
Genetic Structure of Apis cerana Populations from South Korea, Vietnam and the Russian Far East Based on Microsatellite and Mitochondrial DNA Polymorphism

 , ,
, ,
Abstract
:Simple Summary
Abstract
1. Introduction
2. Materials and Methods
2.1. Sampling and DNA Extraction
2.2. tRNAleu-COII Locus Analysis
2.3. Analysis of Microsatellite Loci
3. Results
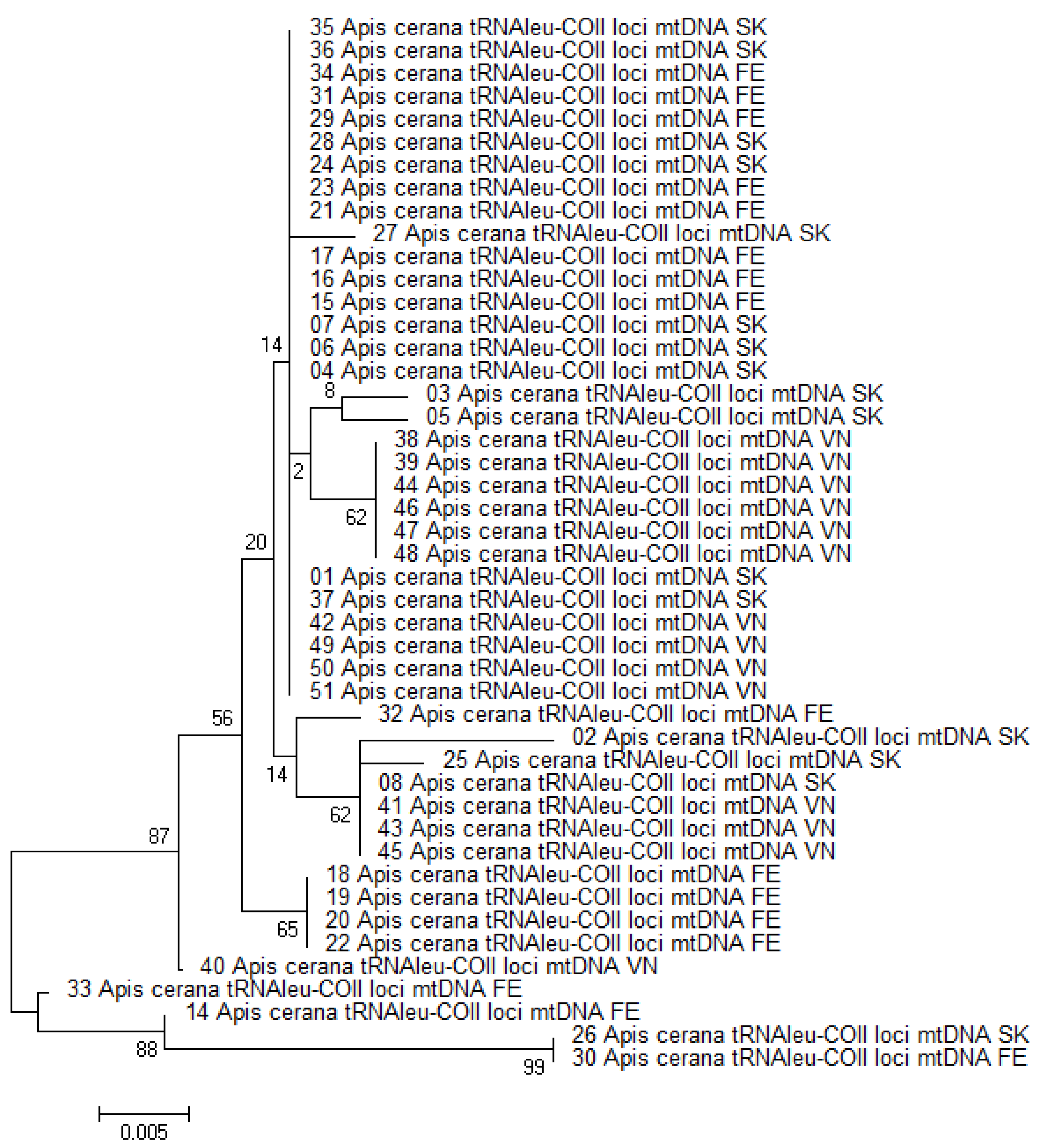
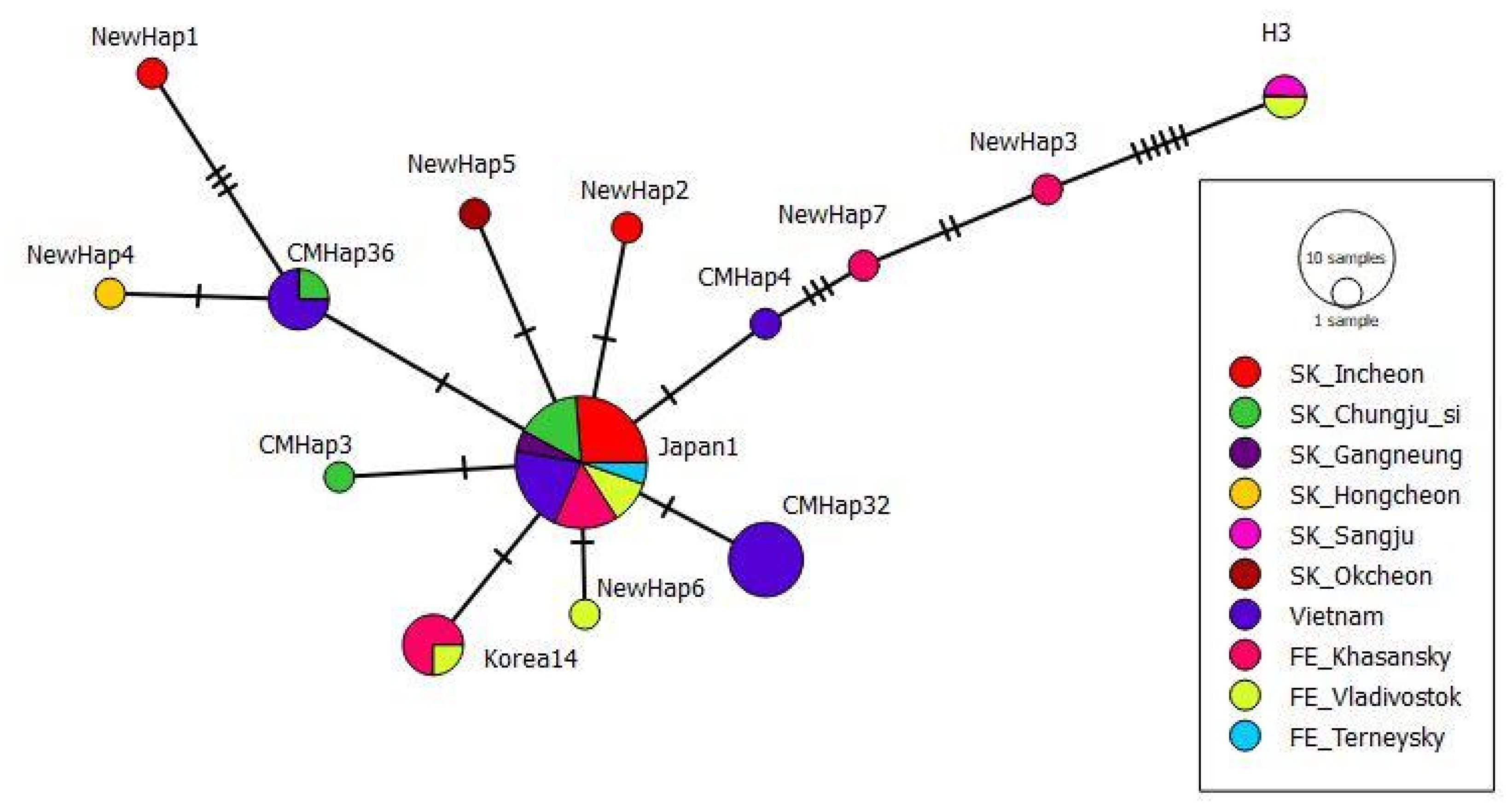
3.1. Haplotype Diversity Based on Polymorphism of tRNAleu-COII Locus
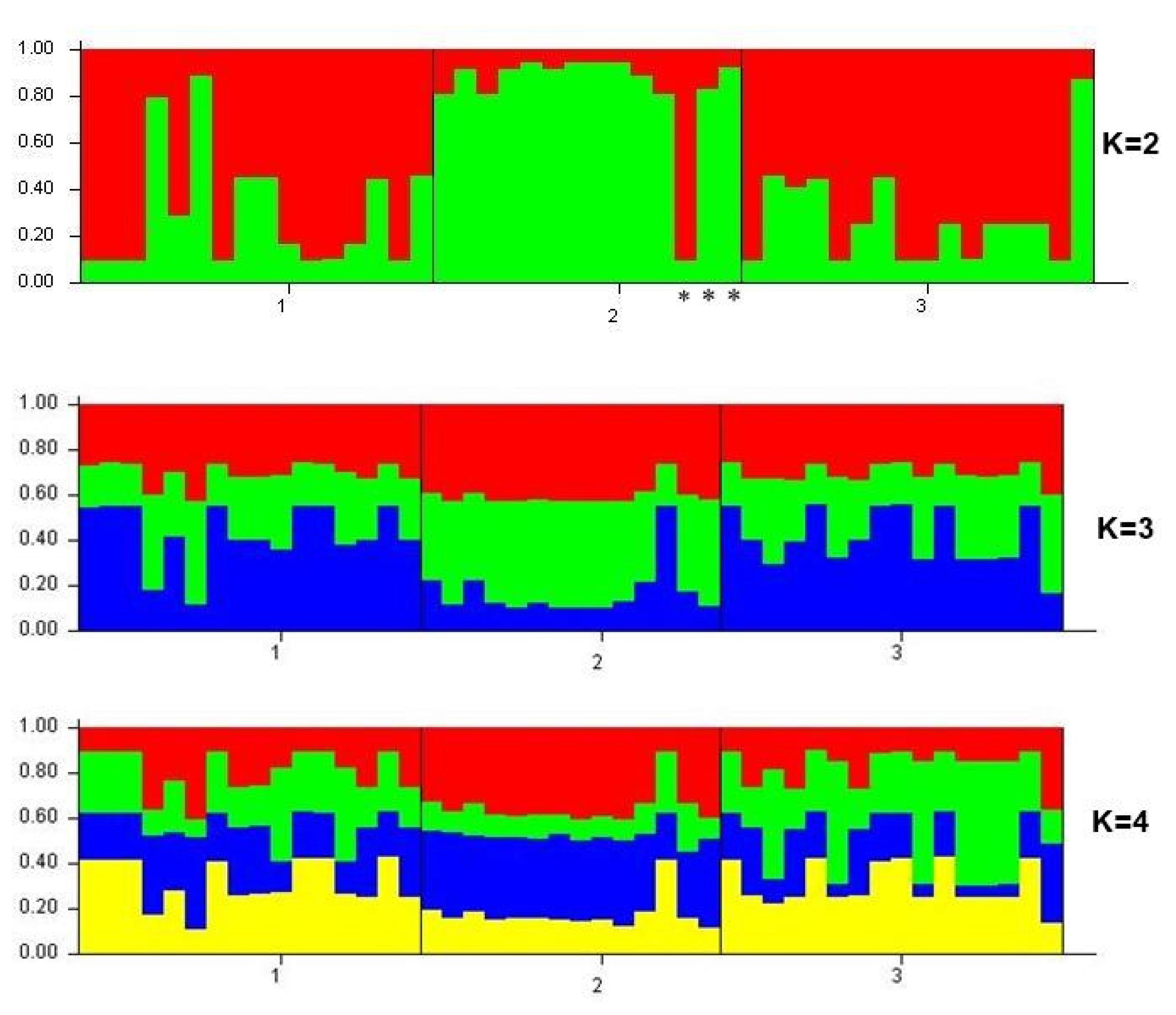
3.2. Genetic Structure of Apis cerana Samples from the Russian Far East, South Korea and Vietnam
4. Discussion
Author Contributions
Funding
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Ruttner, F. Apis cerana Fabricius 1793:327. In Biogeography and Taxonomy of Honeybees; Springer: Berlin/Heidelberg, Germany, 1988. [Google Scholar]
- Peng, Y.S.; Nasr, M.E.; Locke, S.J. Geographical races of Apis cerana Fabricius in China and their distribution. Review of recent Chinese publications and a preliminary statistical analysis. Apidologie 1989, 20, 9–20. [Google Scholar] [CrossRef] [Green Version]
- Radloff, S.E.; Hepburn, R.H.; Hepburn, C.; Fuchs, S.; Otis, G.W.; Sein, M.M.; Aung, H.L.; Pham, H.T.; Tam, D.Q.; Nuru, A.M.; et al. Multivariate morphometric analysis of Apis cerana of southern mainland Asia. Apidologie 2005, 36, 127–139. [Google Scholar] [CrossRef] [Green Version]
