
In Silico and In Vivo Evaluation of Synthesized SCP-2 Inhibiting Compounds on Life Table Parameters of Helicoverpa armigera (Hübner)

 ,
,  , , ,
, , ,  , ,
, ,
Abstract
:Simple Summary
Abstract
1. Introduction
2. Materials and Methods
2.1. Toxicity Predictions
2.2. Molecular Docking
2.3. Molecular Dynamics Simulation
2.4. Larval Diet and Test Concentration Preparation
2.5. Collection and Rearing of Cultures
2.6. Life Table Bioassay
2.7. Statistical Analyses
3. Results
3.1. Toxicity Predictions
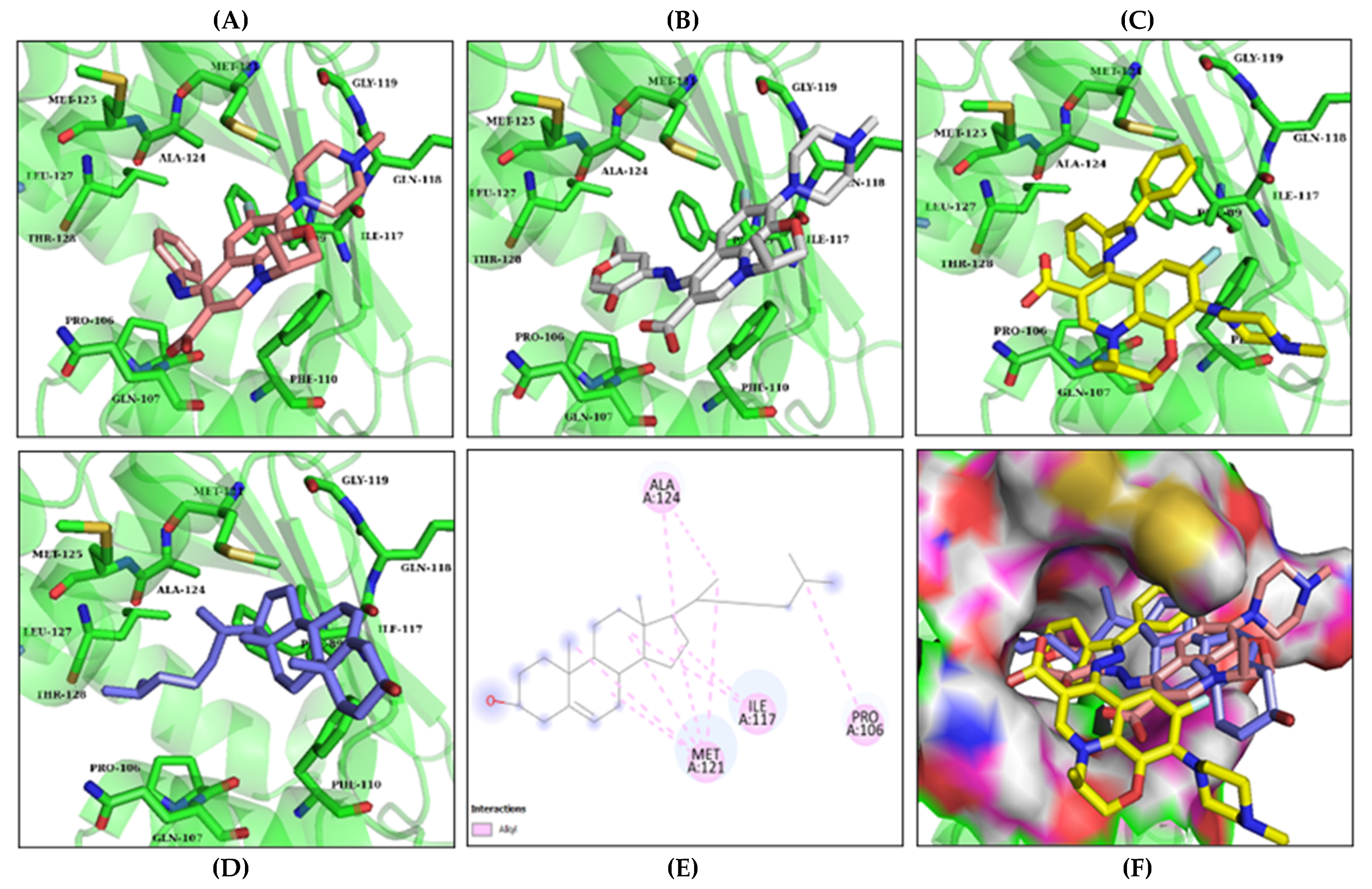
3.2. Molecular Docking
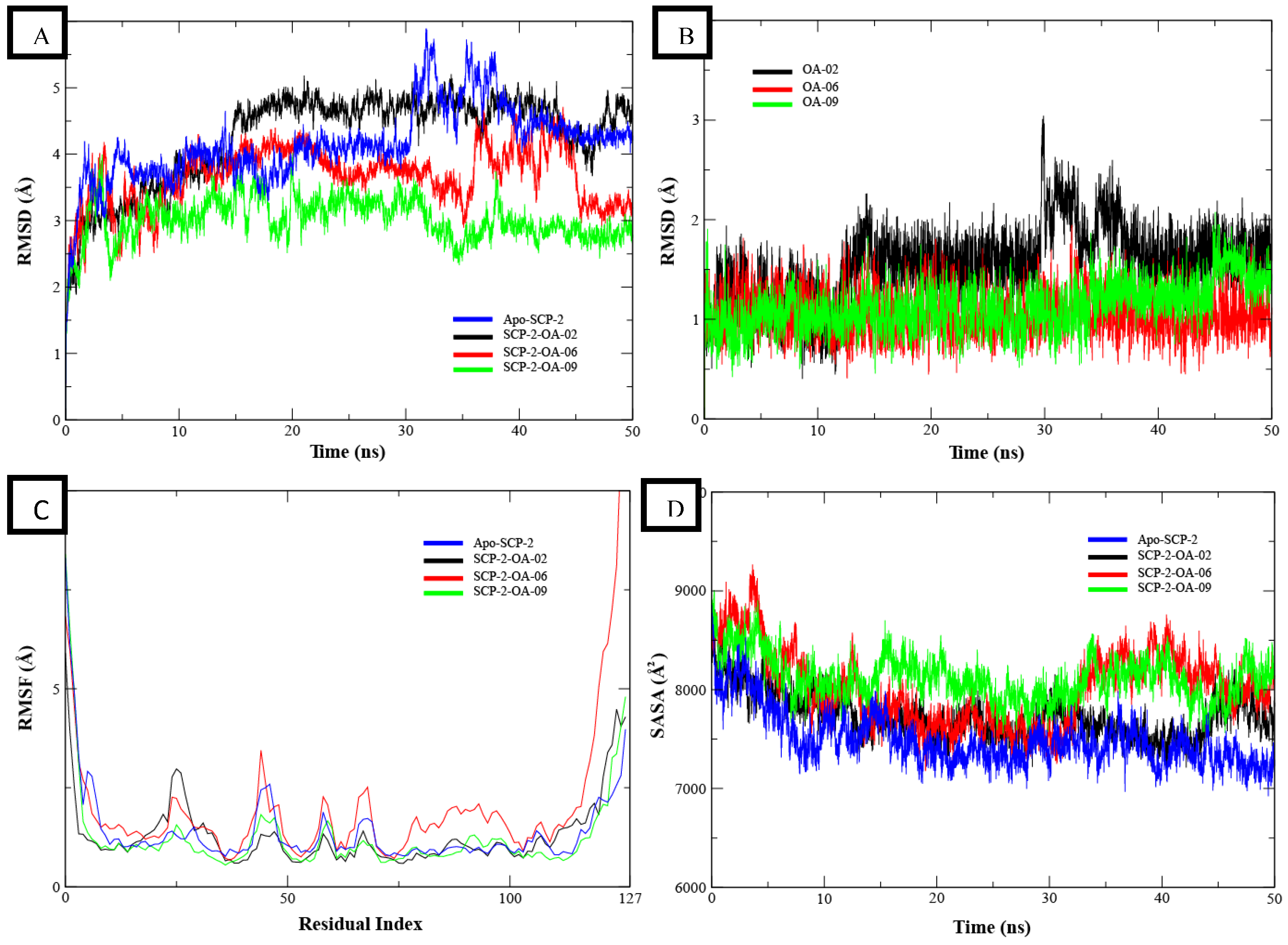
3.3. MD Simulation
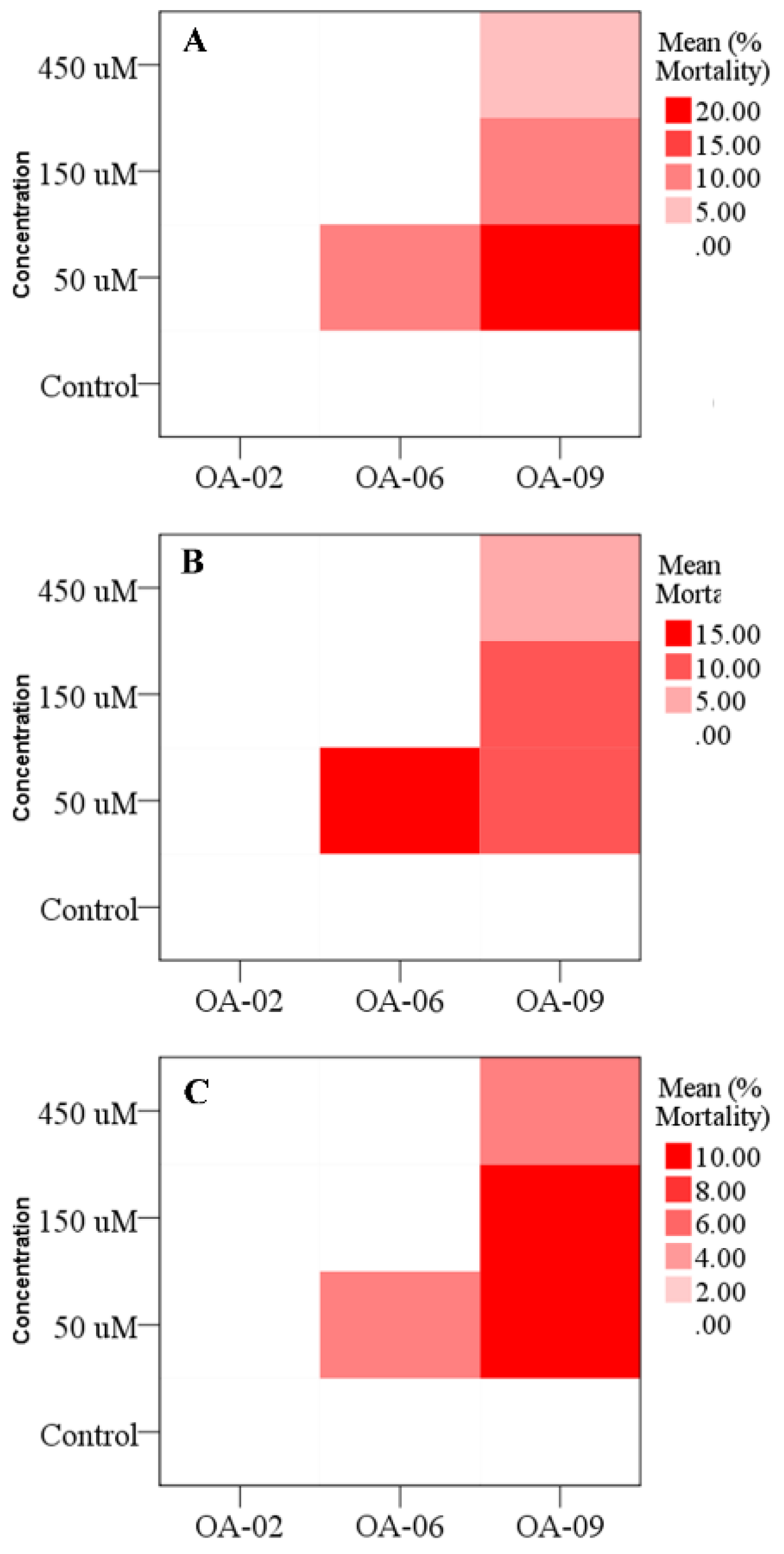
3.4. Larval Mortality
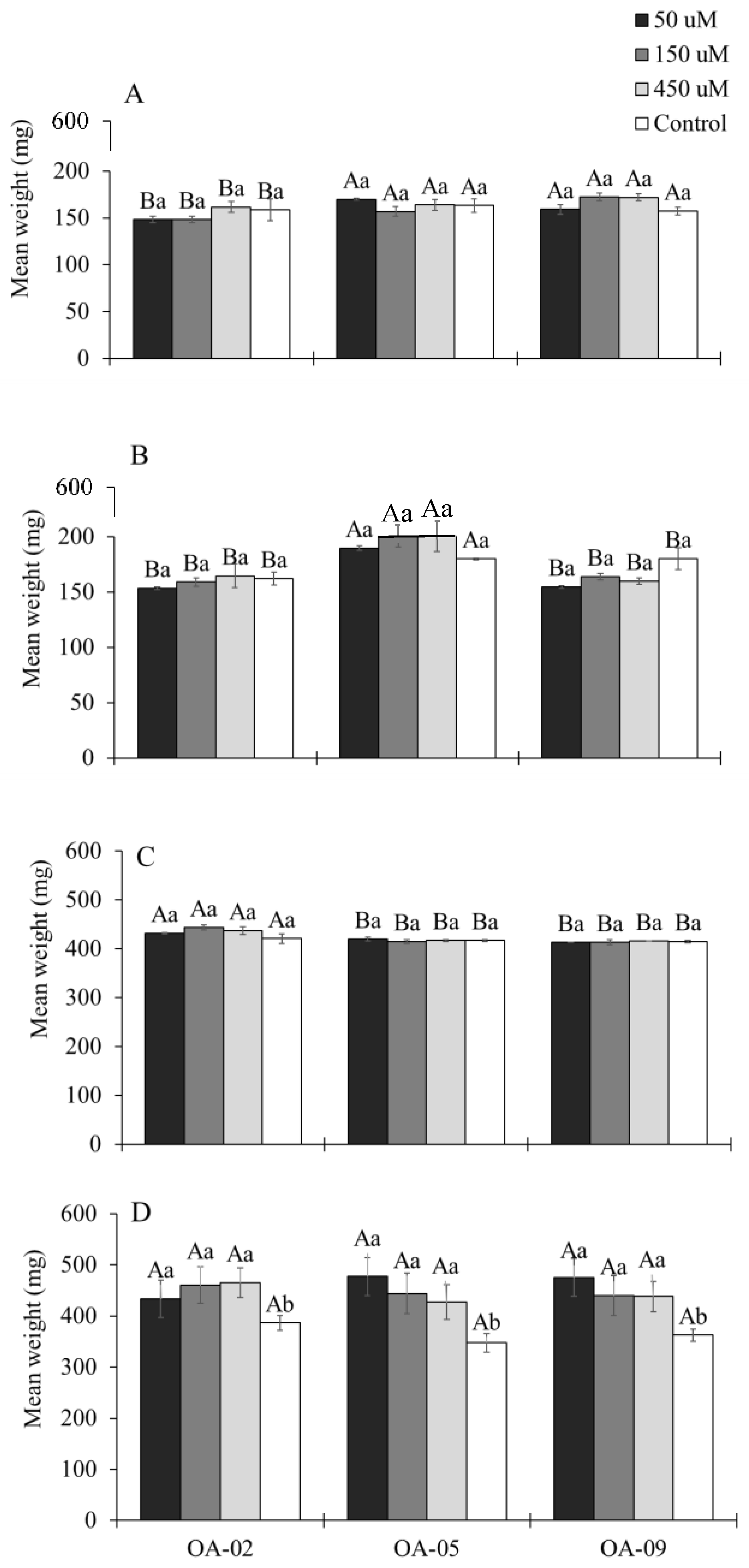
3.5. Growth and Longevity of Different Stages
3.6. Developmental and Reproductive Potentials
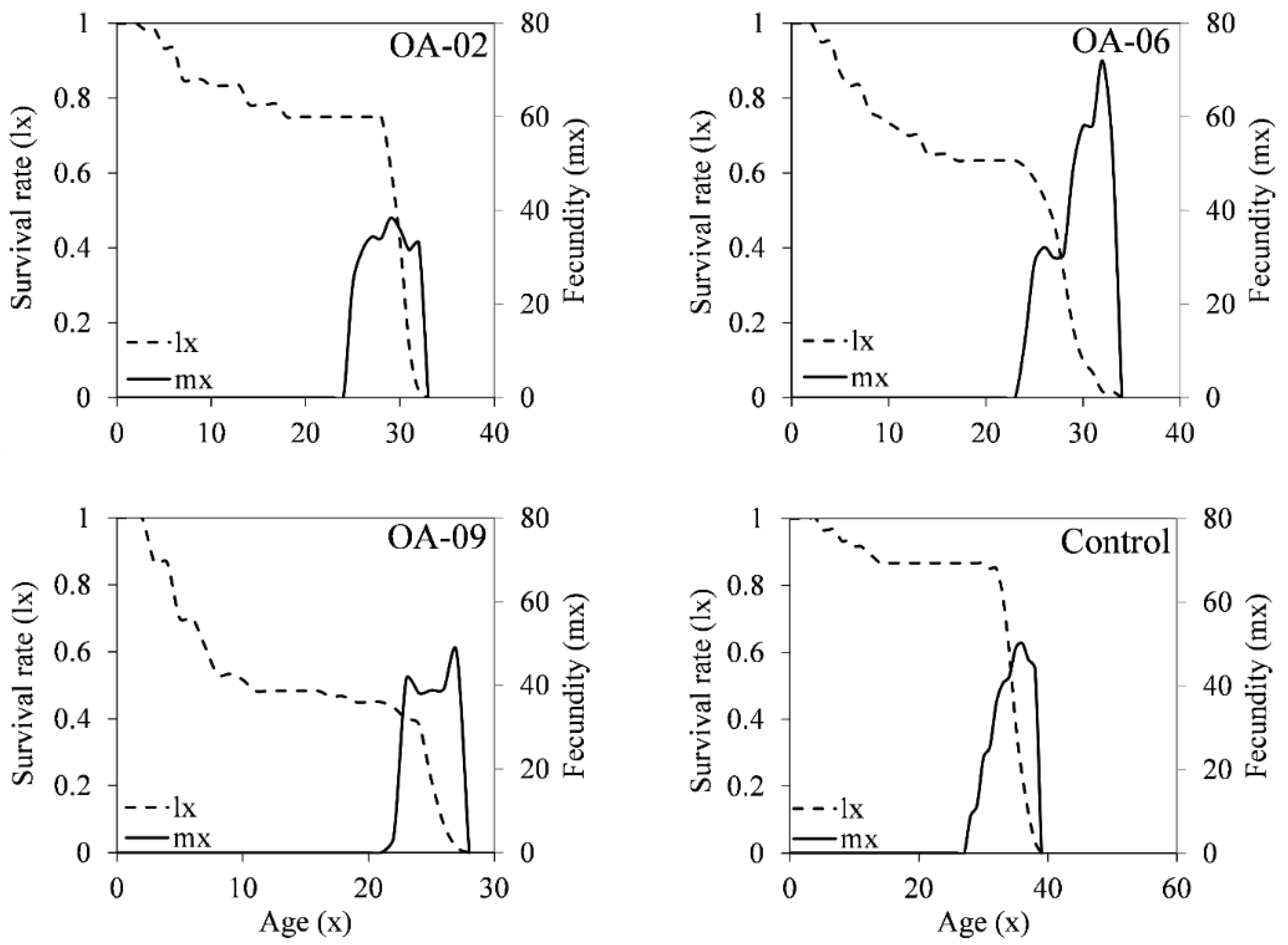
3.7. Lifetable Parameters
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Leong, W.-H.; Teh, S.-Y.; Hossain, M.M.; Nadarajaw, T.; Zabidi-Hussin, Z.; Chin, S.-Y.; Lai, K.-S.; Lim, S.-H.E. Application, monitoring and adverse effects in pesticide use: The importance of reinforcement of Good Agricultural Practices (GAPs). J. Environ. Manag. 2020, 260, 109987. [Google Scholar] [CrossRef] [PubMed]
- Tilman, D. Benefits of intensive agricultural intercropping. Nat. Plants 2020, 6, 604–605. [Google Scholar] [CrossRef] [PubMed]
- Meister, R.T. Farm Chemicals Handbook ’92; Meister Publishing Co.: Willoughby, OH, USA, 1992. [Google Scholar]
- Eddleston, M.; Karalliedde, L.; Buckley, N.; Fernando, R.; Hutchinson, G.; Isbister, G.; Konradsen, F.; Murray, D.; Piola, J.C.; Senanayake, N.; et al. Pesticide poisoning in the developing world—A minimum pesticides list. Lancet 2002, 360, 1163–1167. [Google Scholar] [CrossRef] [PubMed]
- Baker, N.T.; Stone, W.W. Estimated Annual Agricultural Pesticide Use for Counties of the Conterminous United States, 2008–2012; US Geological Survey: Reston, VA, USA, 2015. [Google Scholar]
