

Transcriptomic Analyses Suggest the Adaptation of Bumblebees to High Altitudes
 ,
,
Abstract
:Simple Summary
Abstract
1. Introduction
2. Materials and Methods
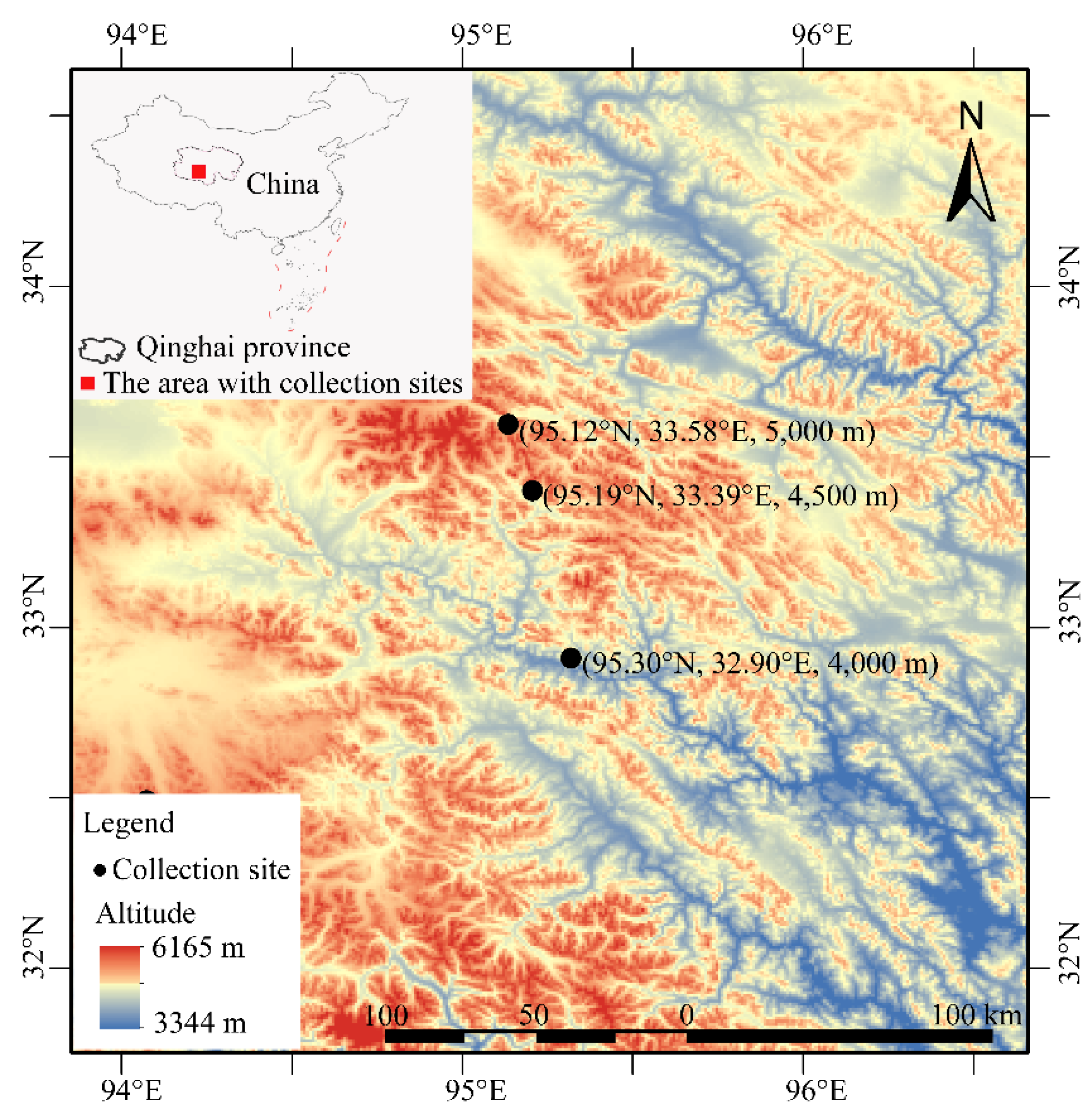
2.1. Transcriptome of Samples
2.2. Identification of Adaptation-Related GO Terms
2.3. KEGG Enrichment of Differentially Expressed Genes
3. Results
3.1. RNA-Seq and Transcriptomic Annotation
3.2. The Gene Set Enrichment Analysis of Gene Expression
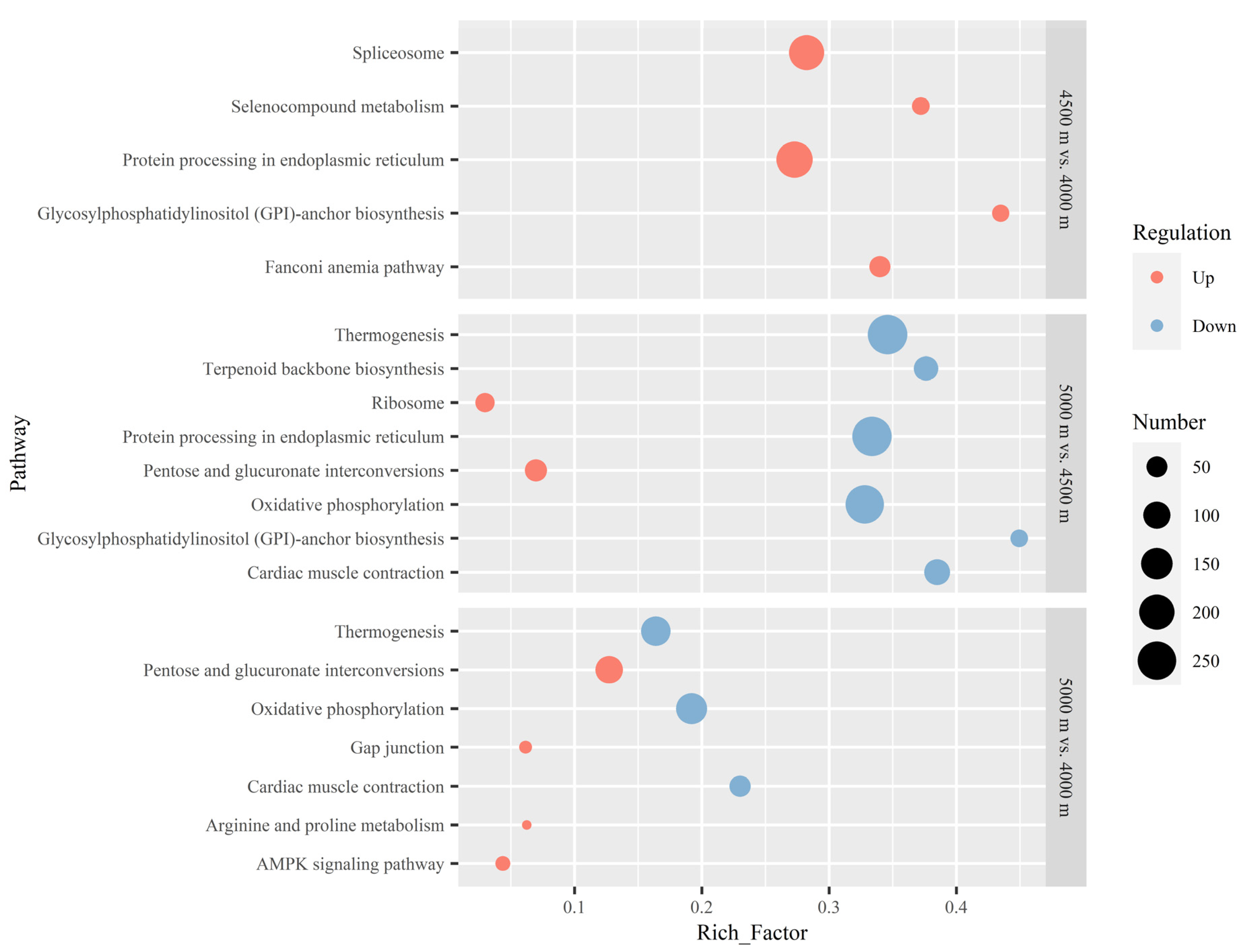
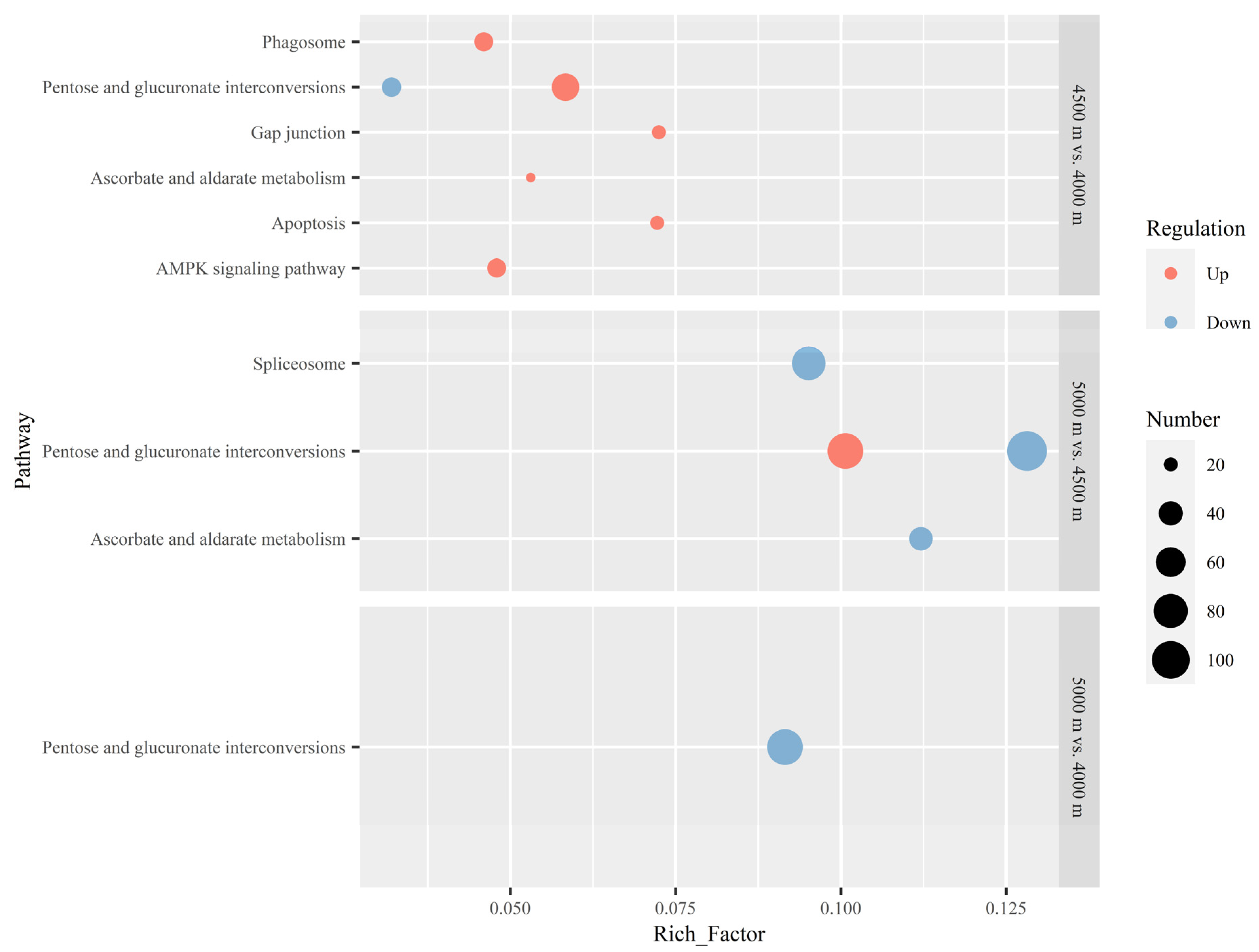
3.3. KEGG Enrichment of DEGs
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional review Board Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Huang, J.X.; An, J.D. Species Diversity, Pollination Application and Strategy for Conservation of the Bumblebees of China. Biodivers. Sci. 2018, 26, 486–497. [Google Scholar] [CrossRef]
- He, Y.P.; Fei, S.M.; Liu, J.Q.; Chen, X.M.; Wang, P.; Jiang, J.M.; He, F. A Preliminary Review of Studies of Alpine Plant Breeding System. J. Sichuan For. Sci. Technol. 2005, 4, 43–49. [Google Scholar] [CrossRef]
- He, J.P.; Liu, J.Q. Pollination Ecology of Gentiana straminea Maxim. (Gentianaceae), an alpine perennial in the Qinghai-Tibet Plateau. Acta Ecol. Sin. 2004, 2, 215–220. [Google Scholar] [CrossRef]
- Naeem, M.; Liu, M.; Huang, J.; Ding, G.; Potapov, G.; Jung, C.; An, J. Vulnerability of East Asian Bumblebee Dpecies to Future Climate and Land Cover Changes. Agric. Ecosyst. Environ. 2019, 277, 11–20. [Google Scholar] [CrossRef]
- Williams, P.H.; Lobo, J.M.; Meseguer, A.S. Bumblebees Take the High Road: Climatically Integrative Biogeography Shows That Escape from Tibet, Not Tibetan Uplift, Is Associated with Divergences of Present-day Mendacibombus. Ecography 2017, 41, 461–477. [Google Scholar] [CrossRef] [Green Version]
- Williams, P.H.; Bystriakova, N.; Huang, J.; Miao, Z.; An, J. Bumblebees, Climate and Glaciers Across the Tibetan plateau (Apidae: Bombus Latreille). Syst. Biodivers. 2015, 13, 164–181. [Google Scholar] [CrossRef]
- Galen, S.C.; Natarajan, C.; Moriyama, H.; Weber, R.E.; Fago, A.; Benham, P.M.; Chavez, A.N.; Cheviron, Z.A.; Storz, J.F.; Witt, C.C. Contribution of a Mutational Hot Spot to Hemoglobin Adaptation in High-altitude Andean house Wrens. Proc. Natl. Acad. Sci. USA 2015, 112, 13958–13963. [Google Scholar] [CrossRef] [Green Version]