- Pesenko, Y.A.; Lelej, A.S.; Radchenko, V.G.; Filatkin, G.N. Chinese wax bee Apis cerana cerana F. (Hymenoptera, Apidae) the Far East of the USSR. Entomol. Rev. 1989, 68, 527–548. [Google Scholar]
- Evans, J.D.; Schwarz, R.S.; Chen, Y.P.; Budge, G.; Cornman, R.S.; De la Rua, P.; de Miranda, J.R.; Foret, S.; Foster, L.; Gauthier, L.; et al. Standard methods for molecular research in Apis mellifera. J. Apic. Res. 2013, 52, 1–55. [Google Scholar] [CrossRef] [Green Version]
- Yang, G.H. The survey of the resource of the Chinese honey bee. Zhongguo Yangfeng 1984, 3, 4–7. [Google Scholar]
- Garnery, L.; Solignac, M.; Celebrano, G.; Cornuet, J.-M. A simple test using restricted PCR-amplified mitochondrial DNA to study the genetic structure of Apis mellifera L. Experientia 1993, 49, 1016–1021. [Google Scholar] [CrossRef]
- Solignac, M.; Vautrin, D.; Loiseau, A.; Mougel, F.; Baudry, E.; Estoup, A.; Garnery, L.; Haberl, M.; Cornuet, J.-M. Five hundred and fifty microsatellite markers for the study of the honeybee (Apis mellifera L.) genome. Mol. Ecol. Notes 2003, 3, 307–311. [Google Scholar] [CrossRef]
- Tan, K.; Warrit, N.; Smith, D.R. Mitochondrial DNA diversity of Chinese Apis cerana. Apidologie 2007, 38, 238–246. [Google Scholar] [CrossRef] [Green Version]
- Zhao, W.; Tan, K.; Zhou, D.; Wang, M.; Cheng, C.; Yu, Z.; Miao, Y.; He, S. Phylogeographic analysis of Apis cerana populations on Hainan Island and southern mainland China, based on mitochondrial DNA sequences. Apidologie 2014, 45, 21–33. [Google Scholar] [CrossRef] [Green Version]