- Thelin, G.P.; Stone, W.W. Estimation of Annual Agricultural Pesticide Use for Counties of the Conterminous United States, 1992–2009; US Department of the Interior, US Geological Survey: Sacramento, CA, USA, 2013. [Google Scholar]
- Tilman, D.; Fargione, J.; Wolff, B.; D′Antonio, C.; Dobson, A.; Howarth, R.; Schindler, D.; Schlesinger, W.H.; Simberloff, D.; Swackhamer, D. Forecasting Agriculturally Driven Global Environmental Change. Science 2001, 292, 281–284. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Tilman, D.; Balzer, C.; Hill, J.; Befort, B.L. Global food demand and the sustainable intensification of agriculture. Proc. Natl. Acad. Sci. USA 2011, 108, 20260–20264. [Google Scholar] [CrossRef] [Green Version]
- Davydov, R.; Sokolov, M.; Hogland, W.; Glinushkin, A.; Markaryan, A. The application of pesticides and mineral fertilizers in agriculture. In Proceedings of the MATEC Web of Conferences, Chengdu, China, 12–14 January 2018; p. 11003. [Google Scholar]
- Haider, N.; Ahmed, K.S.; Haidary, A.A.; Afzal, M.; Majeed, M.Z. Field evaluation of different insecticides against spotted bollworm (Earias spp.) and comparative yield assessment for BT and non-Bt cotton. J. Entomol. Zool. Stud. 2015, 4, 33–35. [Google Scholar]
- Ma, H.; Meng, C.; Zhang, K.; Wang, K.; Fan, H.; Li, Y. Study on Physiological Mechanism of Using Cottonseed Meal to Improve Salt–Alkali Tolerance of Cotton. J. Plant Growth Regul. 2020, 40, 126–136. [Google Scholar] [CrossRef]
- Vitale, J. Economic Importance of Cotton in Burkina Faso; Food and Agriculture Organisation: Rome, Italy, 2018. [Google Scholar]
- Dhawan, A.K. Integrated pest management in cotton. Pestology 1998, 22, 62–74. [Google Scholar]
- OECD. OECD-FAO Agricultural Outlook 2019–2028; Organisation for Economic Co-Operation and Development (OECD): Paris, France, 2019. [Google Scholar]
- Oerke, E.-C. Crop losses to pests. J. Agric. Sci. 2006, 144, 31–43. [Google Scholar] [CrossRef]
- Behmer, S.T. Overturning dogma: Tolerance of insects to mixed-sterol diets is not universal. Curr. Opin. Insect Sci. 2017, 23, 89–95. [Google Scholar] [CrossRef]