- Natarajan, C.; Hoffmann, F.G.; Weber, R.E.; Fago, A.; Witt, C.C.; Storz, J.F. Predictable Convergence in Hemoglobin Function Has Unpredictable Molecular Underpinnings. Science 2016, 354, 336–339. [Google Scholar] [CrossRef] [Green Version]
- Natarajan, C.; Projecto-Garcia, J.; Moriyama, H.; Weber, R.E.; Muñoz-Fuentes, V.; Green, A.J.; Kopuchian, C.; Tubaro, P.L.; Alza, L.; Bulgarella, M. Convergent Evolution of Hemoglobin Function in High-altitude Andean Waterfowl Involves Limited Parallelism at the Molecular Sequence Level. PLoS Genet. 2015, 11, e10056812015-11. [Google Scholar] [CrossRef] [Green Version]
- Projecto-Garcia, J.; Natarajan, C.; Moriyama, H.; Weber, R.E.; Fago, A.; Cheviron, Z.A.; Dudley, R.; McGuire, J.A.; Witt, C.C.; Storz, J.F. Repeated Elevational Transitions in Hemoglobin Function during the Evolution of Andean Hummingbirds. Proc. Natl. Acad. Sci. USA 2013, 110, 20669–20674. [Google Scholar] [CrossRef]
- Zhu, X.; Guan, Y.; Signore, A.V.; Natarajan, C.; DuBay, S.G.; Cheng, Y.; Han, N.; Song, G.; Qu, Y.; Moriyama, H. Divergent and Parallel Routes of Biochemical Adaptation in High-Altitude Passerine Birds from the Qinghai-Tibet Plateau. Proc. Natl. Acad. Sci. USA 2018, 115, 1865–1870. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Simonson, T.S.; Yang, Y.; Huff, C.D.; Yun, H.; Qin, G.; Witherspoon, D.J.; Bai, Z.; Lorenzo, F.R.; Xing, J.; Jorde, L.B. Genetic Evidence for High-Altitude Adaptation in Tibet. Science 2010, 329, 72–75. [Google Scholar] [CrossRef] [Green Version]
- Bigham, A.W. Genetics of Human Origin and Evolution: High-Altitude Adaptations. Curr. Opin. Genet. Dev. 2016, 41, 8–13. [Google Scholar] [CrossRef] [Green Version]
- Gou, X.; Wang, Z.; Li, N.; Qiu, F.; Xu, Z.; Yan, D.; Yang, S.; Jia, J.; Kong, X.; Wei, Z. Whole-genome Sequencing of Six Dog Breeds from Continuous Altitudes Reveals Adaptation to High-altitude Hypoxia. Genome Res. 2014, 24, 1308–1315. [Google Scholar] [CrossRef] [Green Version]
