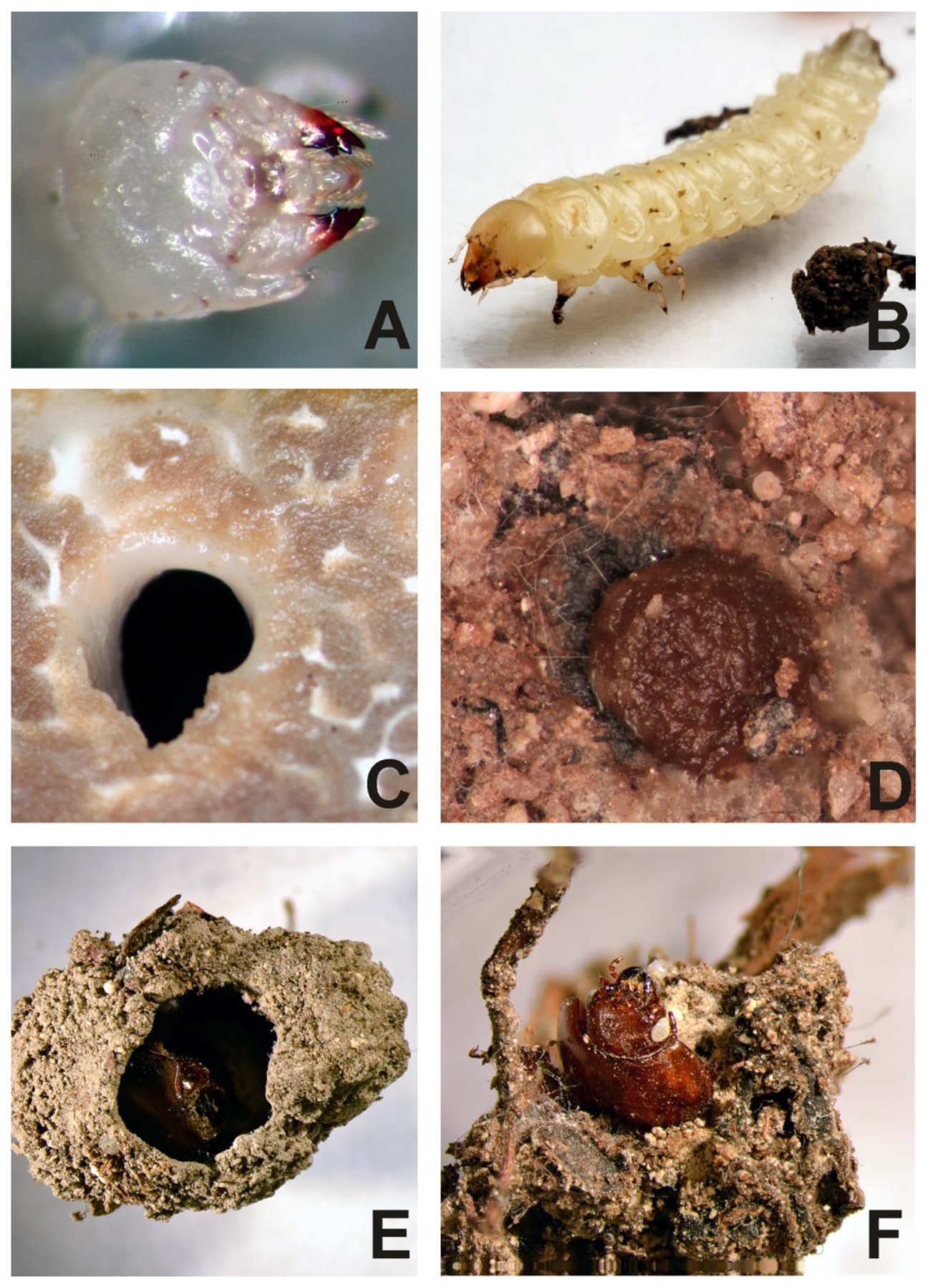
3.1. Biology



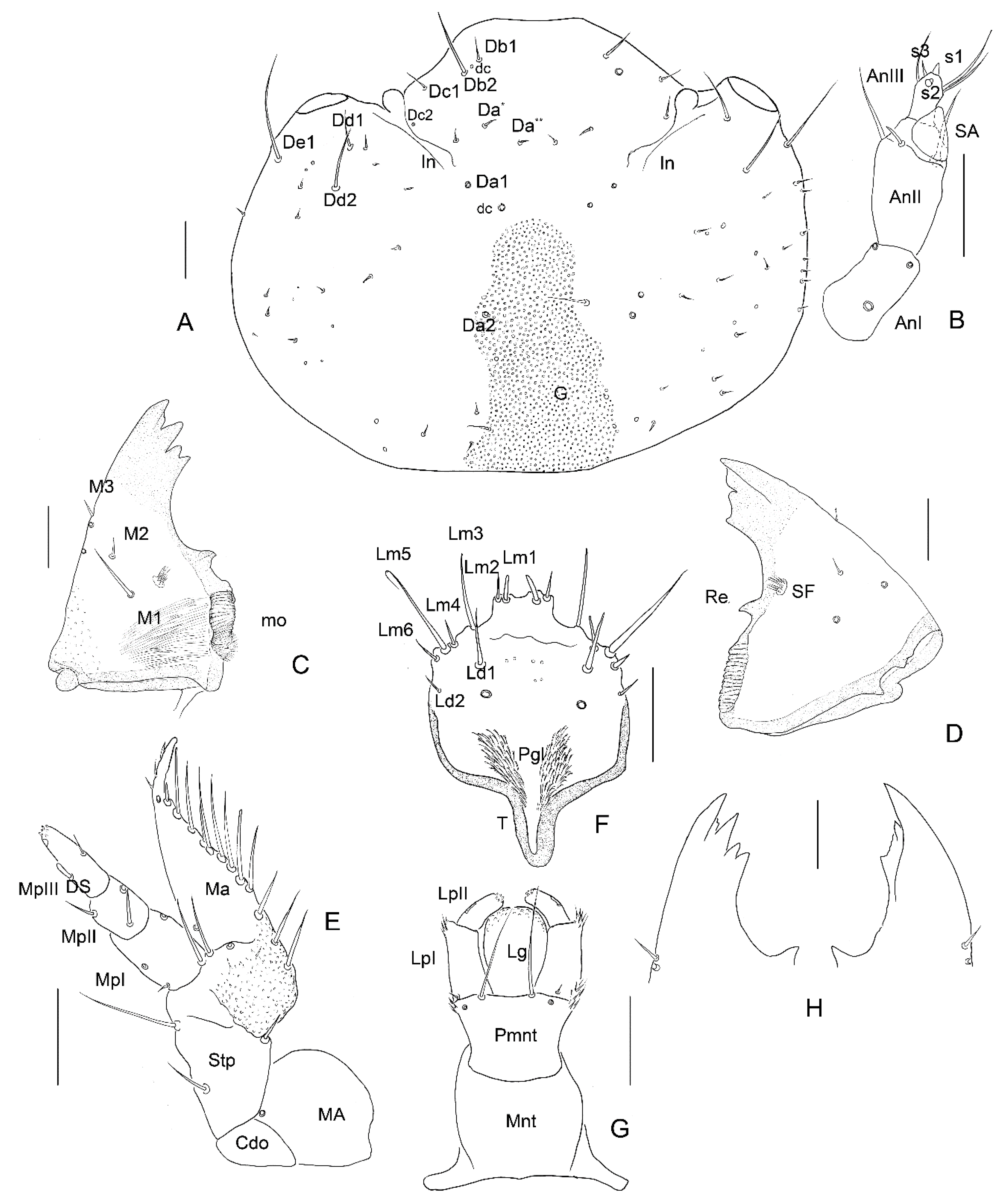
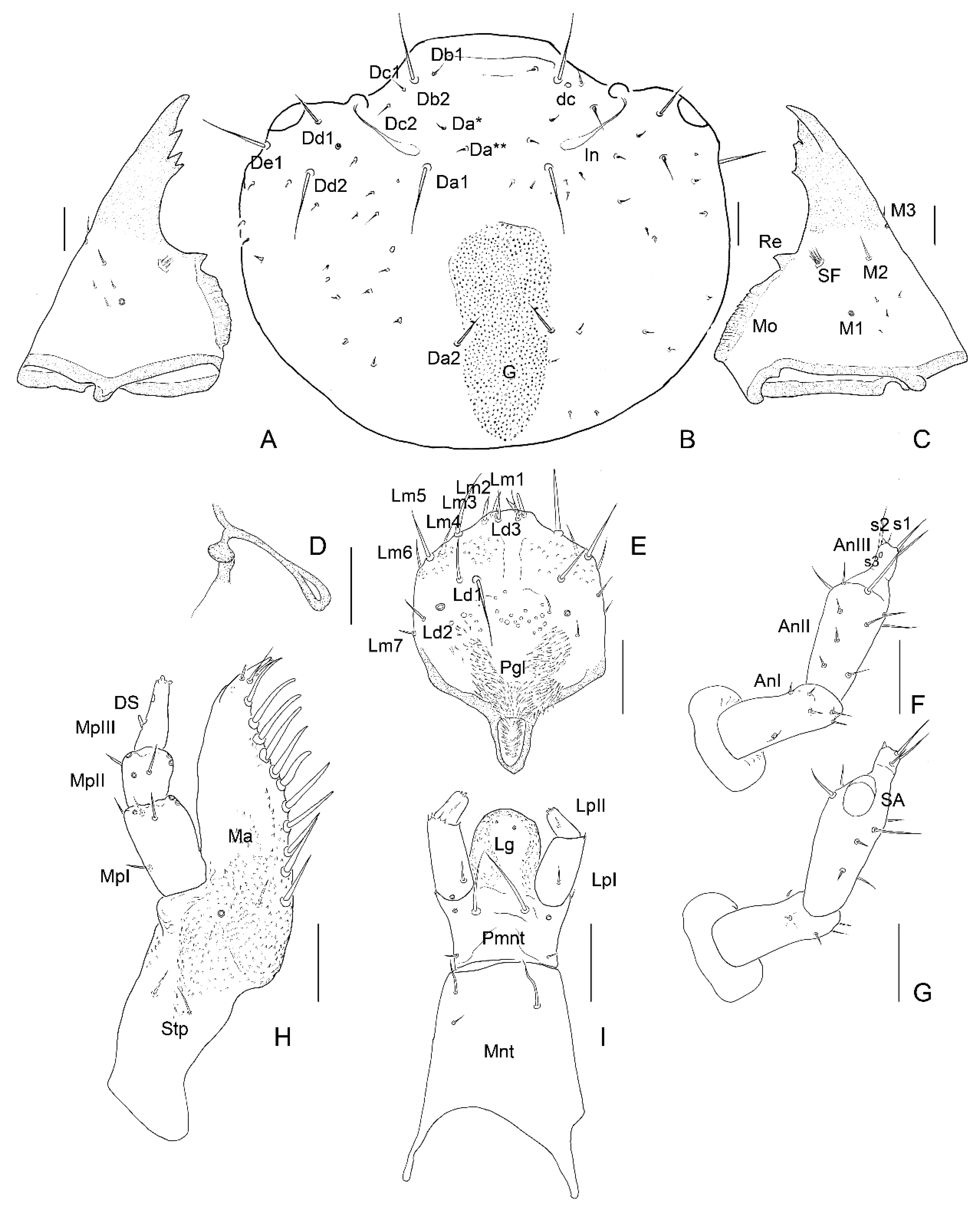
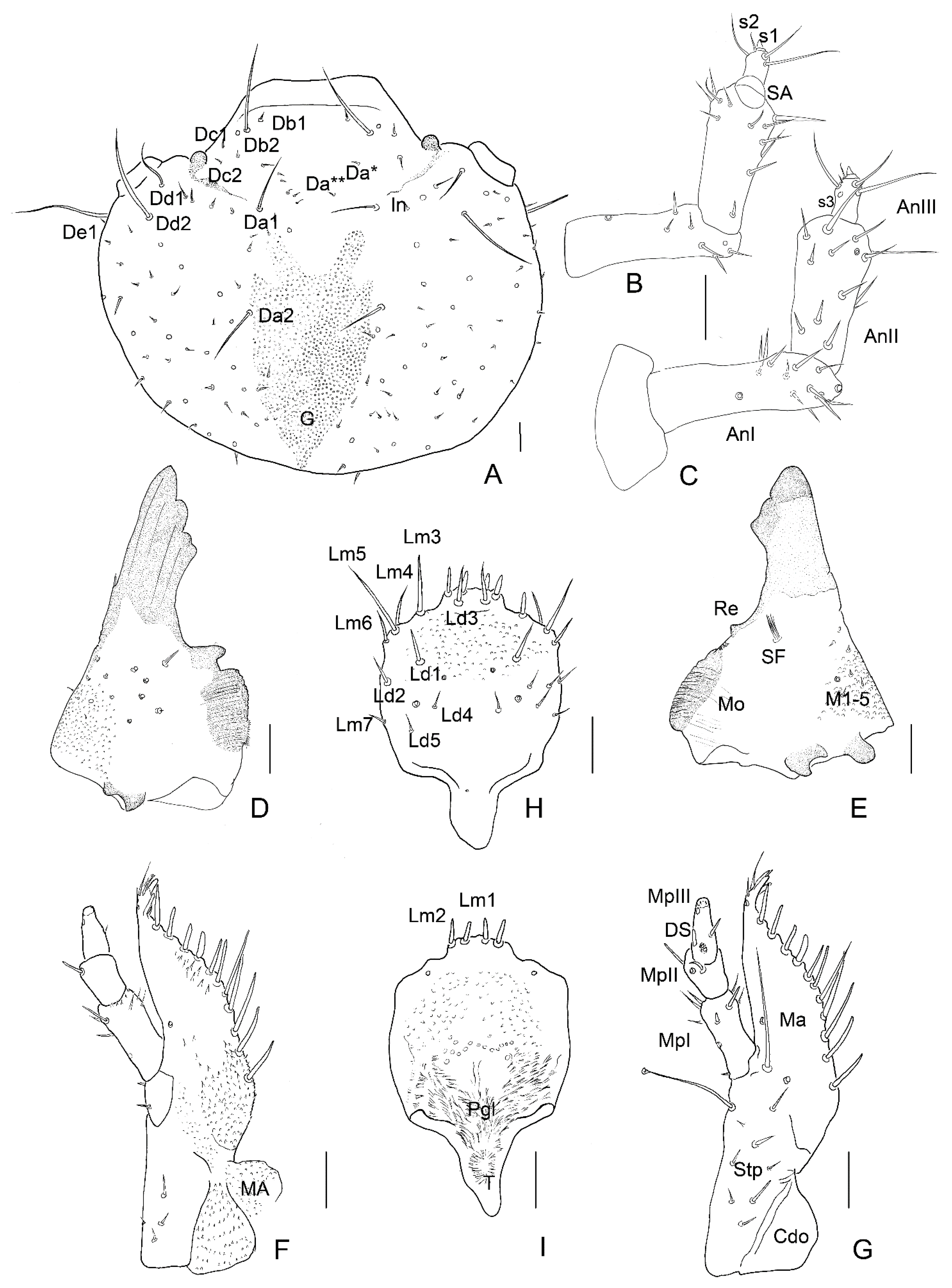
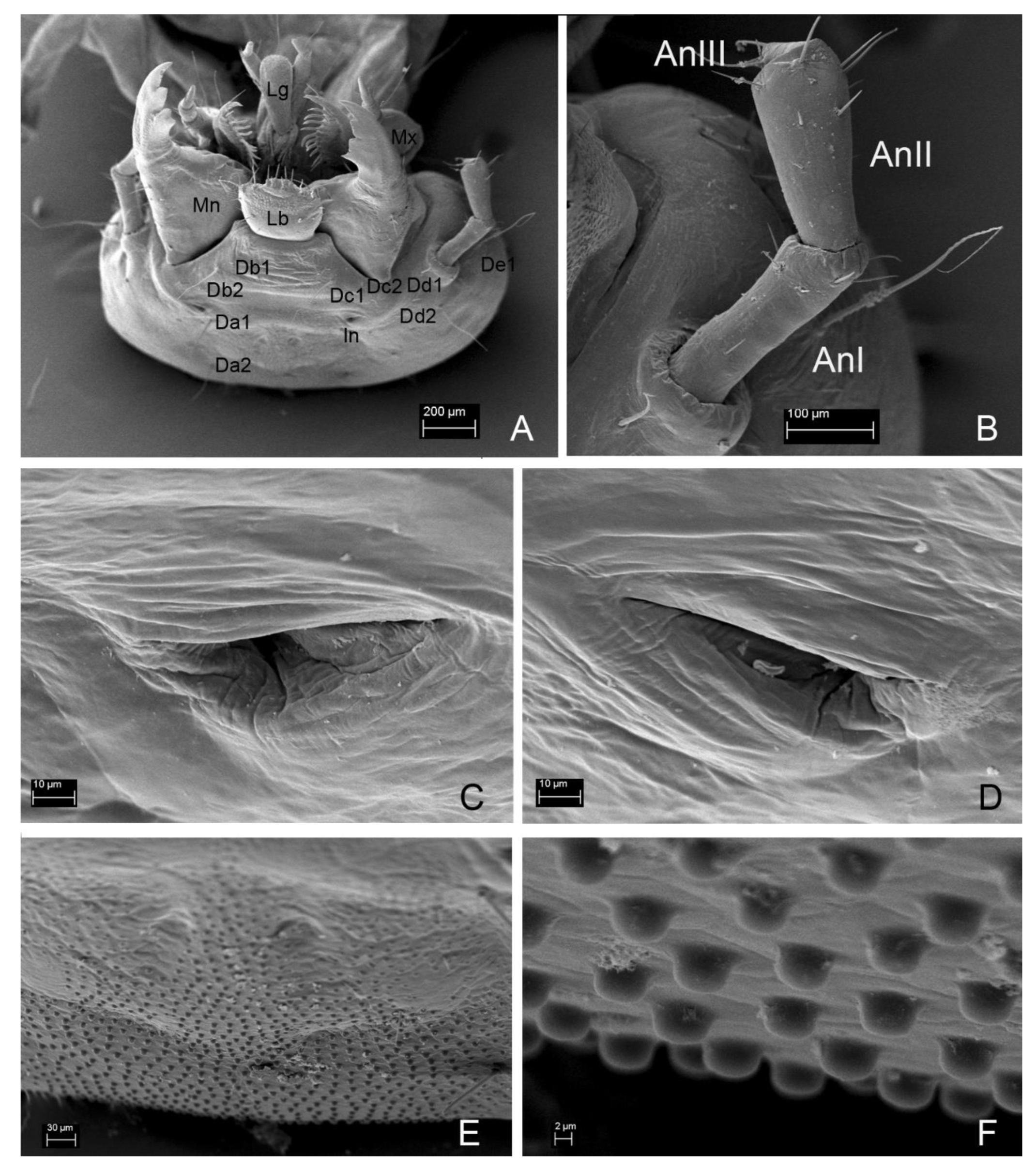
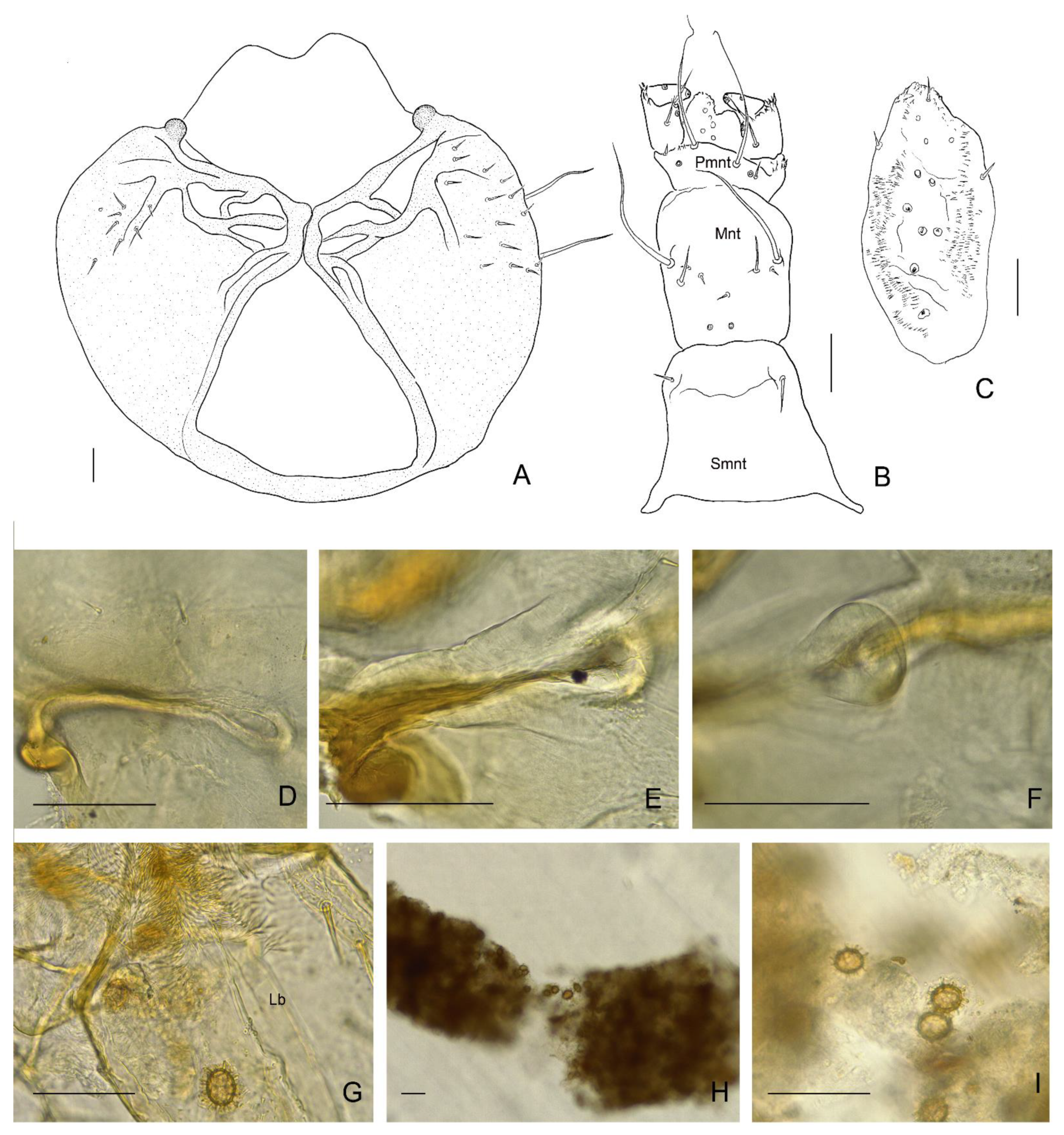
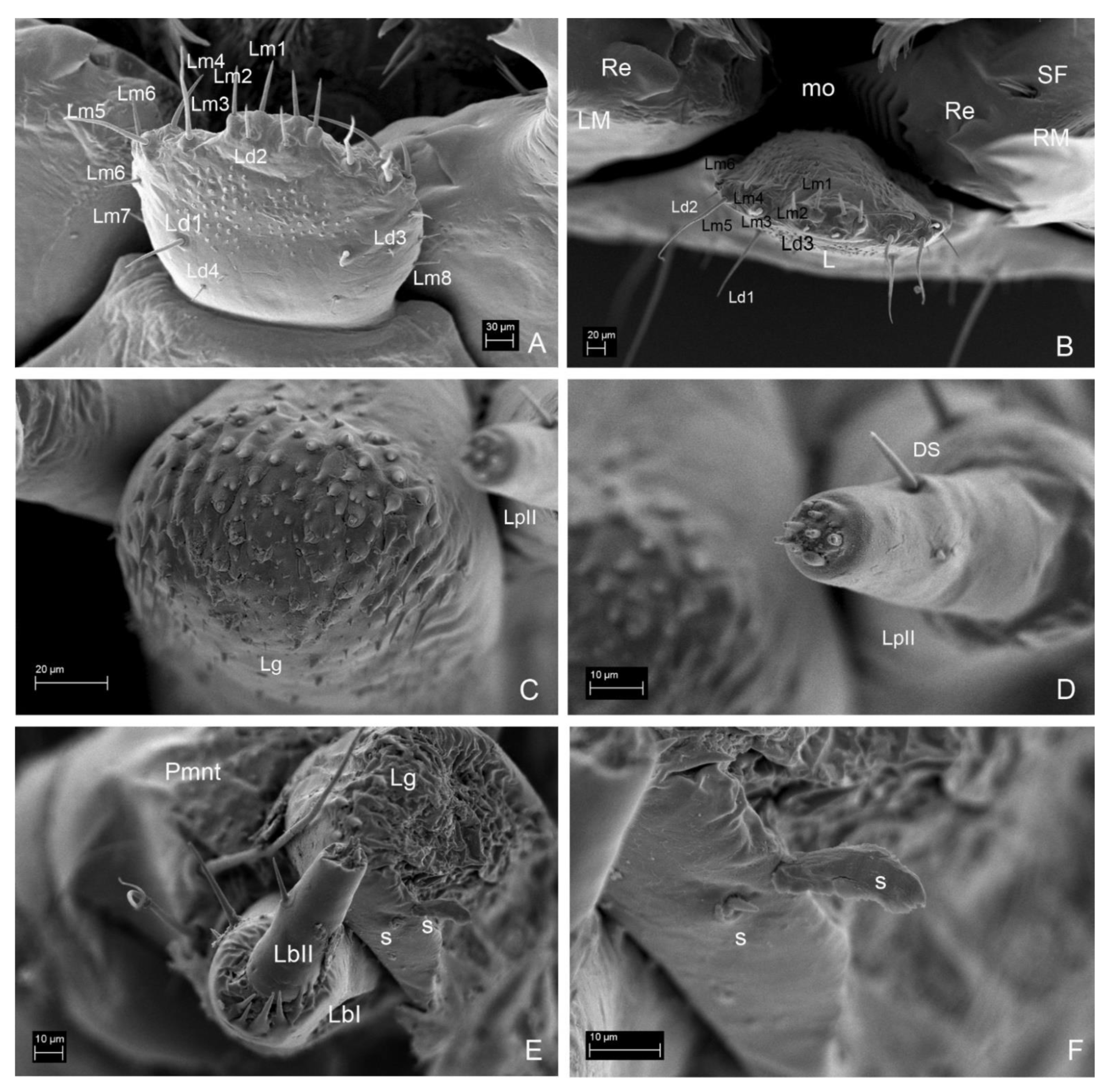
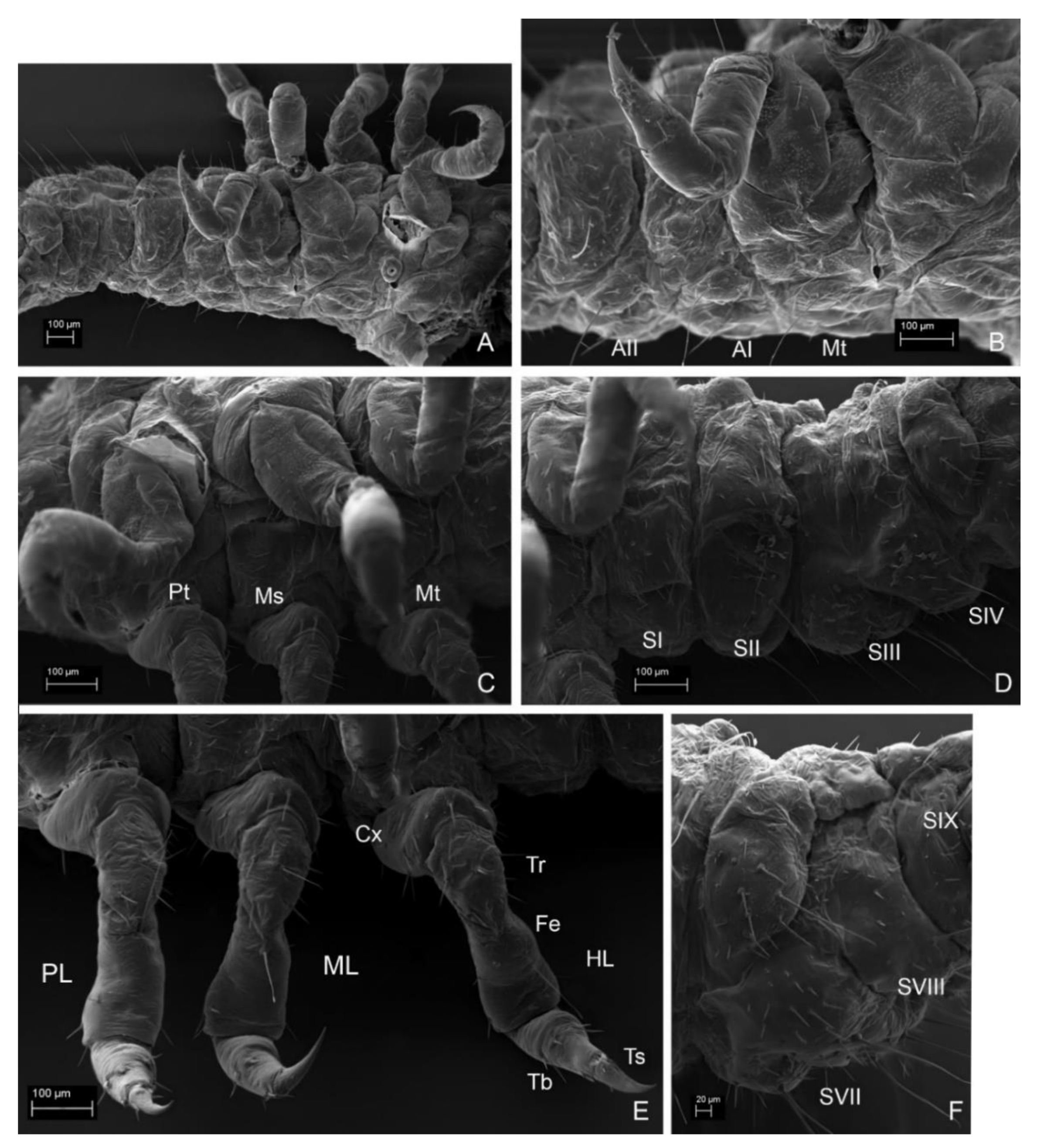
3.2. Description of the Immature Stages of Leiodes cinnamomea
















4. Discussion
5. Conclusions
- Soft-bodied, grub-like larva, with large head and mandibles, lack of stemmata, shortened legs, very small 2-segmented urogomphi and arching behaviour—adaptation to subterranean mycophagy.