- Lee, J.Y.; Wang, A.R.; Choi, Y.S.; Thapa, R.; Kwon, H.W.; Kim, I. Mitochondrial DNA variations in Korean Apis cerana (Hymenoptera: Apidae) and development of another potential marker. Apidologie 2016, 47, 123–134. [Google Scholar] [CrossRef] [Green Version]
- Xu, X.; Zhu, X.; Zhou, S.; Wu, X.; Zhou, B. Genetic differentiation between Apis cerana cerana populations from Damen Island and adjacent mainland in China. Acta Ecol. Sin. 2013, 33, 122–126. [Google Scholar] [CrossRef]
- Yu, Y.; Zhou, S.; Zhu, X.; Xu, X.; Wang, W.; Zha, L.; Wang, P.; Wang, J.; Lai, K.; Wang, S.; et al. Genetic Differentiation of Eastern Honey Bee (Apis cerana) Populations across Qinghai-Tibet Plateau-Valley Landforms. Front. Genet. 2019, 10, 483. [Google Scholar] [CrossRef] [PubMed]
- Fang, F.; Chen, X.; Lv, J.; Shi, X.; Feng, X.; Wang, Z.; Li, X. Population Structure and Genetic Diversity of Chinese Honeybee (Apis cerana cerana) in Central China. Genes 2022, 13, 1007. [Google Scholar] [CrossRef] [PubMed]
- Thai, P.H.; Nguyen, T.H.; Toan, T.V.; Jung, C. Apis cerana Beekeeping and Sacbrood Disease Management in Vietnam: Review. J. Apic. 2018, 33, 269–275. [Google Scholar] [CrossRef]
- Tamura, K.; Stecher, G.; Kumar, S. MEGA11: Molecular Evolutionary Genetics Analysis Version 11. Mol. Biol. Evol. 2021, 38, 3022–3027. [Google Scholar] [CrossRef] [PubMed]
