
Interactions between Rice Resistance to Planthoppers and Honeydew-Related Egg Parasitism under Varying Levels of Nitrogenous Fertilizer


Abstract
:Simple Summary
Abstract
1. Introduction
2. Materials and Methods
2.1. Herbivore Species
2.2. Plant Materials
2.3. Field Plot Design
2.4. Sampling of Free-Living Insects
2.5. Parasitism of Planthopper and Leafhopper Eggs
2.6. Honeydew and Its Effects on Egg Parasitism
2.7. Statistical Analyses
3. Results
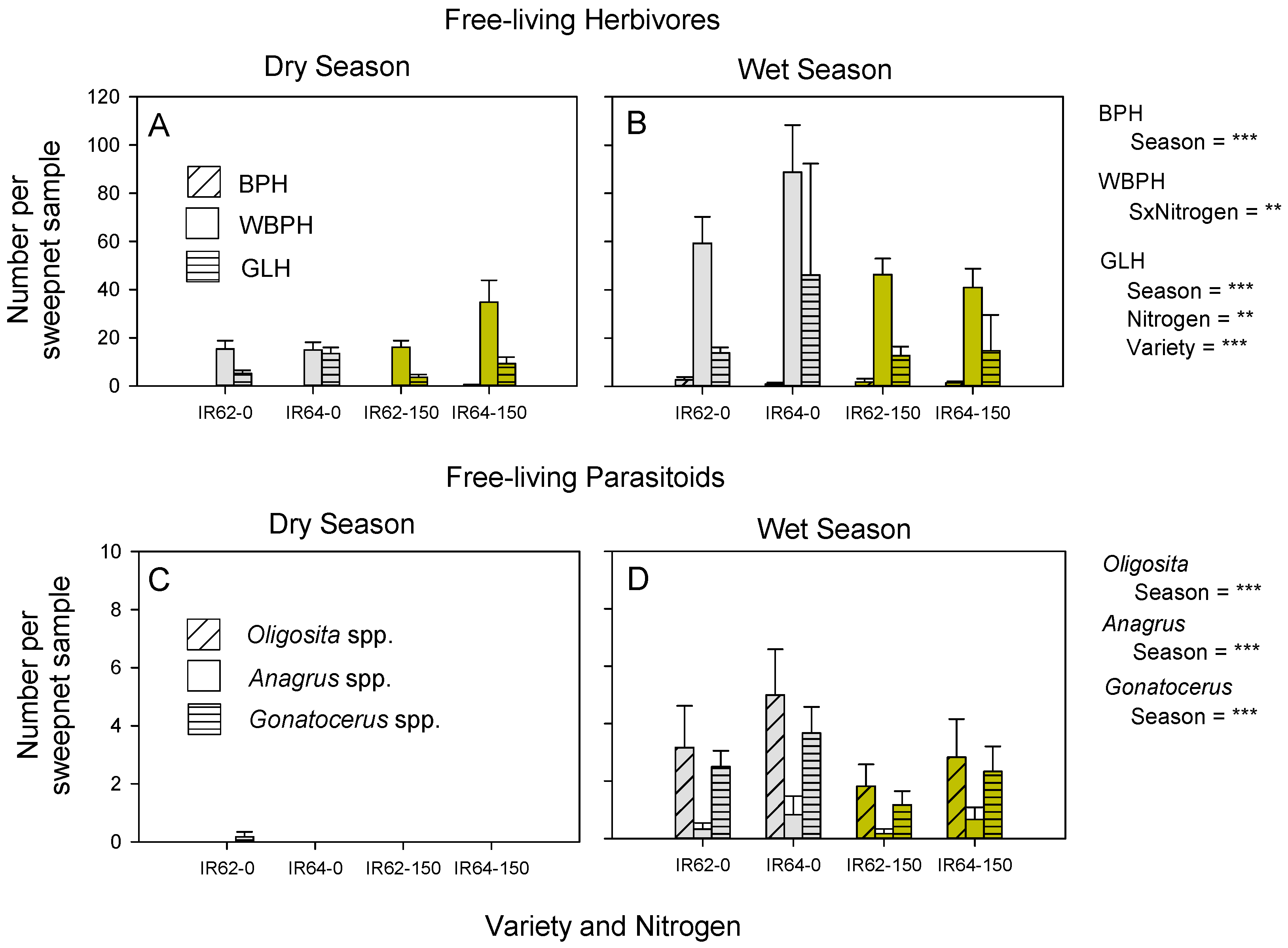
3.1. Arthropod Community
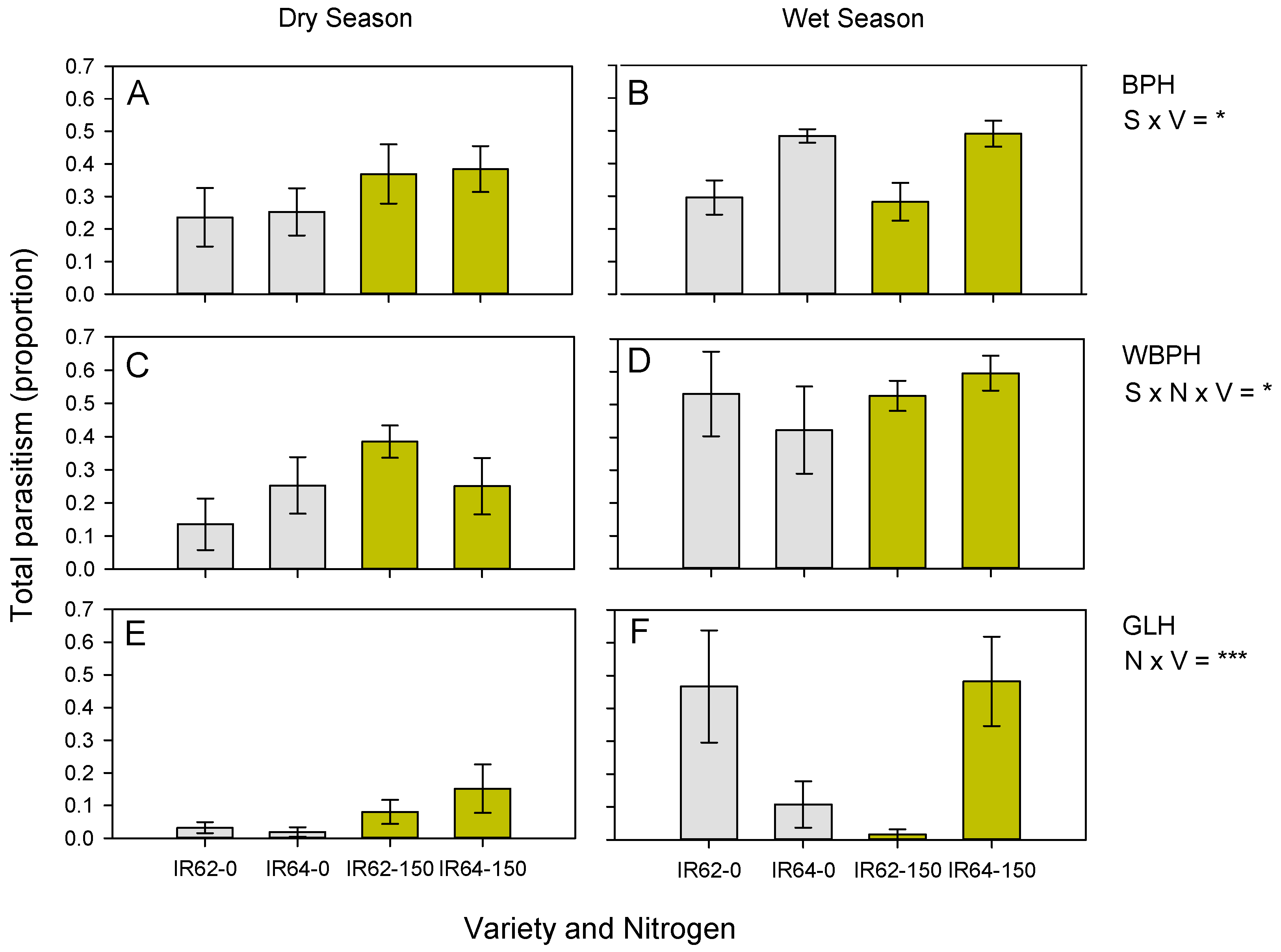
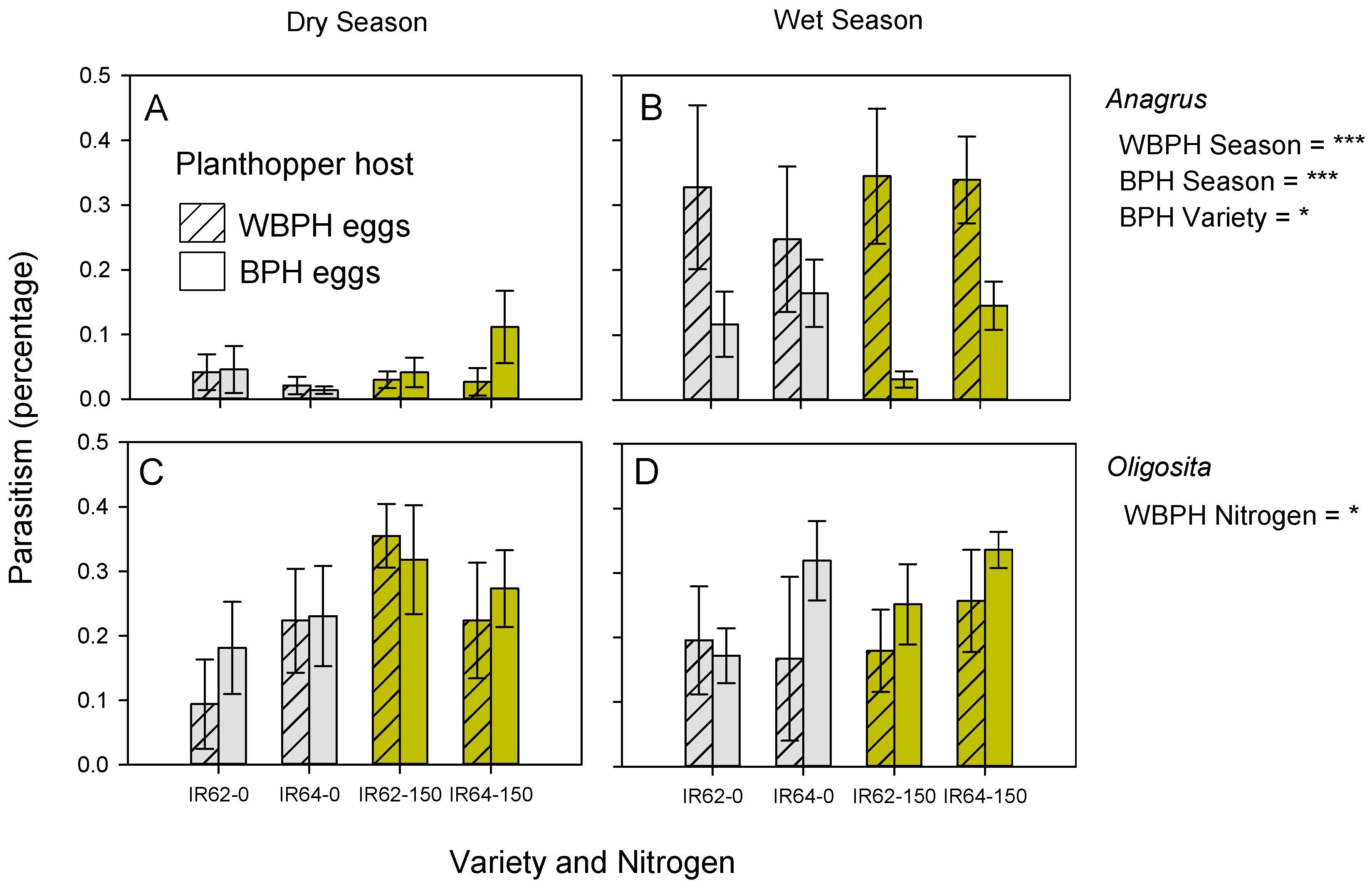
3.2. Parasitism of Planthopper and Leafhopper Eggs
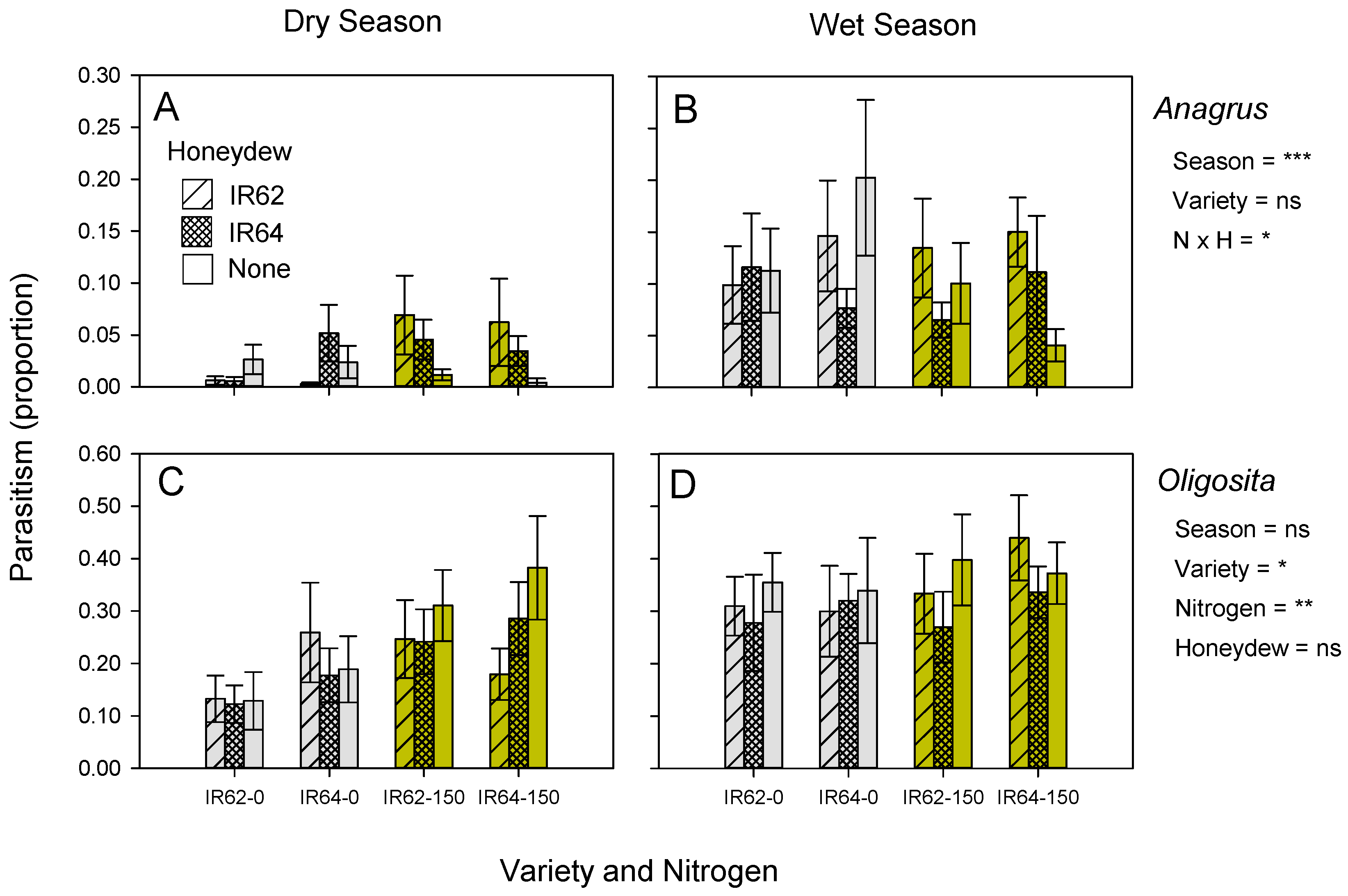
3.3. Honeydew and Its Effects on Parasitism
4. Discussion
4.1. Variety and Nitrogen Effects on Planthopper and Leafhopper Abundance
4.2. Nitrogen Effects on Egg Parasitoids
4.3. Variety Effects on Egg Parasitoids
4.4. The Role of Honeydew in Egg Parasitism
4.5. Incorporating Resistance into Agroecological Designs for Rice Ecosystems
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Muthayya, S.; Sugimoto, J.D.; Montgomery, S.; Maberly, G.F. An overview of global rice production, supply, trade, and consumption. Ann. N. Y. Acad. Sci. 2014, 1324, 7–14. [Google Scholar] [CrossRef] [PubMed]
- Maclean, J.L.; Dawe, D.C.; Hettel, G.P. Rice Almanac; International Rice Research Institute: Metro Manila, Philippines, 2002. [Google Scholar]
- Horgan, F.G. Integrated pest management for sustainable rice cultivation: A holistic approach. In Achieving Sustainable Cultivation of Rice: Cultivation, Pest and Disease Management; Sasaki, T., Ed.; Burleigh Dodds Scientific: Cambridge, UK, 2017; pp. 309–342. [Google Scholar]
- Hu, X.-F.; Cheng, C.; Luo, F.; Chang, Y.-Y.; Teng, Q.; Men, D.-Y.; Liu, L.; Yang, M.-Y. Effects of different fertilization practices on the incidence of rice pests and diseases: A three-year case study in Shanghai, in subtropical southeastern China. Field Crop. Res. 2016, 196, 33–50. [Google Scholar] [CrossRef]
- Lu, Z.; Villareal, S.; Yu, X.-P.; Heong, K.L.; Hu, C. Biodiversity and dynamics of planthoppers and their natural enemies in rice fields with different nitrogen regimes. Rice Sci. 2006, 13, 218. [Google Scholar]
- Awmack, C.S.; Leather, S.R. Host plant quality and fecundity in herbivorous insects. Annu. Rev. Entomol. 2002, 47, 817–844. [Google Scholar] [CrossRef] [PubMed]
- Heinrichs, E.A.; Viajante, F.; Aquino, G. Resurgence-inducing insecticides as a tool in field screening of rice against the brown planthopper. Int. Rice Res. Newsl. 1978, 3, 10–11. [Google Scholar]
- Heinrichs, E.A.; Reissig, W.H.; Valencia, S.; Chellliah, S. Rates and effect of resurgence-inducing insecticides on populations of Nilaparvata lugens (Homoptera: Delphacidae) and its predators. Environ. Entomol. 1982, 11, 1269–1273. [Google Scholar] [CrossRef] [Green Version]
- Wu, J.; Ge, L.; Liu, F.; Song, Q.; Stanley, D. Pesticide-induced planthopper population resurgence in rice cropping systems. Annu. Rev. Entomol. 2002, 65, 409–429. [Google Scholar] [CrossRef] [Green Version]