- Three unique structures among leiodid larvae: exoskeletal invaginations (pseudomycangia) and granulation on head, median row of sclerotized plates on tergites VII–IX.
- Granulation and sclerotized plates seem to be correlated with arching up behaviour as organ of stridulation, although the act was not observed.
- Lack of characters of “leptinid association” of other leiodid larvae [61]: epicranial lines of head, mandibular molae with denticles, fimbriate galea.
- Morphology of larvae, mostly head, mouthparts, tergites and ratios of antennae and urogomphi, similar to those in Agathidiini and Scotocryptini, roughly confirmed the phylogenetic relationship in the family because we do not know larval morphology of other tribes of Leiodinae.
- Small differences of chaetotaxy among instars but tendency toward grub-like form of body with tergites and sternites divided into folds.
- Mosaic strategy—for beetles living in ephemeral and in solid fungi.
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
Abbreviations
| AI | Length of Antennomere I |
| AII | Length of Antennomere II |
| AIII | Length of Antennomere III |
| A1L | Length of Abdominal Segment I |
| A1L/W | Ratio of Abdominal Segment I Length: Abdominal Segment Width |
| A1W | Width of Abdominal Segment I |
| SA | Length of Sensory Appendage |
| HL | Head Length (without labrum) |
| HW | Head Width (at the widest point) |
| HW/HL | Ratio of Head Width: Head Length |
| LPI | Length of Labial Palp Segment I |
| LPII | Length of Labial Palp Segment II |
| MPI | Length of Maxillary Palp Segment I |
| MPII | Length of Maxillary Palp Segment II |
| MPIII | Length of Maxillary Palp Segment III |
| N1L | Pronotal Length |
| N1L/W | Ratio of Pronotal Length: Pronotal Width |
| N1W | Pronotal Width (at the widest point) |
| N2L | Mesonotal Length |
| N2L/W | Ratio of Mesonotal Length: Mesonotal Width |
| N2W | Mesonotal Width (at the widest point) |
| N3L | Metanotal Length |
| N3L/W | Ratio of Metanotal Length: Metanotal Width |
| N3W | Metanotal Width (at the widest point) |
| URI | Length of Urogomphal Segment I |
| URII | Length of Urogomphal Segment II |
| URS | Length of Urogomphal Terminal Seta |
References
- Newton, A.F. Phylogenetic problems, current classification and generic catalogue of world Leiodidae (including Cholevidae). In Phylogeny and Evolution of Subterranean and Endogean Cholevidae (=Leiodidae Cholevinae); Giachino, P.M., Peck, S.B., Eds.; Proceedings of XXI.C.E., Firenze, 1996; Atti Museo Regionale di Scienze Naturali: Torino, Italy, 1998; pp. 41–178. Available online: https://ci.nii.ac.jp/naid/10018719070/ (accessed on 2 February 2022).