- Jha, A.R.; Zhou, D.; Brown, C.D.; Kreitman, M.; Haddad, G.G.; White, K.P. Shared Genetic Signals of Hypoxia Adaptation in Drosophila and in High-altitude Human Populations. Mol. Biol. Evol. 2016, 33, 501–517. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Liu, X.; Zhang, Y.; Li, Y.; Pan, J.; Wang, D.; Chen, W.; Zheng, Z.; He, X.; Zhao, Q.; Pu, Y. EPAS1 gain-of-function mutation contributes to high-altitude adaptation in Tibetan horses. Mol. Biol. Evol. 2019, 36, 2591–2603. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Tufts, D.M.; Natarajan, C.; Revsbech, I.G.; Projecto-Garcia, J.; Hoffmann, F.G.; Weber, R.E.; Fago, A.; Moriyama, H.; Storz, J.F. Epistasis Constrains Mutational Pathways of Hemoglobin Adaptation in High-altitude Pikas. Mol. Biol. Evol. 2015, 32, 287–298. [Google Scholar] [CrossRef] [Green Version]
- Wang, M.-S.; Li, Y.; Peng, M.-S.; Zhong, L.; Wang, Z.-J.; Li, Q.-Y.; Tu, X.-L.; Dong, Y.; Zhu, C.-L.; Wang, L. Genomic Analyses Reveal Potential Independent Adaptation to High Altitude in Tibetan Chickens. Mol. Biol. Evol. 2015, 32, 1880–1889. [Google Scholar] [CrossRef]
- Yu, L.; Wang, G.-D.; Ruan, J.; Chen, Y.-B.; Yang, C.-P.; Cao, X.; Wu, H.; Liu, Y.-H.; Du, Z.-L.; Wang, X.-P. Genomic Analysis of Snub-nosed Monkeys (Rhinopithecus) Identifies Genes and Processes Related to High-altitude Adaptation. Nat. Genet. 2016, 48, 947–952. [Google Scholar] [CrossRef] [Green Version]
- Qiu, Q.; Zhang, G.; Ma, T.; Qian, W.; Wang, J.; Ye, Z.; Cao, C.; Hu, Q.; Kim, J.; Larkin, D.M. The Yak Genome and Adaptation to Life at High Altitude. Nature genetics 2012, 44, 946–949. [Google Scholar] [CrossRef]
- Ding, D.; Liu, G.; Hou, L.; Gui, W.; Chen, B.; Kang, L. Genetic Variation in PTPN1 Contributes to Metabolic Adaptation to High-Altitude Hypoxia in Tibetan Migratory Locusts. Nat. Commun. 2018, 9, 4991. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Dillon, M.E.; Dudley, R. Surpassing Mt. Everest: Extreme Flight Performance of Alpine Bumble-Bees. Biol. Lett. 2014, 10, 20130922. [Google Scholar] [CrossRef] [Green Version]
- Sun, C.; Huang, J.; Wang, Y.; Zhao, X.; Su, L.; Thomas, G.W.; Zhao, M.; Zhang, X.; Jungreis, I.; Kellis, M. Genus-Wide Characterization of Bumblebee Genomes Provides Insights into their Evolution and Variation in Ecological and Behavioral Traits. Mol. Biol. Evol. 2021, 38, 486–501. [Google Scholar] [CrossRef] [PubMed]
- Meyer-Rochow, V. Eyes and Vision of the Bumblebee: A Brief Review on How Bumblebees Detect and Perceive Flowers. J. Apic. 2019, 34, 107–115. [Google Scholar] [CrossRef]