- Tay, W.T.; Soria, M.F.; Walsh, T.; Thomazoni, D.; Silvie, P.; Behere, G.; Anderson, C.; Downes, S. A Brave New World for an Old World Pest: Helicoverpa armigera (Lepidoptera: Noctuidae) in Brazil. PLoS ONE 2013, 8, e80134. [Google Scholar] [CrossRef] [Green Version]
- Mironidis, G.K.; Savopoulou-Soultani, M. Development, survivorship, and reproduction of Helicoverpa armigera (Lepidoptera: Noctuidae) under constant and alternating temperatures. Environ. Entomol. 2014, 37, 16–28. [Google Scholar] [CrossRef]
- Pransopon, P.; Sutantawong, M.; Hormchan, P. Wongpiyasatid. Radiation induced the first filial (F1) sterility in cotton bollworm, Helicoverpa armigera. In Proceedings of the 8th Nuclear Science and Technology Conference, Bangkok, Thailand, 20–21 June 2001. [Google Scholar]
- Haile, F.; Nowatzki, T.; Storer, N. Overview of pest status, potential risk, and management considerations of Helicoverpa armigera (Lepidoptera: Noctuidae) for US soybean production. J. Integr. Pest Manag. 2021, 12, 3. [Google Scholar] [CrossRef]
- Musser, F.; Catchot, A.L., Jr.; Davis, J.; Herbert, D., Jr.; Lorenz, G.; Reed, T.; Reisig, D.; Stewart, S.D. Soybean insect losses in the southern US. Midsouth Entomol. 2012, 6, 12–24. [Google Scholar]
- Michereff-Filho, M.; Fonseca, M.E.N.; Boiteux, L.S.; Torres, J.B.; Silva, K.F.A.D.S.; Specht, A. Helicoverpa armigera Harm 1 Haplotype Predominates in the Heliothinae (Lepidoptera: Noctuidae) Complex Infesting Tomato Crops in Brazil. Neotrop. Entomol. 2021, 50, 258–268. [Google Scholar] [CrossRef]
- Brito, L.O.; Lopes, A.R.; Parra, J.R.P.; Terra, W.R.; Silva-Filho, M.C. Adaptation of tobacco budworm Heliothis virescens to proteinase inhibitors may be mediated by the synthesis of new proteinases. Comp. Biochem. Physiol. Part B Biochem. Mol. Biol. 2001, 128, 365–375. [Google Scholar] [CrossRef]
- Czepak, C.; Albernaz, K.C.; Vivan, L.M.; Guimarães, H.O.; Carvalhais, T. Primeiro registro de ocorrência de Helicoverpa armigera (Hübner) (Lepidoptera: Noctuidae) no Brasil. Pesqui. Agropecuária Trop. 2013, 43, 110–113. [Google Scholar] [CrossRef] [Green Version]