- Wu, J.; Wang, A.; Xu, J.; Yang, G.; Qiu, H.; Li, D.H. Studies on stimulating effect of two selective insecticides on the number of egg laid by rice yellow borer, Tryporyza incertulas (Walker) and their effects on biochemistry of rice plants. Sci. Agric. Sin. 2003, 36, 1163–1170. [Google Scholar]
- Wu, J.-C.; Qiu, H.-M.; Yang, G.-Q.; Liu, J.-L.; Liu, G.-J.; Wilkins, R.M. Effective duration of pesticide-induced susceptibility of rice to brown planthopper (Nilaparvata lugens Stål, Homoptera: Delphacidae), and physiological and biochemical changes in rice plants following pesticide application. Int. J. Pest Manag. 2004, 50, 55–62. [Google Scholar] [CrossRef]
- Wu, J.-C.; Xu, J.-X.; Yuan, S.-Z.; Liu, J.-L.; Jiang, Y.-H.; Xu, J.-F. Pesticide-induced susceptibility of rice to brown planthopper Nilaparvata lugens. Entomol. Exp. Appl. 2001, 100, 119–126. [Google Scholar] [CrossRef] [Green Version]
- Chelliah, S.; Heinrichs, E.A. Factors affecting insecticide-induced resurgence of the brown planthopper, Nilaparvata lugens on Rice. Environ. Entomol. 1980, 9, 773–777. [Google Scholar] [CrossRef]
- Fabellar, L.T.; Heinrichs, E.A. Toxicity of insecticides to predators of rice brown planthoppers, Nilaparvata lugens (Stål) (Homoptera: Delphacidae). Environ. Entomol. 1984, 13, 832–837. [Google Scholar] [CrossRef]
- Fabellar, L.T.; Heinrichs, E.A. Relative toxicity of insecticides to rice planthoppers and leafhoppers and their predators. Crop Prot. 1986, 5, 254–258. [Google Scholar] [CrossRef]
- Cuong, N.L.; Ben, P.T.; Phuong, L.T.; Chau, L.M.; Cohen, M.B. Effect of host plant resistance and insecticide on brown planthopper Nilaparvata lugens (Stål) and predator population development in the Mekong Delta, Vietnam. Crop Prot. 1997, 16, 707–715. [Google Scholar] [CrossRef]
- Fujita, D.; Kohli, A.; Horgan, F.G. Rice resistance to planthoppers and leafhoppers. Crit. Rev. Plant Sci. 2013, 32, 162–191. [Google Scholar] [CrossRef]
- Horgan, F.G.; Almazan, M.-L.P.; Vu, Q.; Ramal, A.F.; Bernal, C.C.; Yasui, H.; Fujita, D. Unanticipated benefits and potential ecological costs associated with pyramiding leafhopper resistance loci in rice. Crop Prot. 2019, 115, 47–58. [Google Scholar] [CrossRef]
- Widawsky, D.; Rozelle, S.; Jin, S.; Huang, J. Pesticide productivity, host-plant resistance and productivity in China. Agric. Econ. 1998, 19, 203–217. [Google Scholar]
- Gurr, G.M.; Lu, Z.; Zheng, X.; Xu, H.; Zhu, P.; Chen, G.; Yao, X.; Cheng, J.; Zhu, Z.; Catindig, J.L.; et al. Multi-country evidence that crop diversification promotes ecological intensification of agriculture. Nat. Plants 2016, 2, 16014. [Google Scholar] [CrossRef]
- Ali, M.P.; Bari, M.N.; Haque, S.S.; Kabir, M.M.M.; Afrin, S.; Norwin, F.; Islam, M.S.; Landis, D.A. Establishing next-generation pest control services in rice fields: Eco-agriculture. Sci. Rep. 2019, 9, 10180. [Google Scholar] [CrossRef] [Green Version]