- Beutel, R.G.; Leschen, R.A.B. Handbuch der Zoologie, Volume 4. Arthropoda: Insecta. Coleoptera, Beetles. Volume 1. Part 38. Morphology and Systematics (Archostemata, Adephaga, Myxophaga, Polyphaga partim); De Gruyt: Berlin, Germany, 2005; pp. 269–280. [Google Scholar]
- Park, S.-J.; Ahn, K.J. The Korean Species of the Genus Leiodes Latreille with descriptions of five new species (Coleoptera: Leiodidae: Leiodinae). J. Kans. Entomol. Soc. 2007, 80, 27–42. [Google Scholar] [CrossRef]
- Burakowski, B.; Mroczkowski, M.; Stefańska, J. Chrząszcze—Coleoptera. Histeroidea i Staphylinoidea prócz Staphylinidae. In Katalog Fauny Polski, XXIII, 5; PWN: Warszawa, Poland, 1978; 356p, Available online: https://rcin.org.pl/publication/36400 (accessed on 2 February 2022).
- Daffner, H. Revision der paläarktischen Arten der Tribus Leiodini Leach (Coleoptera, Leiodidae). Folia Ent. Hung. 1983, 44, 9–163. Available online: https://agris.fao.org/agris-search/search.do?recordID=US201302147576 (accessed on 2 February 2022).
- Pedersen, J. The truffle-beetle Leiodes cinnamomea (Panzer, 1793) found in Denmark (Coleoptera, Leiodidae). Entom. Meddel. 1994, 62, 101–104. Available online: https://www.cabi.org/ISC/abstract/19951114122 (accessed on 2 February 2022).
- Perreau, M. Leiodidae. In Catalogue of Palaearctic Coleoptera, Volume 2: Hydrophiloidea-Staphylinoidea; Löbl, I., Löbl, D., Eds.; Brill: Leiden, Poland; Boston, UK, 2015; xxvi + 1702p; ISBN1 978-90-04-28992-5. ISBN2 978-90-04-29685-5. Available online: https://www.zin.ru/animalia/coleoptera/pdf/palcatalogue_2nded_introduction.pdf (accessed on 2 February 2022).
- Angelini, F.; Švec, Z. Review of Chinese species of the subfamily Leiodinae (Coleoptera, Leiodidae). Acta Soc. Zool. Bohem. 1994, 58, 1–31. Available online: https://www.academia.edu/35630636/Angelini_F._and_Z._Svec_1994_-_Review_of_Chinese_species_of_the_subfamily_Leiodinae.pdf (accessed on 2 February 2022).
- Angelini, F.; Švec, Z. New species and records of Leiodinae from China (Coleoptera: Leiodidae). Linzer biol. Beitr. 1995, 27/2, 507–523. Available online: https://www.zobodat.at/pdf/LBB_0027_2_0507-0523.pdf (accessed on 2 February 2022).
- Švec, Z. Leiodes nikodymi n. sp from China with review of Chinese species of the genus Leiodes Latr. (Coleoptera, Leiodidae). Acta Entoml. Bohemoslov. 1991, 88, 377–380. [Google Scholar]
- Švec, Z. Chinese species of the genus Leiodes (Coleoptera: Leiodidae: Leiodinae). Acta Soc. Zool. Bohem. 2000, 64, 97–113. [Google Scholar]
- Cooter, J.; Kilian, A. New species of Leiodes Latreille (Col, Leiodidae) from China. Entomol.’s Mon. Mag. 2002, 138, 157–164. [Google Scholar]
- Baranowski, R. Revision of the genus Leiodes Latreille of North and Central America (Coleoptera: Leiodidae). Scand. Entomol. Suppl. 1993, 42, 1–149. [Google Scholar]
- Švec, Z. A New Sub-Saharan Leiodes Latreille Species and New Faunistic Data on African Zeadolopus Broun (Coleoptera, Leiodidae, Leiodinae). Studies and Reports of District Museum Prague-East Taxonomical Series 2008. Available online: https://sar.fld.czu.cz/cache/article-data/SaR/Published_volumes/2021-1/141-154_svec_2_kor_2.pdf (accessed on 2 February 2022).
- Newton, A.F. Mycophagy in Staphylinoidea (Coleoptera). In Fungus-Insect Relatuionships: Perspectives in Ecology and Evolution; Wheeler, Q.D., Blackwell, M., Eds.; Columbia University Press: New York, NY, USA, 1984; pp. 302–353. [Google Scholar]
- Cooter, J. Leiodidae. In A Coleopterist’s Handbook, 4th ed.; Cooter, J., Barclay, M.V.L., Eds.; Amateur Entomologists’ Society: Orpington, UK, 2006. [Google Scholar]
- Menta, C.; Pinto, S. Biodiversity and Ecology of Soil Fauna in Relation to Truffle. In True Truffle (Tuber spp.) in the World. Soil Ecology, Systematics and Biochemistry; Zambonelli, A., Iotti, M., Murat, C., Eds.; Springer International Publishing: Berlin/Heidelberg, Germany, 2016; pp. 319–331. [Google Scholar]
- Arzone, A. Nuovi reperti sulla biologia di Liodes cinnamomea Panzer in Tuber magnatum Pico (Coleoptera, Staphylinoidea). In Allionia: Bollettino dell’Instituto ed Orto Botanico dell’Universitaà di Torino. 1971. Available online: https://books.google.com/books/about/Nuovi_reperti_sulla_biologia_di_Liodes_c.html?id=dco7MwEACAAJ (accessed on 2 February 2022).
- Arzone, A. Reperti ecologici ed etologici di Liodes cinnamomea Panzer vivente su Tuber melanosporum Vittadini (Coleoptera Staphylinoidea). In Annali della Facoltà di Scienze Agrarie della Università degli Studi di Torino; Università di Torino. Facoltà di scienze agrarie: Grugliasco, Italy, 1970; Volume 5, pp. 317–357. [Google Scholar]
- Hochberg, M.E.; Bertault, G.M.; Poitrineau, K.; Janssen, A. Olfactory orientation of the truffle beetle, Leiodes cinnamomea. Entomol. Exp. Appl. 2003, 109, 147–153. [Google Scholar] [CrossRef]
- Donisthorpe, H. The British Fungicolous Coleoptera. Entomol.’s Mon. Mag. 1935, 71, 21–31. [Google Scholar]
- Benick, L. Pilzkäfer und Käferpilzen. Ökologie und statische Untersuchngen. Acta Zool. Fenn. 1952, 70, 1–250. Available online: https://www.worldcat.org/title/pilzkafer-und-kaferpilze-okologische-und-statistische-untersuchungen/oclc/5065200?referer=di&ht=edition (accessed on 2 February 2022).