- Arneodo, J.D.; Balbi, E.I.; Flores, F.M.; Sciocco-Cap, A. Molecular identification of Helicoverpa armigera (Lepidoptera: Noctuidae: Heliothinae) in Argentina and development of a novel PCR-RFLP method for its rapid differentiation from H. zea and H. gelotopoeon. J. Econ. Entomol. 2015, 108, 2505–2510. [Google Scholar] [CrossRef]
- Genç, H.; Yücel, S.; Akçal, A. Observation of Helicoverpa armigera Hübner (Lepidoptera: Noctuide) infestation on Gladiolus grandiflorus (Iridaceae) in Çanakkale. COMU J. Agric. Fac. 2017, 5, 105–114. [Google Scholar]
- Murúa, M.G.; Scalora, F.S.; Navarro, F.R.; Cazado, L.E.; Casmuz, A.; Villagrán, M.E.; Lobos, E.; Gastaminza, G. First record of Helicoverpa armigera (Lepidoptera: Noctuidae) in Argentina. Fla. Entomol. 2014, 97, 854–856. [Google Scholar] [CrossRef]
- Gilbert, L.I.; Iatrou, K.; Gill, S.S. Comprehensive Molecular Insect Science; Elsevier: Amsterdam, The Netherlands, 2005. [Google Scholar]
- Hamed, M.; NADEEM, S. Rearing of Helicoverpa armigera (Hub.) on artificial diets in laboratory. Pak. J. Zool. 2008, 40, 447–450. [Google Scholar]
- Zitsanza, E.S.; Giga, D.P.; Knight, J.D. Oviposition site preferences by Helicoverpa armigera (Lepidoptera: Noctuidae): Effect of season and density on distribution of eggs on plants. S. Afr. J. Plant Soil 2006, 23, 138–141. [Google Scholar] [CrossRef]
- Rajapakse, C.N.K.; Walter, G.H. Polyphagy and primary host plants: Oviposition preference versus larval performance in the lepidopteran pest Helicoverpa armigera. Arthropod-Plant Interact. 2007, 1, 17–26. [Google Scholar] [CrossRef]
- Khokhar, S.; Rolania, K.; Singh, G.; Kumar, A. Influence of prevailing weather parameters on population dynamics of fruit borer, Helicoverpa armigera (Hübner) on tomato in Haryana. J. Agrometeorol. 2019, 21, 193–196. [Google Scholar] [CrossRef]
- Khan, M. Adverse Heath Effects, Risk Perception and Pesticie Use Behavior. Munich Personal RePEc Archive. 2009, pp. 3–20. Available online: https://mpra.ub.uni-muenchen.de/16276/ (accessed on 18 October 2022).