- Horgan, F.G.; Crisol Martínez, E.; Stuart, A.M.; Bernal, C.C.; de Cima Martín, E.; Almazan, M.L.P.; Ramal, A.F. Effects of vegetation strips, fertilizer levels and varietal resistance on the integrated management of arthropod biodiversity in a tropical rice ecosystem. Insects 2019, 10, 328. [Google Scholar] [CrossRef] [Green Version]
- Sattler, C.; Schrader, J.; Flor, R.J.; Keo, M.; Chhun, S.; Choun, S.; Hadi, B.A.; Dettele, J. Reducing pesticides and increasing crop diversification offer ecological and economic benefits for farmers—A case study in Cambodian rice fields. Insects 2021, 12, 267. [Google Scholar] [CrossRef] [PubMed]
- Vu, Q.; Ramal, A.F.; Villegas, J.M.; Jamoralin, A.; Bernal, C.C.; Pasang, J.M.; Almazan, M.L.P.; Ramp, D.; Settele, J.; Horgan, F.G. Enhancing the parasitism of insect herbivores through diversification of habitat in Philippine rice fields. Paddy Water Environ. 2018, 16, 379–390. [Google Scholar] [CrossRef]
- Shanker, C.; Mohan, M.; Sampathkumar, M.; Lydia, C.; Katti, G. Selection of flowering forbs for conserving natural enemies in rice fields. Biocontrol Sci. Technol. 2013, 23, 480–484. [Google Scholar] [CrossRef]
- Zhu, P.; Gurr, G.M.; Lu, Z.; Heong, K.; Chen, G.; Zheng, X.; Xu, H.; Yang, Y. Laboratory screening supports the selection of sesame (Sesamum indicum) to enhance Anagrus spp. parasitoids (Hymenoptera: Mymaridae) of rice planthoppers. Biol. Control 2013, 64, 83–89. [Google Scholar] [CrossRef]
- Sogawa, K. Studies on the feeding habits of the brown planthopper. II. Honeydew excretion. Jpn. J. Appl. Entomol. Zool. 1970, 14, 134–139. [Google Scholar] [CrossRef]
- Paguia, P.; Pathak, M.D.; Heinrichs, E.A. Honeydew excretion measurement techniques for determining differential feeding activity of biotypes of Nilaparvata lugens on rice varieties. J. Econ. Entomol. 1980, 73, 35–40. [Google Scholar] [CrossRef]
- Pathak, P.K.; Saxena, R.C.; Heinrichs, E.A. Parafilm sachet for measuring honeydew excretion by Nilaparvata lugens on rice. J. Econ. Entomol. 1982, 75, 194–195. [Google Scholar] [CrossRef]
- Yesuraja, I.; Mariappan, I. Feeding behavior of green leafhopper (GLH) on rice varieties resistant to rice tungro. Int. Rice Res. Newsl. 1991, 16, 15. [Google Scholar]
- Horgan, F.G.; de Freitas, T.F.S.; Crisol-Martínez, E.; Mundaca, E.A.; Bernal, C.C. Nitrogenous fertilizer reduces resistance but enhances tolerance to the brown planthopper in fast-growing, moderately resistant rice. Insects 2021, 12, 989. [Google Scholar] [CrossRef]
- Bouchard, Y.; Cloutier, C. Role of olfaction in host finding by aphid parasitoid Aphidius nigripes (Hymenoptera: Aphidiidae). J. Chem. Ecol. 1985, 11, 801–808. [Google Scholar] [CrossRef]
- Fatouros, N.E.; Dicke, M.; Mumm, R.; Meiners, T.; Hilker, M. Foraging behavior of egg parasitoids exploiting chemical information. Behav. Ecol. 2008, 19, 677–689. [Google Scholar] [CrossRef]
- Jonsson, M.; Wratten, S.D.; Landis, D.A.; Gurr, G.M. Recent advances in conservation biological control of arthropods by arthropods. Biol. Control 2008, 45, 172–175. [Google Scholar] [CrossRef]