- Takahashi, J.; Yoshida, T.; Takagi, T.; Akimoto, S.; Wood, K.S.; Deowanish, S.; Hepburn, R.; Nakamura, S.; Matsuka, M. Geographic variation in the Japanese islands of Apis cerana japonica and in A. cerana populations bordering its geographic range. Apidologie 2007, 38, 335–340. [Google Scholar] [CrossRef]
- Gong, X.; Zhao, W.; Zhou, D.; Zhang, X.; Wang, M.; Dong, K.; He, S. Genetic variation and population structure of Apis cerana in northern, central and southern mainland China, based on COXI gene sequences. J. Apic. Res. 2018, 57, 364–373. [Google Scholar] [CrossRef]
- Radloff, S.E.; Hepburn, C.; Hepburn, H.R.; Fuchs, S.; Hadisoesilo, S.; Tan, K.; Engel, M.S.; Kuznetsov, V. Population structure and classification of Apis cerana. Apidologie 2010, 41, 589–601. [Google Scholar] [CrossRef] [Green Version]
- Momeni, J.; Parejo, M.; Nielsen, R.O.; Langa, J.; Montes, I.; Papoutsis, L.; Farajzadeh, L.; Bendixen, C.; Cauia, E.; Charriere, J.-D.; et al. Authoritative subspecies diagnosis tool for European honey bees based on ancestry informative SNPs. BMC Genom. 2021, 22, 101. [Google Scholar] [CrossRef] [PubMed]
- lyasov, R.A.; Youn, H.G.; Lee, M.; Kim, K.W.; Proshchalykin, M.; Lelej, A.S.; Takahashi, J.; Kwon, H.W. Phylogenetic relationships of russian Far-east Apis cerana with other north asian populations. J. Apic. Sci. 2019, 63, 289–314. [Google Scholar]