- Khan, M.A.; Iqbal, M.; Ahmad, I.; Soomro, M.H. Economic Evaluation of Pesticide Use Externalities in the Cotton Zones of Punjab, Pakistan. Pak. Dev. Rev. 2002, 41, 683–698. [Google Scholar] [CrossRef]
- Ahmad, M.; Arif, M.I.; Attique, M.R. Pyrethroid resistance of Helicoverpa armigera (Lepidoptera: Noctuidae) in Pakistan. Bull. Entomol. Res. 1997, 87, 343–347. [Google Scholar] [CrossRef]
- Ahmad, M.; Arif, M.I.; Ahmad, Z. Resistance to carbamate insecticides in Helicoverpa armigera (Lepidoptera: Noctuidae) in Pakistan. Crop Prot. 2001, 20, 427–432. [Google Scholar] [CrossRef]
- Hussain, D.; Saleem, M.; Ghouse, G.; Abbas, M. Insecticide Resistance in Field Populations of Helicoverpa armigera (Hübner) (Lepidoptera: Noctuidae). J. Entomol. Sci. 2015, 50, 119–128. [Google Scholar]
- Tossou, E.; Tepa-Yotto, G.; Kpindou, O.K.D.; Sandeu, R.; Datinon, B.; Zeukeng, F.; Akoton, R.; Tchigossou, G.M.; Djègbè, I.; Vontas, J.; et al. Susceptibility Profiles of Helicoverpa armigera (Hübner) (Lepidoptera: Noctuidae) to Deltamethrin Reveal a Contrast between the Northern and the Southern Benin. Int. J. Environ. Res. Public Health 2019, 16, 1882. [Google Scholar] [CrossRef] [Green Version]
- Akhurst, R.J.; James, W.; Bird, L.J.; Beard, C. Resistance to the Cry1Ac δ-endotoxin of Bacillus thuringiensis in the cotton bollworm, Helicoverpa armigera (Lepidoptera: Noctuidae). J. Econ. Entomol. 2003, 96, 1290–1299. [Google Scholar] [CrossRef]
- Gao, Y.; Wu, K.; Gould, F.; Shen, Z. Cry2Ab Tolerance Response of Helicoverpa armigera (Lepidoptera: Noctuidae) Populations from Cry1Ac Cotton Planting Region. J. Econ. Entomol. 2009, 102, 1217–1223. [Google Scholar] [CrossRef]
- Saeed, Q.; Saeed, S.; Ahmad, F. Switching among natal and auxiliary hosts increases vulnerability of Spodoptera exigua (Hübner) (Lepidoptera: Noctuidae) to insecticides. Ecol. Evol. 2017, 7, 2725–2734. [Google Scholar] [CrossRef]
- Taleh, M.; Pourabad, R.F.; Geranmaye, J.; Ebadollahi, A. Toxicity of Hexaflumuron as an insect growth regulator (IGR) against Helicoverpa armigera Hubner (Lepidoptera: Noctuidae). J. Entomol. Zool. Stud. 2015, 3, 274–277. [Google Scholar]
- Tunaz, H.; Uygun, N. Insect growth regulators for insect pest control. Turk. J. Agric. For. 2004, 28, 377–387. [Google Scholar]
- Mian, L.S.; Mulla, M.S. Biological and environmental dynamics of insect growth regulators (IGRs) as used against Diptera of public health importance. Residue Rev. 1982, 84, 27–112. [Google Scholar]
- Al-Shannaf, H.; Mead, H.M.; Sabry, A.H. Toxic and biochemical effects of some bioinsecticides and igrs on american bollworm, Helicoverpa armigera (Hüb.) (Noctuidae: Lepidoptera) in cotton fields. J. Biofertil. Biopestic. 2012, 3, 1000118. [Google Scholar]
- Heemers, H.V.; Verhoeven, G.; Swinnen, J. Androgen Activation of the Sterol Regulatory Element-Binding Protein Pathway: Current Insights. Mol. Endocrinol. 2006, 20, 2265–2277. [Google Scholar] [CrossRef]
- Behmer, S.T.; Nes, W.D. Insect Sterol Nutrition and Physiology: A Global Overview. Adv. Insect Physiol. 2003, 31, 1–72. [Google Scholar]
- Lavrynenko, O.; Rodenfels, J.; Carvalho, M.; Dye, N.A.; Lafont, R.; Eaton, S.; Shevchenko, A. The Ecdysteroidome of Drosophila: Influence of diet and development. Development 2015, 142, 3758–3768. [Google Scholar] [CrossRef] [Green Version]
- Douglas, A.E. The microbial dimension in insect nutritional ecology. Funct. Ecol. 2009, 23, 38–47. [Google Scholar] [CrossRef]
- Xue, J.; Zhou, X.; Zhang, C.-X.; Yu, L.; Fan, H.-W.; Wang, Z.; Xu, H.-J.; Xi, Y.; Zhu, Z.-R.; Zhou, W.-W.; et al. Genomes of the rice pest brown planthopper and its endosymbionts reveal complex complementary contributions for host adaptation. Genome Biol. 2014, 15, 1–20. [Google Scholar] [CrossRef] [PubMed]
- Hazarika, P.; McCarty, M.F.; Prieto, V.G.; George, S.; Babu, D.; Koul, D.; Bar-Eli, M.; Duvic, M. Up-regulation of Flotillin-2 Is Associated with Melanoma Progression and Modulates Expression of the Thrombin Receptor Protease Activated Receptor 1. Cancer Res. 2004, 64, 7361–7369. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zheng, J.-C.; Yue, X.-R.; Kuang, W.-Q.; Liu, T.-X.; Jing, X. Genome-wide identification of sterol carrier proteins and their evolutionary diversity in insects. Pak. J. Agric. Sci. 2019, 56, 1065–1074. [Google Scholar]
- Ma, H.; Ma, Y.; Liu, X.; Dyer, D.H.; Xu, P.; Liu, K.; Lan, Q.; Hong, H.; Peng, J.; Peng, R. NMR structure and function of Helicoverpa armigera sterol carrier protein-2, an important insecticidal target from the cotton bollworm. Sci. Rep. 2015, 5, 18186. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Cui, L.; Wang, Q.; Qi, H.; Wang, Q.; Yuan, H.; Rui, C. Resistance selection of indoxacarb in Helicoverpa armigera (Hübner) (Lepidoptera: Noctuidae): Cross-resistance, biochemical mechanisms and associated fitness costs. Pest Manag. Sci. 2018, 74, 2636–2644. [Google Scholar] [CrossRef] [PubMed]
- Khorshidi, M.; Abad, R.F.P.; Saber, M.; Zibaee, A. Effects of hexaflumuron, lufenuron and chlorfluazuron on certain biological and physiological parameters of Helicoverpa armigera (Hübner) (Lepidoptera: Noctuidae). Biocatal. Agric. Biotechnol. 2019, 21, 101270. [Google Scholar] [CrossRef]
- Wang, D.; Qiu, X.; Ren, X.; Zhang, W.; Wang, K. Effects of spinosad on Helicoverpa armigera (Lepidoptera: Noctuidae) from China: Tolerance status, synergism and enzymatic responses. Pest Manag. Sci. Former. Pestic. Sci. 2009, 65, 1040–1046. [Google Scholar] [CrossRef]
- Bueno, A.D.F.; Pomari-Fernandes, A.; Sosa-Gómez, D.R. Helicoverpa armigera: Current status and future perspectives in Brazil. Curr. Agric. Sci. Technol. 2015, 21, 1–7. [Google Scholar]
- Karim, S. Management of Helicoverpa armigera: A Review and Prospectus for Pakistan. Pak. J. Biol. Sci. 2000, 3, 1213–1222. [Google Scholar] [CrossRef] [Green Version]
- Dandan, Z.; Yutao, X.; Wenbo, C.; Yanhui, L.; Kongming, W. Field monitoring of Helicoverpa armigera (Lepidoptera: Noctuidae) Cry1Ac insecticidal protein resistance in China (2005–2017). Pest Manag. Sci. 2018, 75, 753–759. [Google Scholar] [CrossRef]
- De Barros, M.; da Silva, L.M.; Boeing, T.; Somensi, L.B.; Cury, B.J.; Burci, L.D.M.; Santin, J.R.; de Andrade, S.F.; Monache, F.D.; Cechinel-Filho, V. Pharmacological reports about gastroprotective effects of methanolic extract from leaves of Solidago chilensis (Brazilian arnica) and its components quercitrin and afzelin in rodents. Naunyn-Schmiedeberg’s Arch. Pharmacol. 2016, 389, 403–417. [Google Scholar] [CrossRef]
- Schrödinger Release 2022-3: Maestro; Schrödinger, LLC: New York, NY, USA, 2021.