- Luquet, M.; Peñalver-Cruz, A.; Satour, P.; Anton, S.; Cortesero, A.-M.; Lavandero, B.; Jaloux, B. Aphid honeydew may be the predominant sugar source for Aphidius parasitoids even in nectar-providing intercrops. Biol. Control 2021, 158, 104596. [Google Scholar] [CrossRef]
- Horgan, F.G. Integrating gene deployment and crop management for improved rice resistance to Asian planthoppers. Crop Prot. 2018, 110, 21–33. [Google Scholar] [CrossRef]
- Lou, Y.; Hua, X.; Turlings, T.C.J.; Cheng, J.; Chen, X.; Ye, G. Differences in induced volatile emissions among rice varieties result in differential attraction and parasitism of Nilaparvata lugens eggs by the parasitoid Anagrus nilaparvatae in the field. J. Chem. Ecol. 2006, 32, 2375. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Nalini, R.; Gunathilagaraj, K.; Uthamasamy, S. Interaction among resistant rice genotypes, whitebacked planthopper Sogatella furcifera (Horvath), and egg parasitoid Anagrus nr. flaveolus. Int. Rice Res. Notes 2005, 30, 29–30. [Google Scholar]
- McKey, D. Adaptive patterns in alkaloid physiology. Am. Nat. 1974, 108, 305–320. [Google Scholar] [CrossRef]
- Kvedaras, O.L.; An, M.; Choi, Y.S.; Gurr, G.M. Silicon enhances natural enemy attraction and biological control through induced plant defences. Bull. Entomol. Res. 2010, 100, 367–371. [Google Scholar] [CrossRef] [Green Version]
- Liu, J.; Zhu, J.; Zhang, P.; Han, L.; Reynolds, O.L.; Zeng, R.; Wu, J.; Shao, Y.; You, M.; Gurr, G.M. Silicon supplementation alters the composition of herbivore induced plant volatiles and enhances attraction of parasitoids to infested rice plants. Front. Plant Sci. 2017, 8, 1265. [Google Scholar] [CrossRef] [Green Version]
- Gontijo, L.; Cascone, P.; Giorgini, M.; Michelozzi, M.; Rodrigues, H.S.; Spiezia, G.; Iodice, L.; Guerrieri, E. Relative importance of host and plant semiochemicals in the foraging behavior of Trichogramma achaeae, an egg parasitoid of Tuta absoluta. J. Pest Sci. 2019, 92, 1479–1488. [Google Scholar] [CrossRef]
- Ali, A.N.; Wright, M.G. Response of Trichogramma papilionis to semiochemicals induced by host oviposition on plants. Biol. Control 2021, 154, 104510. [Google Scholar] [CrossRef]
- Fuchsberg, J.R.; Yong, T.-H.; Losey, J.E.; Carter, M.E.; Hoffmann, M.P. Evaluation of corn leaf aphid (Rhopalosiphum maidis; Homoptera: Aphididae) honeydew as a food source for the egg parasitoid Trichogramma ostriniae (Hymenoptera: Trichogrammatidae). Biol. Control 2007, 40, 230–236. [Google Scholar] [CrossRef]
- Wäckers, F.L.; van Rijn, P.C.J.; Heimpel, G.E. Honeydew as a food source for natural enemies: Making the best of a bad meal? Biol. Control 2008, 45, 176–184. [Google Scholar] [CrossRef] [Green Version]
- Tian, J.C.; Wang, G.W.; Romeis, J.; Zheng, X.S.; Xu, H.X.; Zang, L.S.; Lu, Z.X. Different performance of two Trichogramma (Hymenoptera: Trichogrammatidae) species feeding on sugars. Environ. Entomol. 2016, 45, 1316–1321. [Google Scholar] [CrossRef]