- Liu, Y.; Jin, H.; Naeem, M.; An, J. Comparative Transcriptome Analysis Reveals Regulatory Genes Involved in Cold Tolerance and Hypoxic Adaptation of High-Altitude Tibetan Bumblebees. Apidologie 2020, 51, 1166–1181. [Google Scholar] [CrossRef]
- Liu, Y.; Zhao, H.; Luo, Q.; Yang, Y.; Zhang, G.; Zhou, Z.; Naeem, M.; An, J. De Novo Transcriptomic and Metabolomic Analyses Reveal the Ecological Adaptation of High-Altitude Bombus pyrosoma. Insects 2020, 11, 631. [Google Scholar] [CrossRef] [PubMed]
- Naeem, M. Distribution Pattern of Chinese Bumblebees and Their Facing Threat Assessment. Ph.D. Thesis, Chinese Academy of Agricultural Sciences Dissertation, Beijing, China, 2019. [Google Scholar]
- Williams, P.; Tang, Y.; Yao, J.; Cameron, S. The Bumblebees of Sichuan (Hymenoptera: Apidae, Bombini). Syst. Biodivers. 2009, 7, 101–189. [Google Scholar] [CrossRef] [Green Version]
- Raina, R.H.; Saini, M.S.; Khan, Z.H. Altitudinal food preference of bumblebee species (Hymenoptera: Apidae) from Indian Himalaya. J. Entomol. Zool. Stud. 2019, 7, 234–237. [Google Scholar]
- Grabherr, M.G.; Haas, B.J.; Yassour, M.; Levin, J.Z.; Thompson, D.A.; Amit, I.; Adiconis, X.; Fan, L.; Raychowdhury, R.; Zeng, Q. Full-length Transcriptome Assembly from RNA-Seq Data without a Reference Genome. Nat. Biotechnol. 2011, 29, 644–652. [Google Scholar] [CrossRef] [Green Version]
- Li, B.; Dewey, C.N. RSEM: Accurate Transcript Quantification from RNA-Seq Data with or without a Reference Genome. BMC Bioinform. 2011, 12, 323. [Google Scholar] [CrossRef] [Green Version]
- Love, M.I.; Huber, W.; Anders, S. Moderated Estimation of Fold Change and Dispersion for RNA-Seq Data with DESeq2. Genome Biol. 2014, 15, 550. [Google Scholar] [CrossRef] [PubMed]
- Xin, L.; Guo, G.Q.; Shen, W.Z. Progress in protein families of gap junction. Chin. J. Pathophysiol. 2007, 23, 1240–1243. [Google Scholar] [CrossRef]
- Ma, W.K.; Liu, Q.P.; Liu, K.; Zhang, L.; Wang, L.C.; Han, Z.; Liu, L.; Li, C.; Sun, C.Y. Hypoxic Preconditioning Increases the Expression of Connexins and the Hemichannel Filtration Function. Prog. Mod. Biomed. 2020, 20, 1420–1424. [Google Scholar] [CrossRef]
- Surholt, B.; Greive, H.; Baal, T.; Bertsch, A. Non-shivering Thermogenesis in Asynchronous Flight Muscles of Bumblebees? Comparative Studies on Males of Bombus terrestris, Xylocopa sulcatipes and Acherontia atropos. Comp. Biochem. Physiol. Part A Physiol. 1990, 97, 493–499. [Google Scholar] [CrossRef]