{kind=link}
{kind=link}
{kind=link}
| NCBI PopSet id | b.p. | N | Number of Haplotypes |
|---|---|---|---|
| 2178350964 | 414 | 2 | 2 |
| 1593526727 | 757 | 72 | 72 |
| 1368163505 | 414 | 10 | 10 |
| 1368162158 | 414 | 2 | 2 |
| 727929638 | 470 | 184 | 10 |
| 87477350 | 432 | 3 | 3 |
| 299829809 | 357 | 10 | 10 |
| 298108794 | 414 | 4 | 4 |
| This study | 431 | 46 | 14 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Kaskinova, M.; Gaifullina, L.; Ilyasov, R.; Lelej, A.; Kwon, H.W.; Thai, P.H.; Saltykova, E. Genetic Structure of Apis cerana Populations from South Korea, Vietnam and the Russian Far East Based on Microsatellite and Mitochondrial DNA Polymorphism. Insects 2022, 13, 1174. https://doi.org/10.3390/insects13121174
Kaskinova M, Gaifullina L, Ilyasov R, Lelej A, Kwon HW, Thai PH, Saltykova E. Genetic Structure of Apis cerana Populations from South Korea, Vietnam and the Russian Far East Based on Microsatellite and Mitochondrial DNA Polymorphism. Insects. 2022; 13(12):1174. https://doi.org/10.3390/insects13121174
Chicago/Turabian StyleKaskinova, Milyausha, Luisa Gaifullina, Rustem Ilyasov, Arkady Lelej, Hyung Wook Kwon, Pham Hong Thai, and Elena Saltykova. 2022. "Genetic Structure of Apis cerana Populations from South Korea, Vietnam and the Russian Far East Based on Microsatellite and Mitochondrial DNA Polymorphism" Insects 13, no. 12: 1174. https://doi.org/10.3390/insects13121174