- Cooter, J. Observations on Leiodes species with particular reference to Leiodes cinnamomea (Panzer, 1793) in a truffle wood (Coleoptera: Leiodidae). Entomol.’s Mon. Mag. 2015, 151, 289–290. [Google Scholar]
- Rosa-Gruszecka, A.; Hilszczańska, D.; Plewa, R.; Szmidla, H. Insects associated with truffles in Poland. In Proceedings of the 6th TAUESG (Tuber aestivum/uncinatum European Scientists Group) Conference, Norwich, UK, 26–28 August 2015. [Google Scholar]
- Lyszkowski, R.M. The larval habitat of Leiodes rufipennis (Paykull) (=clavicornis (Rye) (Col.: Leiodidae): Some preliminary observations. Entomol. Rec. J. Var. 1995, 107, 39–40. Available online: https://www.biodiversitylibrary.org/part/195539 (accessed on 2 February 2022).
- Laboulbène, A. Observations sur les insectes tubérivores. Avec refutation de l’erreur qui, attribuant les Truffes a la piqùre d’un Insecte, les a fait assimiler aux Galles végétales. Ann. Soc. Entomol. Fr. 1864, 4, 69–114. [Google Scholar]
- Klausnitzer, B. Ordnung Coleoptera (Larven). In Die Larven der Käfer Mitteleuropas. Band 4. Polyphaga. Teil 3. Leiodidae; Akademie-Verlag: Berlin, Germany, 1978; pp. 91–92. [Google Scholar]
- Paolocii, F.; Rubbini, A.; Riccioni, C.; Arcioni, S. Reevaluation of the life cycle of Tuber magnatum. Appl. Environ. Microbiol. 2006, 72, 2390–2393. [Google Scholar] [CrossRef] [Green Version]
- Ashe, J.S.; Watrous, L.E. Larval chaetotaxy of Aleocharinae (Staphylinidae) based on a description of Atheta coriaria Kraatz. Coleop. Bull. 1984, 38, 165–179. [Google Scholar]
- Wheeler, Q.D. Morphology and ontogeny of postembryonic larval Agathidium and Anisotoma (Coleoptera: Leiodidae). Am. Mus. Novit. 1990, 2986, 46. [Google Scholar]
- Kilian, A. Morphology and phylogeny of the larval stages of the tribe Agathidiini (Coleoptera: Leiodidae: Leiodinae). Ann. Zool. 1998, 48, 125–220. [Google Scholar]
- Kilian, A. Phylogenetic analysis of Camiarinae (Coleoptera: Leiodidae) based on larval morphology. Ann. Zool. 2012, 62, 11–68. [Google Scholar] [CrossRef]
- Newton, A.F. Leiodidae Fleming, 1821. In Handbook of Zoology. Arthropoda: Insecta. Coleoptera, Beetles. Volume 1: Morphology and Systematics (Archostemata, Adephaga, Myxophaga, Polyphaga partim), 2nd ed.; Beutel, R.G., Leschen, R.A.B., Eds.; De Gruyter: Berlin, Germany, 2016; pp. 364–376. [Google Scholar]
- Chandler, P.J. Bradysia smithae Menzel & Heller, 2005, (Diptera: Sciaridae) reared from truffles, a new host association. Entomol.’s Mon. Mag. 2015, 151, 292. [Google Scholar]
- Lawrence, J.F. Mycophagy in the Coleoptera: Feeding strategies and morphological adaptations. In Insect-Fungus Interactions; Wilding, N., Collins, N.M., Hammond, P.M., Webber, J.F., Eds.; Royal Entomological Society Symposium Series 14; Academic Press: London, UK, 1989; pp. 1–23, xvi + 344p. [Google Scholar]
- Newton, A.F., Jr. Leiodidae. Round fungus Beetles, Small Carrion Beetles and Allies. In Immature Insects; Stehr, F.W., Ed.; Kendall/Hunt Pub. Co.: Dubuque, IA, USA, 1991; Volume 2, pp. 327–329. [Google Scholar]
- Wheeler, Q.D. Revision and cladistics of the Middle American genus Creagrophorus Matthews (Coleoptera: Leiodidae). Quaest. Entomol. 1979, 15, 447–479. [Google Scholar]
- Wheeler, Q.D. Larval characters of Neotropical Scotocryptus (Coleoptera: Leiodidae), a nest associate of stingless bees (Hymenoptera: Apidae). J. N. Y. Entomol. Soc. 1985, 93, 1082–1088. [Google Scholar]
- Costa, C.; Vanin, S.A.; Casari-Chen, S.A. Larvas de Coleoptera do Brasil; Museum de Zoologia Universidade de Sao Paulo: São Paulo, Brazil, 1988; 282p. [Google Scholar]
- Ratajczak, A. Redescription of the third larval instar of Anisotoma humeralis (F.) (Coleoptera: Leiodidae). Genus 1995, 6, 277–288. [Google Scholar]
- Ratajczak, A. Description of the third larval stage of Anisotoma orbicularis (Herbst) and redescription of the third larval stage of A. castanea (Herbst) (Coleoptera: Leiodidae). Genus 1996, 7, 351–378. [Google Scholar]
- Růžička, J. Description of the third instar larvae of Anisotoma axillaris and A. glabra (Coleoptera: Leiodidae), with a key to larvae of European Anisotoma species. Acta Soc. Zool. Bohem. 1996, 60, 455–468. [Google Scholar]
- Kilian, A. A description of the larva of Agathidium kumaonicum Angelini et De Marzo, 1985 (Coleoptera: Leiodidae). Genus 2001, 12, 563–575. [Google Scholar]
- Kilian, A. Studies on the morphology of immature stages of the tribe Agathidiini (Coleoptera: Leiodidae). Part I. Anisotoma discolor Melsheimer, 1844. Ann. Zool. 2003, 53, 655–661. [Google Scholar]
- Kilian, A. Studies on the morphology of immature stages of the tribe Agathidiini (Coleoptera: Leiodidae). Part II. Anisotoma blanchardi. Eur. J. Entomol. 2005, 102, 97–105. [Google Scholar] [CrossRef] [Green Version]
- Kilian, A. Studies on the morphology of immature stages of the tribe Agathidiini (Coleoptera: Leiodidae). Part III. Anisotoma horni Wheeler, 1979. Ann. Zool. 2005, 55, 45–56. [Google Scholar]
- Kilian, A. Studies on the morphology of immature stages of the tribe Agathidiini (Coleoptera: Leiodidae). Part IV. Agathidium angulare Mannerheim 1852. Ann. Zool. 2006, 56, 587–600. [Google Scholar]