- Dillon, M.E.; Frazier, M.R.; Dudley, R. Into thin air: Physiology and Evolution of Alpine Insects. Integr. Comp. Biol. 2006, 46, 49–61. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Shilo, B.-Z.; Gabay, L.; Glazer, L.; Reichman-Fried, M.; Wappner, P.; Wilk, R.; Zelzer, E. Branching morphogenesis in the Drosophila tracheal system. In Cold Spring Harbor Symposia on Quantitative Biology; Cold Spring Harbor Laboratory Press: New York, NJ, USA, 1997; pp. 241–247. [Google Scholar]
- Arbetman, M.P.; Gleiser, G.; Morales, C.L.; Williams, P.; Aizen, M.A. Global Decline of Bumblebees is Phylogenetically Structured and Inversely Related to Species Range Size and Pathogen Incidence. Proc. R. Soc. B Biol. Sci. 2017, 284, 20170204. [Google Scholar] [CrossRef] [Green Version]
- Roberts, S.P.; Harrison, J.F.; Dudley, R. Allometry of Kinematics and Energetics in Carpenter Bees (Xylocopa varipuncta) Hovering in Variable-density Gases. J. Exp. Biol. 2004, 207, 993–1004. [Google Scholar] [CrossRef]
- Jarecki, J.; Johnson, E.; Krasnow, M.A. Oxygen Regulation of Airway Branching in Drosophila is Mediated by Branchless FGF. Cell 1999, 99, 211–220. [Google Scholar] [CrossRef] [Green Version]
- Wigglesworth, V. Growth and Regeneration in the Tracheal System of an Insect, Rhodnius prolixus (Hemiptera). J. Cell Sci. 1954, 3, 115–137. [Google Scholar] [CrossRef]
- Henry, J.R.; Harrison, J.F. Plastic and Evolved Responses of Larval Tracheae and Mass to Varying Atmospheric Oxygen Content in Drosophila melanogaster. J. Exp. Biol. 2004, 207, 3559–3567. [Google Scholar] [CrossRef] [Green Version]
- Locke, M. The Co-ordination of Growth in the Tracheal System of Insects. J. Cell Sci. 1958, 3, 373–391. [Google Scholar] [CrossRef]
- LOUDON, C. Tracheal Hypertrophy in Mealworms: Design and Plasticity in Oxygen Supply Systems. J. Exp. Biol. 1989, 147, 217–235. [Google Scholar] [CrossRef]
- Conley, K.E.; Lindstedt, S.L. Energy-Saving Mechanisms in Muscle: The Minimization Strategy. J. Exp. Biol. 2002, 205, 2175–2181. [Google Scholar] [CrossRef] [PubMed]
- Hoback, W.W.; Stanley, D.W. Insects in Hypoxia. J. Insect Physiol. 2001, 47, 533–542. [Google Scholar] [CrossRef] [PubMed]
- Ge, R.-L.; Cai, Q.; Shen, Y.-Y.; San, A.; Ma, L.; Zhang, Y.; Yi, X.; Chen, Y.; Yang, L.; Huang, Y. Draft Genome Sequence of the Tibetan Antelope. Nat. Commun. 2013, 4, 1858. [Google Scholar] [CrossRef]