- Shivakumar, D.; Harder, E.; Damm, W.; Friesner, R.A.; Sherman, W. Improving the Prediction of Absolute Solvation Free Energies Using the Next Generation OPLS Force Field. J. Chem. Theory Comput. 2012, 8, 2553–2558. [Google Scholar] [CrossRef]
- Schrödinger Release 2022-3: LigPrep; Schrödinger, LLC: New York, NY, USA, 2021.
- Friesner, R.A.; Banks, J.L.; Murphy, R.B.; Halgren, T.A.; Klicic, J.J.; Mainz, D.T.; Repasky, M.P.; Knoll, E.H.; Shelley, M.; Perry, J.K.; et al. Glide: A new approach for rapid, accurate docking and scoring. 1. Method and assessment of docking accuracy. J. Med. Chem. 2004, 47, 1739–1749. [Google Scholar] [CrossRef]
- Humphrey, W.; Dalke, A.; Schulten, K. VMD: Visual molecular dynamics. J. Mol. Graph. 1996, 14, 33–38. [Google Scholar] [CrossRef]
- Phillips, J.C.; Hardy, D.J.; Maia, J.D.C.; Stone, J.E.; Ribeiro, J.V.; Bernardi, R.C.; Buch, R.; Fiorin, G.; Hénin, J.; Jiang, W.; et al. Scalable molecular dynamics on CPU and GPU architectures with NAMD. J. Chem. Phys. 2020, 153, 044130. [Google Scholar] [CrossRef]
- Case, D.A.; Aktulga, H.M.; Belfon, K.; Ben-Shalom, I.; Brozell, S.R.; Cerutti, D.S.; Cheatham, T.E., III; Cruzeiro, V.W.D.; Darden, T.A.; Duke, R.E.; et al. Amber 2021: Reference Manual; University of California: San Francisco, CA, USA, 2021. [Google Scholar]
- Jorgensen, W.L.; Chandrasekhar, J.; Madura, J.D.; Impey, R.W.; Klein, M.L. Comparison of simple potential functions for simulating liquid water. J. Chem. Phys. 1983, 79, 926–935. [Google Scholar] [CrossRef]
- Weinzierl, R. Molecular Dynamics Simulations of Human FOXO3 Reveal Intrinsically Disordered Regions Spread Spatially by Intramolecular Electrostatic Repulsion. Biomolecules 2021, 11, 856. [Google Scholar] [CrossRef]
- Grant, B.J.; Skjærven, L.; Yao, X.Q. The Bio3D packages for structural bioinformatics. Protein Sci. 2021, 30, 20–30. [Google Scholar] [CrossRef]
- Jia, B.; Liu, Y.; Zhu, Y.C.; Liu, X.; Gao, C.; Shen, J. Inheritance, fitness cost and mechanism of resistance to tebufenozide in Spodoptera exigua (Hübner) (Lepidoptera: Noctuidae). Pest Manag. Sci. Former. Pestic. Sci. 2009, 65, 996–1002. [Google Scholar] [CrossRef]
- Kakimoto, T.; Fujisaki, K.; Miyatake, T. Egg Laying Preference, Larval Dispersion, and Cannibalism in Helicoverpa armigera (Lepidoptera: Noctuidae). Ann. Entomol. Soc. Am. 2003, 96, 793–798. [Google Scholar] [CrossRef]
- Naseri, B.; Fathipour, Y.; Moharramipour, S.; Hosseininaveh, V. Comparative life history and fecundity of Helicoverpa armigera (Hubner) (Lepidoptera: Noctuidae) on different soybean varieties. Entomol. Sci. 2009, 12, 147–154. [Google Scholar] [CrossRef]
- Bouslama, T.; Chaieb, I.; Jerbi-Elayed, M.; Laarif, A. Observations of some biological characteristics of Helicoverpa armigera reared under controlled conditions. Tunis. J. Plant Prot. 2019, 14, 17–27. [Google Scholar]
- Chi, H.S.I.N.; Liu, H.S.I. Two new methods for the study of insect population ecology. Bull. Inst. Zool. Acad. Sin. 1985, 24, 225–240. [Google Scholar]
- Chi, H. TWOSEX-MSChart: A Computer Program for the Age-Stage, Two-Sex Life Table Analysis. 2018. Available online: http://140.120.197.173/Ecology/ (accessed on 18 October 2022).
- Chi, H. Life-Table Analysis Incorporating Both Sexes and Variable Development Rates Among Individuals. Environ. Entomol. 1988, 17, 26–34. [Google Scholar] [CrossRef]
- Efron, B.; Tibshirani, R.J. An Introduction to the Bootstrap; CRC Press: Boca Raton, FL, USA, 1994. [Google Scholar]
- Gore, J.; Adamczyk, J.J., Jr.; Blanco, C.A. Selective feeding of tobacco budworm and bollworm (Lepidoptera: Noctuidae) on meridic diet with different concentrations of Bacillus thuringiensis proteins. J. Econ. Entomol. 2005, 98, 88–94. [Google Scholar] [CrossRef]
- Liu, T.-X.; Stansly, P.A. Lethal and sublethal effects of two insect growth regulators on adult Delphastus catalinae (Coleoptera: Coccinellidae), a predator of whiteflies (Homoptera: Aleyrodidae). Biol. Control. 2004, 30, 298–305. [Google Scholar] [CrossRef]
- Liu, Z.; Li, D.; Gong, P.; Wu, K. Life table studies of the cotton bollworm, Helicoverpa armigera (Hübner) (Lepidoptera: Noctuidae), on different host plants. Environ. Entomol. 2004, 33, 1570–1576. [Google Scholar] [CrossRef]
- Jha, R.K.; Tuan, S.-J.; Chi, H.; Tang, L.-C. Life table and consumption capacity of corn earworm, Helicoverpa armigera, fed asparagus, Asparagus officinalis. J. Insect Sci. 2014, 14, 34. [Google Scholar] [CrossRef] [Green Version]
- Alam, S.; Sharma, M.; Kumar, R.; Das, J.; Rode, S.; Kumar, P.; Prasad, R.; Sharma, A.K. In silico identification of potential phytochemical inhibitors targeting farnesyl diphosphate synthase of cotton bollworm (Helicoverpa armigera). J. Biomol. Struct. Dyn. 2022, 1–10. [Google Scholar] [CrossRef]
- Kumar, R.; Das, J.; Sharma, M.; Kumar, P.; Sharma, A.K. Identifi-cation of Small-Molecule Potential Inhibitor (s) for Helicoverpa armigera Juvenile Hormone Acid-o-Methyl Transferase (HaJHAMT) through Molecular Docking and MD Simulation Approaches. In Proceedings of the 1st International Electronic Conference on Entomology, Online, 1–15 July 2021. [Google Scholar]