- Horgan, F.G.; Srinivasan, T.S.; Bentur, J.S.; Kumar, R.; Bhanu, K.V.; Sarao, P.S.; Chien, H.V.; Almazan, M.L.P.; Bernal, C.C.; Ramal, A.F.; et al. Geographic and research center origins of rice resistance to Asian planthoppers and leafhoppers: Implications for rice breeding and gene deployment. Agronomy 2017, 7, 62. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Horgan, F.G.; Ramal, A.F.; Bentur, J.S.; Kumar, R.; Bhanu, K.V.; Sarao, P.S.; Iswanto, E.H.; Chien, H.V.; Phyu, M.H.; Bernal, C.C.; et al. Virulence of brown planthopper (Nilaparvata lugens) populations from South and South East Asia against resistant rice varieties. Crop Prot. 2015, 78, 222–231. [Google Scholar] [CrossRef] [Green Version]
- Peñalver Cruz, A.; Arida, A.; Heong, K.L.; Horgan, F.G. Aspects of brown planthopper adaptation to resistant rice varieties with the Bph3 gene. Entomol. Exp. Appl. 2011, 141, 245–257. [Google Scholar] [CrossRef]
- Nguyen, C.D.; Zheng, S.-H.; Sanada-Morimura, S.; Matsumura, M.; Yasui, H.; Daisuke, F. Substitution mapping and characterization of brown planthopper resistance genes from indica rice variety, ‘PTB33’ (Oryza sativa L.). Breed. Sci. 2021, 71, 5. [Google Scholar] [CrossRef]
- Mackill, D.J.; Khush, G.S. IR64: A high-quality and high-yielding mega variety. Rice 2018, 11, 18. [Google Scholar] [CrossRef] [Green Version]
- Alam, S.N.; Cohen, M.B. Detection and analysis of QTLs for resistance to the brown planthopper, Nilaparvata lugens, in a double-haploid rice population. Theor. Appl. Genet. 1998, 97, 1370–1379. [Google Scholar] [CrossRef]
- Horgan, F.G.; Peñalver-Cruz, A. Compatibility of insecticides with rice resistance to planthoppers as influenced by the timing and frequency of applications. Insects 2022, 13, 106. [Google Scholar] [CrossRef] [PubMed]
- Horgan, F.G.; Peñalver Cruz, A.; Bernal, C.C.; Ramal, A.F.; Almazan, M.L.P.; Wilby, A. Resistance and tolerance to the brown planthopper, Nilaparvata lugens (Stål), in rice infested at different growth stages across a gradient of nitrogen applications. Field Crop. Res. 2018, 217, 53–65. [Google Scholar] [CrossRef] [PubMed]
- Shepard, B.M.; Barrion, A.T.; Litsinger, J.A. Friends of the Rice Farmer: Helpful Insects, Spiders and Pathogens; International Rice Research Institute: Metro Manila, Philippines, 2000. [Google Scholar]
- R Core Team. R: A Language and Environment for Statistical Computing. R Foundation for Statistical Computing; R-Project: Vienna, Austria, 2020. [Google Scholar]
- Ferrater, J.B.; Naredo, A.I.; Almazan, M.L.P.; de Jong, P.W.; Dicke, M.; Horgan, F.G. Varied responses by yeast-like symbionts during virulence adaptation in a monophagous phloem-feeding insect. Arthropod-Plant Interact. 2015, 9, 215–224. [Google Scholar] [CrossRef]
- Horgan, F.G.; Garcia, C.P.F.; Haverkort, F.; de Jong, P.W.; Ferrater, J.B. Changes in insecticide resistance and host range performance of planthoppers artificially selected to feed on resistant rice. Crop Prot. 2020, 127, 104963. [Google Scholar] [CrossRef] [PubMed]