{kind=link}
{kind=link}
{kind=link}
| Sample | Raw Reads | Clean Reads | Clean Bases (G) | Q30 (%) | Mapping Ratio (%) |
|---|---|---|---|---|---|
| KH1 | 49,297,998 | 48,807,124 | 6.68 | 93.53 | 76.12 |
| KH2 | 46,165,938 | 45,665,128 | 6.27 | 93.57 | 69.74 |
| KH3 | 51,256,632 | 50,814,744 | 6.94 | 93.85 | 73.49 |
| KH4 | 52,799,682 | 52,316,554 | 7.12 | 93.75 | 76.73 |
| KH5 | 51,856,992 | 51,368,296 | 7.04 | 93.47 | 74.33 |
| KH6 | 54,244,072 | 53,759,034 | 7.37 | 93.50 | 75.81 |
| KH7 | 44,083,406 | 43,501,182 | 5.96 | 92.98 | 71.97 |
| KM1 | 48,574,042 | 48,087,392 | 6.63 | 93.46 | 77.37 |
| KM2 | 44,543,734 | 44,187,246 | 6.11 | 93.77 | 77.92 |
| KM3 | 45,822,524 | 45,421,310 | 6.28 | 93.66 | 78.59 |
| KM4 | 43,216,750 | 42,745,790 | 5.92 | 93.65 | 74.83 |
| KM5 | 51,397,656 | 50,997,892 | 7.07 | 93.79 | 76.58 |
| KL1 | 46,079,538 | 45,670,082 | 6.32 | 94.07 | 78.60 |
| KL2 | 44,536,062 | 44,243,768 | 6.12 | 93.69 | 78.77 |
| KL3 | 48,138,454 | 47,609,454 | 6.53 | 93.74 | 76.33 |
| KL4 | 53,660,950 | 53,159,194 | 7.27 | 94.01 | 83.32 |
| KL5 | 41,377,256 | 40,918,026 | 5.62 | 93.34 | 74.40 |
| WH1 | 45,484,192 | 45,036,622 | 6.21 | 93.74 | 74.02 |
| WH2 | 43,073,130 | 42,713,976 | 5.92 | 92.14 | 76.93 |
| WH3 | 43,477,462 | 42,996,858 | 5.95 | 93.50 | 76.23 |
| WH4 | 48,568,010 | 48,144,898 | 6.66 | 93.68 | 74.11 |
| WH5 | 52,250,474 | 51,806,312 | 7.17 | 93.71 | 75.00 |
| WM1 | 52,761,382 | 52,233,550 | 7.23 | 93.45 | 75.00 |
| WM2 | 50,270,968 | 49,854,368 | 6.88 | 93.97 | 75.44 |
| WM3 | 51,491,370 | 51,017,398 | 7.07 | 93.78 | 74.25 |
| WL1 | 49,706,148 | 49,026,188 | 6.72 | 93.79 | 77.19 |
| WL2 | 45,457,166 | 44,781,906 | 6.16 | 93.84 | 82.28 |
| WL3 | 63,441,978 | 62,100,090 | 7.74 | 93.45 | 74.63 |
| WL4 | 52,275,552 | 51,754,244 | 7.13 | 93.87 | 75.41 |
| WL5 | 49,883,828 | 49,414,792 | 6.81 | 94.07 | 74.79 |
| 4500 m vs. 4000 m | 5000 m vs. 4500 m | 5000 m vs. 4000 m | |||
|---|---|---|---|---|---|
| Enriched GO term | NES | Enriched GO term | NES | Enriched GO term | NES |
| RNA-directed DNA polymerase activity | −2.78 | G-protein coupled receptor activity | 3.00 | Odorant binding | 2.18 |
| Ligand-gated channel activity | −2.47 | Structural constituent of cuticle | 2.90 | Olfactory receptor activity | 2.16 |