- Cai, J.; Du, X.; Wang, C.; Lin, J.; Du, X. Identification of Potential Helicoverpa armigera (Lepidoptera: Noctuidae) Sterol Carrier Protein-2 Inhibitors Through High-Throughput Virtual Screening. J. Econ. Entomol. 2017, 110, 1779–1784. [Google Scholar] [CrossRef]
- Sargsyan, K.; Grauffel, C.; Lim, C. How molecular size impacts RMSD applications in molecular dynamics simulations. J. Chem. Theory Comput. 2017, 13, 1518–1524. [Google Scholar] [CrossRef]
- Martínez, L. Automatic identification of mobile and rigid substructures in molecular dynamics simulations and fractional structural fluctuation analysis. PLoS ONE 2015, 10, e0119264. [Google Scholar] [CrossRef] [Green Version]
- Genheden, S.; Ryde, U. The MM/PBSA and MM/GBSA methods to estimate ligand-binding affinities. Expert Opin. Drug Discov. 2015, 10, 449–461. [Google Scholar] [CrossRef]
- Golizadeh, A.; Abedi, Z. Feeding performance and life table parameters of Khapra Beetle, Trogoderma granarium Everts (Coleoptera: Dermestidae) on various barley cultivars. Bull. Entomol. Res. 2017, 107, 689–698. [Google Scholar] [CrossRef]
- Du, X.; Ma, H.; Zhang, X.; Liu, K.; Peng, J.; Lan, Q.; Hong, H. Characterization of the sterol carrier protein-x/sterol carrier protein-2 gene in the cotton bollworm, Helicoverpa armigera. J. Insect Physiol. 2012, 58, 1413–1423. [Google Scholar] [CrossRef]
- Abbas, N.; Shad, S.A.; Razaq, M. Fitness cost, cross resistance and realized heritability of resistance to imidacloprid in Spodoptera litura (Lepidoptera: Noctuidae). Pestic. Biochem. Physiol. 2012, 103, 181–188. [Google Scholar] [CrossRef]
- Afzal, M.B.S.; Shad, S.A.; Abbas, N.; Ayyaz, M.; Walker, W.B. Cross-resistance, the stability of acetamiprid resistance and its effect on the biological parameters of cotton mealybug, Phenacoccus solenopsis (Homoptera: Pseudococcidae), in Pakistan. Pest Manag. Sci. 2014, 71, 151–158. [Google Scholar] [CrossRef]
- Ullah, I.; Wazir, S.; Abbas, N.; Naeem, M.; Abdullah, K.; Mahmood, Z.; Rashid, M.-U.; Hafez, A.M. Monitoring of field-evolved resistance to flonicamid, neonicotinoid, and conventional insecticides in the Oxycarenus hyalinipennis costa. Environ. Monit. Assess. 2021, 193, 1–9. [Google Scholar] [CrossRef]
- Fathipour, Y.; Baghery, F.; Bagheri, A.; Naseri, B. Development, reproduction and life table parameters of Helicoverpa armigera (Lepidoptera: Noctuidae) on five main host plants. J. Crop Prot. 2020, 9, 551–561. [Google Scholar]
- Kurtuluş, A.; Pehlivan, S.; Achiri, T.D.; Atakan, E. Influence of different diets on some biological parameters of the Mediterranean flour moth, Ephestia kuehniella Zeller (Lepidoptera: Pyralidae). J. Stored Prod. Res. 2019, 85, 101554. [Google Scholar] [CrossRef]
- Saeed, S.; Jaleel, W.; Naqqash, M.N.; Saeed, Q.; Zaka, S.M.; Sarwar, Z.M.; Ishtiaq, M.; Qayyum, M.A.; Sial, M.U.; Batool, M.; et al. Fitness parameters of Plutella xylostella (L.) (Lepidoptera; Plutellidae) at four constant temperatures by using age-stage, two-sex life tables. Saudi J. Biol. Sci. 2019, 26, 1661–1667. [Google Scholar] [CrossRef] [PubMed]
- Afzal, M.; Khalil, M.S.; Raza, A.M.; Aqueel, M.A.; Khalil, H.; Mansoor, M.M. Bitrophic effects of artificial diets of American bollworm (Helicoverpa armigera H.) on different biological aspects of Bracon hebetor Say. J. Saudi Soc. Agric. Sci. 2020, 19, 26–30. [Google Scholar] [CrossRef]
- Zhu, T.; Zhou, Q.; Yang, Z.; Zhang, Y.; Luo, J.; Zhang, X.; Shen, Y.; Jiao, L.; Tocher, D.R.; Jin, M. Dietary cholesterol promotes growth and ecdysone signalling pathway by modulating cholesterol transport in swimming crabs (Portunus trituberculatus). Anim. Nutr. 2022, 10, 249–260. [Google Scholar] [CrossRef] [PubMed]






{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| ID | Mol. Wt. | log P | Tumorigenicity | Reproductive Effect | Irritant | Mutagenicity |
|---|---|---|---|---|---|---|
| OA-01 | 457.50 | 1.31 | None | High | High | High |
| OA-02 | 463.51 | 2.85 | None | None | None | None |
| OA-03 | 509.51 | 2.93 | None | None | None | None |
| OA-04 | 542.40 | 3.57 | None | None | None | None |
| OA-05 | 479.51 | 2.50 | None | None | None | None |
| OA-06 | 499.49 | 0.14 | None | None | None | None |
| OA-07 | 509.53 | 2.43 | Low | None | None | None |
| OA-08 | 523.56 | 2.71 | None | None | None | None |
| OA-09 | 539.60 | 4.04 | None | None | None | None |
OA-01 −5.341 | OA-02 −5.369 | OA-03 −7.666 |
OA-04 −4.505 | OA-05 −4.254 | OA-06 −3.365 |
OA-07 −4.305 | OA-08 −4.587 | OA-09 −5.031 |
| Parameters | OA-02 | OA-06 | OA-09 |
|---|---|---|---|
| n | 60 | 60 | 60 |
| LC50 (95% FL) (mg/L) | 313.2 (194.2–845.5) | 143.9 (80.6–250.6) | 60.9 (21.1–98.6) |
| LC90 (95% FL) (mg/L) | 7248.7 (1843.4–532,962.6) | 3778.2 (1141.1–175,820.4) | 1235.9 (533.5–14,300) |
| Probit model fit | |||
| χ2 | 0.90 | 1.20 | 1.80 |