- Horgan, F.G.; Srinivasan, T.S.; Naik, B.S.; Ramal, A.F.; Bernal, C.C.; Almazan, M.L.P. Effects of nitrogen on egg-laying inhibition and ovicidal response in planthopper-resistant rice varieties. Crop Prot. 2016, 89, 223–230. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Watanabe, T.; Wada, T.; Salleh, N.M.N.b.N. Parasitic activities of egg parasitoids on the rice planthoppers, Nilaparvata lugens (Stål) and Sogatella furcifera (Horváth) (Homoptera: Delphacidae), in the Muda Area, Peninsular Malaysia. Appl. Entomol. Zool. 1992, 27, 205–211. [Google Scholar] [CrossRef] [Green Version]
- Horgan, F.G.; Arida, A.; Ardestani, G.; Almazan, M.L.P. Elevated temperatures diminish the effects of a highly resistant rice variety on the brown planthopper. Sci. Rep. 2021, 11, 262. [Google Scholar] [CrossRef]
- Vu, Q.; Quintana, R.; Fujita, D.; Bernal, C.C.; Yasui, H.; Medina, C.D.; Horgan, F.G. Responses and adaptation by Nephotettix virescens to monogenic and pyramided rice lines with Grh-resistance genes. Entomol. Exp. Appl. 2014, 150, 179–190. [Google Scholar] [CrossRef]
- Zhou, X.; Xyu, H.; Zheng, X.; Yang, Y.; Chen, L.; He, J.; Lyu, Z. Population dynamics of white-backed planthopper and its predatory spiders in hybrid rice fields with different nitrogen regimes. Acta Agric. Zhejiangensis 2012, 24, 865–869. [Google Scholar]
- Horgan, F.G.; Crisol, E. Hybrid rice and insect herbivores in Asia. Entomol. Exp. Appl. 2013, 148, 1–19. [Google Scholar] [CrossRef]
- Claridge, M.F.; Morgan, J.C.; Steenkiste, A.E.; Iman, M.; Damyanti, D. Experimental field studies on predation and egg parasitism of rice brown planthopper in Indonesia. Agric. For. Entomol. 2002, 4, 203–209. [Google Scholar] [CrossRef]
- Sivapragasam, A.; Chua, T.H. Functional response of the brown planthopper egg parasitoids Anagrus nr flaveolas (Waterhouse) and Oligosita sp. (Walker). MARDI Res. J. 1992, 20, 161–165. [Google Scholar]
- Heinrichs, E.A.; Aquino, G.B.; Chelliah, S.; Valencia, S.L.; Reissig, W.H. Resurgence of Nilaparvata lugens (Stål) populations as influenced by method and timing of insecticide applications in lowland rice. Environ. Entomol. 1982, 11, 78–84. [Google Scholar] [CrossRef]
- Ma, M.-Y.; Peng, Z.-P.; He, Y. Effects of temperature on functional response of Anagrus nilaparvatae Pang et Wang (Hymenoptera: Mymaridae) on the eggs of whitebacked planthopper, Sogatella furcifera Horváth and brown planthopper, Nilaparvata lugens Stål. J. Integr. Agric. 2012, 11, 1313–1320. [Google Scholar] [CrossRef]
- Horgan, F.G.; Ramal, A.F.; Villegas, J.M.; Jamoralin, A.; Bernal, C.C.; Perez, M.O.; Pasang, J.M.; Naredo, A.I.; Almazan, M.L.P. Effects of bund crops and insecticide treatments on arthropod diversity and herbivore regulation in tropical rice fields. J. Appl. Entomol. 2017, 141, 587–599. [Google Scholar] [CrossRef]
- Barrion, A.T.; Pantua, P.C.; Bandong, J.P.; dela Cruz, C.G.; Raymundo, F.A.; Lumaban, M.D.; Apostol, R.F.; Litsinger, J.A. Food web of the rice brown planthopper in the Philippines. Int. Rice Res. Notes 1981, 6, 13–15. [Google Scholar]