| Ligand-gated ion channel activity | −2.25 | Transmembrane signaling receptor activity | 2.89 | G-protein coupled receptor activity | 2.13 |
| Extracellular ligand-gated ion channel activity | −2.25 | Signaling receptor activity | 2.89 | Transmembrane signaling receptor activity | 2.12 |
| Motile cilium | −2.03 | Ligand-gated ion channel activity | 2.86 | Signaling receptor activity | 2.18 |
| Glutamate receptor activity | −2.00 | Ligand-gated channel activity | 2.82 | Intra-Golgi vesicle-mediated transport | −2.11 |
| Neurotransmitter receptor activity | −1.95 | Extracellular ligand-gated ion channel activity | 2.70 | Protein kinase complex | −2.09 |
| Odorant binding | −1.94 | Extracellular region | 2.65 | Positive regulation of nucleic acid-templated transcription | −2.07 |
| S-methyltransferase activity | 1.92 | Odorant binding | 2.58 | Positive regulation of RNA metabolic process | −2.07 |
| Ion channel complex | −1.91 | Molecular transducer activity | 2.53 | Cyclin-dependent protein kinase holoenzyme complex | −2.07 |
| Structural constituent of cuticle | −1.90 | Neurotransmitter receptor activity | 2.49 | Positive regulation of transcription, DNA-templated | −2.05 |
| Protein lipidation | 1.90 | Olfactory receptor activity | 2.47 | Positive regulation of RNA biosynthetic process | −2.05 |
| G-protein coupled receptor activity | −1.89 | Receptor ligand activity | 2.40 | Endoplasmic reticulum-Golgi intermediate compartment | −2.02 |
| 5-methyltetrahydropteroyltri-L-Glutamate-dependent methyltransferase activity | 1.89 | Hormone activity | 2.40 | Positive regulation of transcription by RNA polymerase II | −2.01 |
| Ionotropic glutamate receptor activity | −1.88 | Signaling receptor binding | 2.39 | Positive regulation of nucleobase-containing compound metabolic process | −2.00 |
| Chitin binding | −1.88 | Cell–cell adhesion | 2.35 | Meiosis I cell cycle process | −1.99 |
| Olfactory receptor activity | −1.87 | Transmitter-gated ion channel activity | 2.35 | Positive regulation of gene expression | −1.98 |
| Transmitter-gated channel activity | −1.87 | Homophilic cell adhesion via plasma membrane adhesion molecules | 2.34 | COPI vesicle coat | −1.98 |
| Steroid dehydrogenase activity | 1.87 | Motile cilium | 2.34 | Acylglycerol O-acyltransferase activity | 1.97 |