| df | 1 | 1 | 1 |
| Slope ± SE | 0.93 ± 0.25 | 0.91 ± 0.25 | 1.01 ± 0.26 |
| p | 0.336 | 0.183 | 0.279 |
| Parameters (Days) | Compound | Concentrations | Test Statistics | |||
|---|---|---|---|---|---|---|
| 50 μM | 150 μM | 450 μM | Control | |||
| Larval duration | OA-02 | 13.36 ± 0.05 Aa | 13.42 ± 0.07 Ab | 5.76 ± 0.07 Ac | 15.03 ± 0.53 Aa | F2, 24 = 140.48 p < 0.001 |
| OA-06 | 16.14 ± 0.12 Ba | 7.14 ± 0.27 Bb | 3.93 ± 0.01 Bc | 11.38 ± 2.25 Ba | ||
| OA-09 | 10.00 ± 0.10 Ca | 6.01 ± 0.04 Cb | 3.75 ± 0.02 Cc | 11.25 ± 2.16 Ca | ||
| Pupal duration | OA-02 | 16.19 ± 0.12 Aa | 15.41 ± 0.11 Aa | 17.12 ± 0.03 Aa | 15.55 ± 0.74 Aa | F2, 24 = 11.137 p < 0.001 |
| OA-06 | 14.41 ± 0.15 Ba | 13.39 ± 0.03 Ba | 14.42 ± 0.09 Ba | 15.53 ± 0.83 Ba | ||
| OA-09 | 10.96 ± 0.30 Ca | 12.23 ± 0.06 Ca | 12.95 ± 0.02 Ca | 15.16 ± 0.87 Ca | ||
| 3rd Instar to adult emergence | OA-02 | 30.56 ± 0.16 Aa | 29.84 ± 0.18 Ab | 23.88 ± 0.10 Ac | 28.25 ± 3.25 Aa | F2, 24 = 4.044 p = 0.006 |
| OA-06 | 31.55 ± 0.26 Ba | 21.53 ± 0.24 Bb | 19.36 ± 0.11 Bc | 27.91 ± 2.77 Ba | ||
| OA-09 | 21.96 ± 0.36 Ca | 19.25 ± 0.10 Cb | 17.70 ± 0.04 Cc | 27.41 ± 2.71 Ca | ||
| Longevity of male moths | OA-02 | 8.47 ± 0.18 Ab | 7.79 ± 0.15 Ab | 7.14 ± 0.09 Ac | 9.01 ± 0.05 Aa | F2, 24 = 28.226 p < 0.001 |
| OA-06 | 6.03 ± 0.04 Bb | 5.70 ± 0.05 Bb | 5.32 ± 0.16 Bc | 8.19 ± 0.03 Ba | ||
| OA-09 | 4.62 ± 0.06 Cb | 5.10 ± 0.10 Cb | 5.40 ± 0.20 Cc | 8.06 ± 0.12 Ca | ||
| Longevity of female moths | OA-02 | 8.92 ± 0.03 Ab | 8.68 ± 0.11 Ab | 8.23 ± 0.13 Ac | 10.02 ± 0.05 Aa | F2, 24 = 19.399 p < 0.001 |
| OA-06 | 7.41 ± 0.09 Bb | 7.07 ± 0.08 Bb | 6.26 ± 0.13 Bc | 9.34 ± 0.08 Ba | ||
| OA-09 | 6.03 ± 0.08 Cb | 6.54 ± 0.02 Cb | 5.80 ± 0.15 Cc | 9.10 ± 0.15 Ca | ||
| Parameters | Concentrations | Test Statistics | ||||
|---|---|---|---|---|---|---|
| 50 μM | 150 μM | 450 μM | Control | |||
| Pupation (%) | OA-02 | 88.00 ± 1.15 Ab | 84.39 ± 0.58 Ab | 81.34 ± 0.49 Ab | 95.00 ± 0.57 Aa | F6,24 = 28.926 p < 0.001 |
| OA-06 | 74.93 ± 0.19 Bb | 80.41 ± 0.59 Bb | 80.86 ± 0.70 Bb | 97.66 ± 1.45 Ba | ||
| OA-09 | 79.78 ± 0.28 Bb | 76.33 ± 0.88 Bb | 82.96 ± 0.35 Bb | 94.93 ± 0.17 Ba | ||
| Adult emergence (%) | OA-02 | 84.29 ± 0.41 Ab | 81.40 ± 0.43 Ac | 81.46 ± 1.08 Ad | 94.35 ± 0.54 Aa | F6,24 = 115.577 p < 0.001 |
| OA-06 | 79.96 ± 0.20 Bb | 80.61 ± 0.77 Bc | 74.70 ± 0.35 Bd | 86.33 ± 0.88 Ba | ||
| OA-09 | 61.95 ± 0.21 Cb | 54.75 ± 1.14 Cc | 54.94 ± 0.33 Cd | 90.25 ± 0.49 Ca | ||
| Fecundity (numbers) | OA-02 | 415.00 ± 2.88 Ab | 461.66 ± 4.40 Ab | 405.33 ± 7.12 Ab | 513.71 ± 3.60 Aa | F6,24 = 7.376 p < 0.001 |
| OA-06 | 391.80 ± 1.74 Bb | 394.72 ± 2.84 Bb | 409.09 ± 2.61 Bb | 499.31 ± 25.53 Ba | ||
| OA-09 | 305.66 ± 3.84 Cb | 291.33 ± 1.45 Cb | 279.44 ± 5.05 Cb | 448.31 ± 15.64 Ca | ||
| Fertility (%) | OA-02 | 84.35 ± 1.74 Ab | 73.64 ± 0.95 Ac | 74.83 ± 0.23 Ad | 87.48 ± 1.61 Aa | F6,24 = 11.591 p < 0.001 |
| OA-06 | 72.73 ± 0.41 Bb | 64.69 ± 0.21 Bc | 61.02 ± 0.23 Bd | 82.88 ± 1.88 Ba | ||
| OA-09 | 64.67 ± 0.45 Cb | 64.01 ± 0.73 Cc | 58.82 ± 0.22 Cd | 82.48 ± 0.99 Ca | ||
| Lifetable Indices | Compounds | Test Statistics | |||
|---|---|---|---|---|---|
| OA-02 | OA-06 | OA-09 | Control | ||
| Intrinsic rate of increase (r) | 0.17 ± 0.01 a | 0.16 ± 0.01 a | 0.17 ± 0.01 a | 0.16 ± 0 a | F3, 399996 = 86675.405 p < 0.001 |
| Finite rate of increase (λ) | 1.19 ± 0.01 a | 1.17 ± 0.01 a | 1.18 ± 0.01 a | 1.17 ± 0.01 a | F3, 399996 = 87349.024 p < 0.001 |
| Net reproductive rate (Ro) | 136.43 ± 22.88 b | 86.27 ± 17.12 c | 57.0 ± 13.64 c | 183.48 ± 26.49 a | F3, 399996 = 730371.962 p < 0.001 |
| Gross reproductive rate (GRR) | 262.86 ± 37.93 b | 424.82 ± 75.65 a | 236.36 ± 43.84 b | 376.86 ± 32.79 ab | F3, 399996 = 262973.719 p < 0.001 |
| Mean generation time (T) | 28.32 ± 0.1 b | 27.67 ± 0.38 b | 24.43 ± 0.17 c | 33.14 ± 0.27 a | F3, 399996 = 20563756.674 p < 0.001 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Saeed, Q.; Ahmad, F.; Yousaf, N.; Ali, H.; Tirmazi, S.A.A.S.; Alshammari, A.; Kausar, N.; Ahmed, M.; Imran, M.; Jamshed, M.; et al. In Silico and In Vivo Evaluation of Synthesized SCP-2 Inhibiting Compounds on Life Table Parameters of Helicoverpa armigera (Hübner). Insects 2022, 13, 1169. https://doi.org/10.3390/insects13121169
Saeed Q, Ahmad F, Yousaf N, Ali H, Tirmazi SAAS, Alshammari A, Kausar N, Ahmed M, Imran M, Jamshed M, et al. In Silico and In Vivo Evaluation of Synthesized SCP-2 Inhibiting Compounds on Life Table Parameters of Helicoverpa armigera (Hübner). Insects. 2022; 13(12):1169. https://doi.org/10.3390/insects13121169
Chicago/Turabian StyleSaeed, Qamar, Faheem Ahmad, Numan Yousaf, Haider Ali, Syed Azhar Ali Shah Tirmazi, Abdulrahman Alshammari, Naeema Kausar, Mahmood Ahmed, Muhammad Imran, Muhammad Jamshed, and et al. 2022. "In Silico and In Vivo Evaluation of Synthesized SCP-2 Inhibiting Compounds on Life Table Parameters of Helicoverpa armigera (Hübner)" Insects 13, no. 12: 1169. https://doi.org/10.3390/insects13121169