- Sann, C.; Wemheuer, F.; Beaurepaire, A.; Daniel, R.; Erler, S.; Vidal, S. Preliminary investigation of species diversity of rice hopper parasitoids in Southeast Asia. Insects 2018, 9, 19. [Google Scholar] [CrossRef] [Green Version]
- Lou, Y.-G.; Cheng, J.-A. Host-recognition kairomone from Sogatella furcifera for the parasitoid Anagrus nilaparvatae. Entomol. Exp. Appl. 2001, 101, 59–67. [Google Scholar] [CrossRef]
- Lou, Y.-G.; Du, M.-H.; Turlings, T.C.J.; Cheng, J.-A.; Shan, W.-F. Exogenous application of jasmonic acid induces volatile emissions in rice and enhances parasitism of Nilaparvata lugens eggs by the parasitoid Anagrus nilaparvatae. J. Chem. Ecol. 2005, 31, 1985–2002. [Google Scholar] [CrossRef]
- Lou, Y.-G.; Ma, B.; Cheng, J.-A. Attraction of the parasitoid Anagrus nilaparvatae to rice volatiles induced by the rice brown planthopper Nilaparvata lugens. J. Chem. Ecol. 2005, 31, 2357–2372. [Google Scholar] [CrossRef]
- He, X.; Xu, H.; Gao, G.; Zhou, X.; Zheng, X.; Sun, Y.; Yang, Y.; Tian, J.; Lu, Z. Virus-mediated chemical changes in rice plants impact the relationship between non-vector planthopper Nilaparvata lugens Stål and its egg parasitoid Anagrus nilaparvatae Pang et Wang. PLoS ONE 2014, 9, e105373. [Google Scholar] [CrossRef] [PubMed]
- Wari, D.; Kabir, M.A.; Mujiono, K.; Hojo, Y.; Shinya, T.; Tani, A.; Nakatani, H.; Galis, I. Honeydew-associated microbes elicit defense responses against brown planthopper in rice. J. Exp. Bot. 2019, 70, 1683–1696. [Google Scholar] [CrossRef] [Green Version]
- Kimmins, F.M. Electrical penetration graphs from Nilaparvata lugens on resistant and susceptible rice varieties. Entomol. Exp. Appl. 1989, 50, 69–79. [Google Scholar] [CrossRef]
- Price, P.W.; Bouton, C.E.; Gross, P.; McPheron, B.A.; Thompson, J.N.; Weis, A.E. Interactions among three trophic levels: Influence of plants on interactions between insect herbivores and natural enemies. Annu. Rev. Ecol. Syst. 1980, 11, 41–65. [Google Scholar] [CrossRef] [Green Version]
- Zhu, J.; Zhu, K.; Li, L.; Li, Z.; Qin, W.; Park, Y.; He, Y. Proteomics of the honeydew from the brown planthopper and green rice leafhopper reveal they are rich in proteins from insects, rice plant and bacteria. Insects 2020, 11, 582. [Google Scholar] [CrossRef]
- Shigematsu, Y.; Murofushi, N.; Ito, K.; Kaneda, C.; Kawabe, S.; Takahashi, N. Sterols and asparagine in the rice plant, endogenous factors related to resistance against the brown planthopper (Nilaparvata lugens). Agric. Biol. Chem. 1982, 46, 2877–2879. [Google Scholar]
- Tena, A.; Wäckers, F.L.; Heimpel, G.E.; Urbaneja, A.; Pekas, A. Parasitoid nutritional ecology in a community context: The importance of honeydew and implications for biological control. Curr. Opin. Insect Sci. 2016, 14, 100–104. [Google Scholar] [CrossRef]
- Bargen, H.; Saudhof, K.; Poehling, H.-M. Prey finding by larvae and adult females of Episyrphus balteatus. Entomol. Exp. Appl. 1998, 87, 245–254. [Google Scholar] [CrossRef]