| 5-methyltetrahydropteroyltriglutamate-homocysteine S-methyltransferase activity | 1.86 | Transmitter-gated channel activity | 2.33 | Regulation of DNA-templated transcription, elongation | −1.97 |
| 4500 m vs. 4000 m | 5000 m vs. 4000 m | ||
|---|---|---|---|
| GO term | NES | GO term | NES |
| Extracellular region | −2.64 | Regulation of chemotaxis | −3.12 |
| Motile cilium | −2.61 | Regulation of locomotion | −2.55 |
| Structural constituent of cuticle | −2.50 | Response to herbicide | −2.48 |
| Odorant binding | −2.45 | snoRNA binding | −2.36 |
| Cilium | −2.42 | rRNA modification | −2.30 |
| 1-acyl-2-lysophosphatidylserine acylhydrolase activity | −2.32 | Electron transporter, transferring electrons within the cyclic electron transport pathway of photosynthesis activity | −2.37 |
| Fatty-acyl-CoA reductase (alcohol-forming) activity | −2.29 | Odorant binding | −2.11 |
| Alcohol-forming fatty acyl-CoA reductase activity | −2.28 | Regulation of response to external stimulus | −2.07 |
| Phospholipase A1 activity | −2.28 | Cytosolic ribosome | −1.96 |
| Phosphatidylserine 1-acylhydrolase activity | −2.26 | Demethylation | 2.08 |
| Chitin binding | −2.25 | Olfactory receptor activity | −1.90 |
| Signaling receptor activity | −2.25 | ||
| Cell junction | −2.23 | ||
| Cell projection | −2.23 | ||
| Dynein complex | −2.22 | ||
| Transmembrane signaling receptor activity | −2.18 | ||
| Triglyceride lipase activity | −2.16 | ||
| Plasma membrane bounded cell projection | −2.15 | ||
| Cyclic-nucleotide phosphodiesterase activity | −2.06 | ||
| Wnt signaling pathway | −2.06 | ||
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Liang, C.; Liu, D.; Song, P.; Zhou, Y.; Yu, H.; Sun, G.; Ma, X.; Yan, J. Transcriptomic Analyses Suggest the Adaptation of Bumblebees to High Altitudes. Insects 2022, 13, 1173. https://doi.org/10.3390/insects13121173
Liang C, Liu D, Song P, Zhou Y, Yu H, Sun G, Ma X, Yan J. Transcriptomic Analyses Suggest the Adaptation of Bumblebees to High Altitudes. Insects. 2022; 13(12):1173. https://doi.org/10.3390/insects13121173
Chicago/Turabian StyleLiang, Chengbo, Daoxin Liu, Pengfei Song, Yuantao Zhou, Hongyan Yu, Guo Sun, Xiaoxuan Ma, and Jingyan Yan. 2022. "Transcriptomic Analyses Suggest the Adaptation of Bumblebees to High Altitudes" Insects 13, no. 12: 1173. https://doi.org/10.3390/insects13121173