- Grebennikov, V.V.; Leschen, R.A.B. External exoskeletal cavities in Coleoptera and their possible mycangial functions. Entomol. Sci. 2010, 13, 81–98. [Google Scholar] [CrossRef]
- Kilian, A.; Ptak, M. Larval morphology of Neocamiarus kuscheli kuscheli Jeannel 1957 and Inocatops sp. (Coleoptera, Leiodidae, Camiarinae): The first modern descriptions of two members of the tribe Camiarini in comparison with other Camiarinae. Aust. J. Entomol. 2013, 52, 320–337. [Google Scholar] [CrossRef]
- Peck, S.B. The eyeless inquiline genera Parabystus and Scotocryptus of Costa Rica and Panama; Inhabitants of nests of stingless bees (Coleptera: Leiodidae). Sociobiology 2003, 42, 65–80. [Google Scholar]
- Betz, O.; Thayer, M.; Newton, A.F. Comparative morphology and evolutionary pathways of the mouthparts in spore–feeding Staphylinoidea (Coleoptera). Acta Zool. 2003, 84, 179–238. [Google Scholar] [CrossRef]
- Young, D.K. Synchroidae (Tenebrionoidea). In Immature Insects; Stehr, F.W., Ed.; Michigan State University: East Lansing, MI, USA; Kendall/Hunt Pub. Co.: Dubuque, IA, USA, 1991; pp. 516–518. [Google Scholar]
- Leschen, R.A.B. Evolutionary Patterns of Feeding in selected Staphylinoidea (Coleptera): Shifts among Food Textures. In Functional Morphology of Insect Feeding; Schaefer, C.W., Leschen, R.A.B., Eds.; Thomas Say Publication in Entomology; Entomological Society of America: Lanham, MD, USA, 1993; pp. 59–104. [Google Scholar]
- Kadowaki, K.; Leschen, R.A.B.; Beggs, J.R. Spore consumption and life history of Zearagytodes maculifer (Broun) (Coleoptera, Leiodidae) on Ganoderma, its fungal host. N. Z. J. Ecol. 2010, 35, 61–68. [Google Scholar]
- Kadowaki, K.; Leschen, R.A.B.; Beggs, J.R. No evidence for a Ganoderma spore dispersal mutualism in an obligate spore-feeding beetle Zearagytodes maculifer. Fungal Biol. 2011, 115, 768–774. [Google Scholar] [CrossRef]
- Lawrence, J.F. Order Coleoptera. In Immature Insects; Stehr, F.W., Ed.; Michigan State University: East Lansing, MI, USA; Kendall/Hunt Pub. Co.: Dubuque, IA, USA, 1991; pp. 144–184. [Google Scholar]
- Gutowski, J.M. Wydawanie Dźwięków Przez Owady. Sound Emmissions in Insects. Pr. Inst. Badaw. Leśnictwa Ser. A 1998, 851, 189–217. Available online: https://agro.icm.edu.pl/agro/element/bwmeta1.element.agro-article-8fe8c24f-e99a-49e6-982c-dc443c4dc1d8 (accessed on 2 February 2022).
- Garcia-Montero, L.G.; Pérez Andueza, G.; Diaz, P.; Manjón, J.L. Contribución al conociemiento ecológico de los dípteros fitófagos (Diptera: Heleomyzidae) de las trufas en Espaňa. Bol. San. Veg. Plagas 2004, 30, 679–683. [Google Scholar]
- Leschen, R.A.B. Ecological and behavioral correlates among mycophagous Coleoptera. Folia Entomol. Mex. 1994, 92, 9–19. [Google Scholar]
- Kilian, A.; Kleina, A. Comparative morphology of larval Camiarinae (Coleoptera, Leiodidae). Part II. Agyrtodes, Dictydiella and Zearagytodes, with a review of characters of known larvae of Agyrtodini. Ann. Zool. 2011, 61, 381–420. [Google Scholar] [CrossRef]
- Böving, A.G.; Craighead, F.C. An illustrated synopsis of the principal larval forms of the Order Coleoptera. Entomol. Am. 1931, 11, 1–351. [Google Scholar]

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Instar I | Instar II | Instar III | |
|---|---|---|---|
| total length (mm) | 4.5 | 6.15 | 10 |
| HW/HL | 1.15 | 1.22 | 1.34 |
| HW (mm) | 0.81–0.88 (av. 0.84) | 1.25–1.42 (av. 1.34) | 1.6 |
| HL (mm) | 0.69–0.78 (av. 0.73) | 0.90–1.28 (av. 1.1) | 1.00–1.44 (av. 1.19) |
| N1W (mm) | 0.75 | 1.28 | 1.92 |
| N2W (mm) | 0.7 | 0.98 | 1.28 |
| N3W (mm) | 0.67 | 0.8 | 0.96 |
| A1W (mm) | 0.51 | 1.2 | 1.76 |
| N1L (mm) | 0.35 | 0.48 | 0.64 |
| N2L (mm) | 0.22 | 0.24 | 0.24 |
| N3L (mm) | 0.16 | 0.24 | 0.32 |
| A1L (mm) | 0.13 | 0.24 | 0.32 |
| N1L/W | 0.47 | 0.38 | 0.33 |
| N2L/W | 0.31 | 0.25 | 0.19 |
| N3L/W | 0.24 | 0.3 | 0.33 |
| A1L/W | 0.25 | 0.2 | 0.18 |
| Antenna length (mm) | 0.29 | 0.4 | 0.55 |
| AI/AII/AIII/SA | 3.3/3.7/1.3/1 | 3.9/4.7/1/1 | 4.8/3.8/1.6 |
| Labrum width (mm) | 0.1 | 0.19 | 0.32 |
| Labrum length (mm) | 0.13 | 0.22 | 0.37 |
| Mandible width (mm) | L 0.29/R 0.36 | L 0.34/R 0.36 | L 0.48/R 0.56 |
| Mandible length (mm) | L 0.50/R 0.48 | L 0.65/R 0.60 | L 0.88/R 0.73 |
| MPI/MPII/MPIII | 2.5/1/2.3 | 1.8/1/1.7 | 2/1/1.4 |
| LPI/LPII | 1.74 | 1.84 | 1.6 |
| Urogomphal length (mm) | 0.2 | 0.25 | 0.31 |
| URI/URII/URS | 1.6/1.6/1 | 1.4/1.8/1 | 2.1/2.1/1 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Kilian, A.; Kadej, M.; Cooter, J.; Harvey, D.J. Larval Morphological Adaptations of Leiodes cinnamomea (Panzer, 1793) (Coleoptera: Leiodidae: Leiodinae)—Obligatory Feeder of Tuber Species. Insects 2022, 13, 249. https://doi.org/10.3390/insects13030249
Kilian A, Kadej M, Cooter J, Harvey DJ. Larval Morphological Adaptations of Leiodes cinnamomea (Panzer, 1793) (Coleoptera: Leiodidae: Leiodinae)—Obligatory Feeder of Tuber Species. Insects. 2022; 13(3):249. https://doi.org/10.3390/insects13030249
Chicago/Turabian StyleKilian, Aleksandra, Marcin Kadej, Jonathan Cooter, and Deborah J. Harvey. 2022. "Larval Morphological Adaptations of Leiodes cinnamomea (Panzer, 1793) (Coleoptera: Leiodidae: Leiodinae)—Obligatory Feeder of Tuber Species" Insects 13, no. 3: 249. https://doi.org/10.3390/insects13030249