- Dominik, C.; Seppelt, R.; Horgan, F.G.; Settele, J.; Václavík, T. Landscape composition, configuration, and trophic interactions shape arthropod communities in rice agroecosystems. J. Appl. Ecol. 2018, 55, 2461–2472. [Google Scholar] [CrossRef]
- Sann, C.; Theodorou, P.; Heong, K.L.; Villareal, S.; Settele, J.; Vidal, S.; Westphal, C. Hopper parasitoids do not significantly benefit from non-crop habitats in rice production landscapes. Agric. Ecosyst. Environ. 2018, 254, 224–232. [Google Scholar] [CrossRef]
- Calvo-Agudo, M.; Tooker, J.F.; Dicke, M.; Tena, A. Insecticide-contaminated honeydew: Risks for beneficial insects. Biol. Rev. 2022. online early. [Google Scholar] [CrossRef]





{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Honeydew Production | Variety and Nitrogen | χ2-Values 1 | ||||
|---|---|---|---|---|---|---|
| IR62 | IR64 | Variety | Nitrogen | |||
| 0 Added N | 150 kg N ha−1 | 0 Added N | 150 kg N ha−1 | |||
| Total (mm2) | 0.31 ± 0.08 | 0.44 ± 0.09 | 1.20 ± 0.07 | 0.99 ± 0.15 | 51.173 *** | 0.186 ns |
| Xylem Derived (proportion) | 0.85 ± 0.10 | 0.65 ± 0.10 | 0.10 ± 0.03 | 0.20 ± 0.08 | 38.908 *** | 0.284 ns |
| Herbivores 1 | Parasitoids | Variety Effect 2 | Nitrogen Effect 2 | Season Effect 1,2 | Interaction Effects 2 | Egg Density (Covariate) 2 |
|---|---|---|---|---|---|---|
| Abundance in sweep-nets | ||||||
| BPH | ns | ns | WS *** | ns | ||
| WBPH | ns | - | - | Low nitrogen × WS ** | ||
| GLH | IR64 *** | Low ** | WS *** | ns | ||
| Oligosita spp. | ns | ns | WS *** | ns | ||
| Anagrus spp. | ns | ns | WS ** | ns | ||
| Gonatocerus spp. | ns | ns | WS *** | ns | ||
| Parasitism of eggs in sentinel plants | ||||||
| BPH | All combined | - | ns | - | IR64 × WS * | ns |
| WBPH | All combined | - | - | - | Variety × nitrogen × season *,3 | ns |
| GLH | All combined | - | - | ns | IR62 × low nitrogen; IR64 × high nitrogen *** | ns |
| BPH | Oligosita spp. | ns | ns | ns | ns | ns |
| WBPH | Oligosita spp. | ns | High * | ns | ns | ns |
| BPH | Anagrus spp. | IR64 ** | ns | WS *** | ns | ns |
| WBPH | Anagrus spp. | ns | ns | WS *** | ns | ns |
| Parasitism of eggs in sentinel plants (honeydew) | ||||||
| BPH | Oligosita spp. | IR64 ** | High *** | ns | ns | *** |
| BPH | Anagrus spp. | - | ns | WS *** | IR62-derived honeydew × high nitrogen ** | ns |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Peñalver-Cruz, A.; Horgan, F.G. Interactions between Rice Resistance to Planthoppers and Honeydew-Related Egg Parasitism under Varying Levels of Nitrogenous Fertilizer. Insects 2022, 13, 251. https://doi.org/10.3390/insects13030251
Peñalver-Cruz A, Horgan FG. Interactions between Rice Resistance to Planthoppers and Honeydew-Related Egg Parasitism under Varying Levels of Nitrogenous Fertilizer. Insects. 2022; 13(3):251. https://doi.org/10.3390/insects13030251
Chicago/Turabian StylePeñalver-Cruz, Ainara, and Finbarr G. Horgan. 2022. "Interactions between Rice Resistance to Planthoppers and Honeydew-Related Egg Parasitism under Varying Levels of Nitrogenous Fertilizer" Insects 13, no. 3: 251. https://doi.org/10.3390/insects13030251