


Expression of Immunity- and Stress-Related Genes during an Intermolt Period in the Colorado Potato Beetle
 ,
,
Abstract
:Simple Summary
Abstract
1. Introduction
2. Materials and Methods
2.1. Insects and Fungi
2.2. The Infection Procedure and General Experimental Setup
2.3. Sample Preparation and Quantitative PCR (qPCR)
2.4. Statistics
3. Results
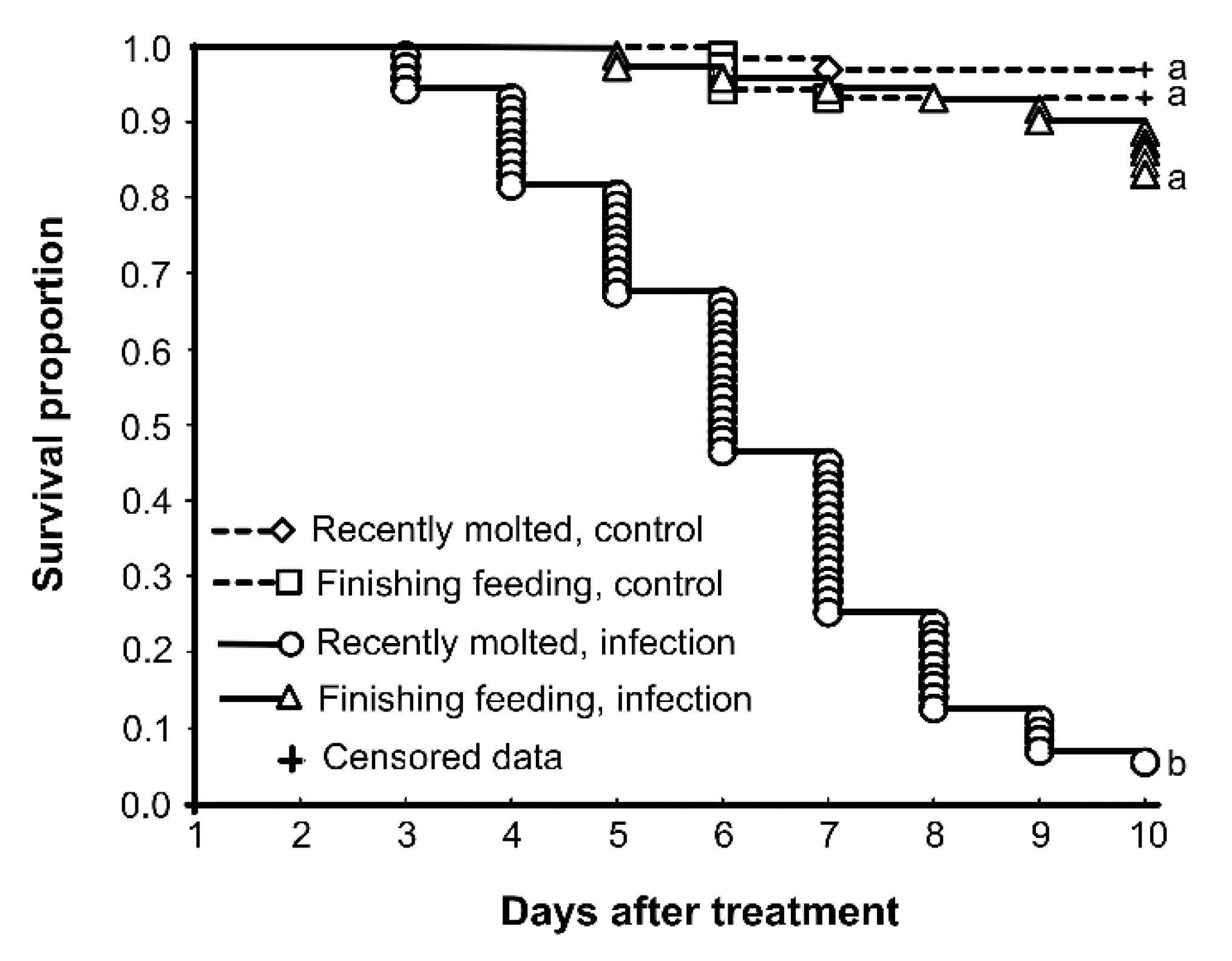
3.1. Susceptibility to M. robertsii
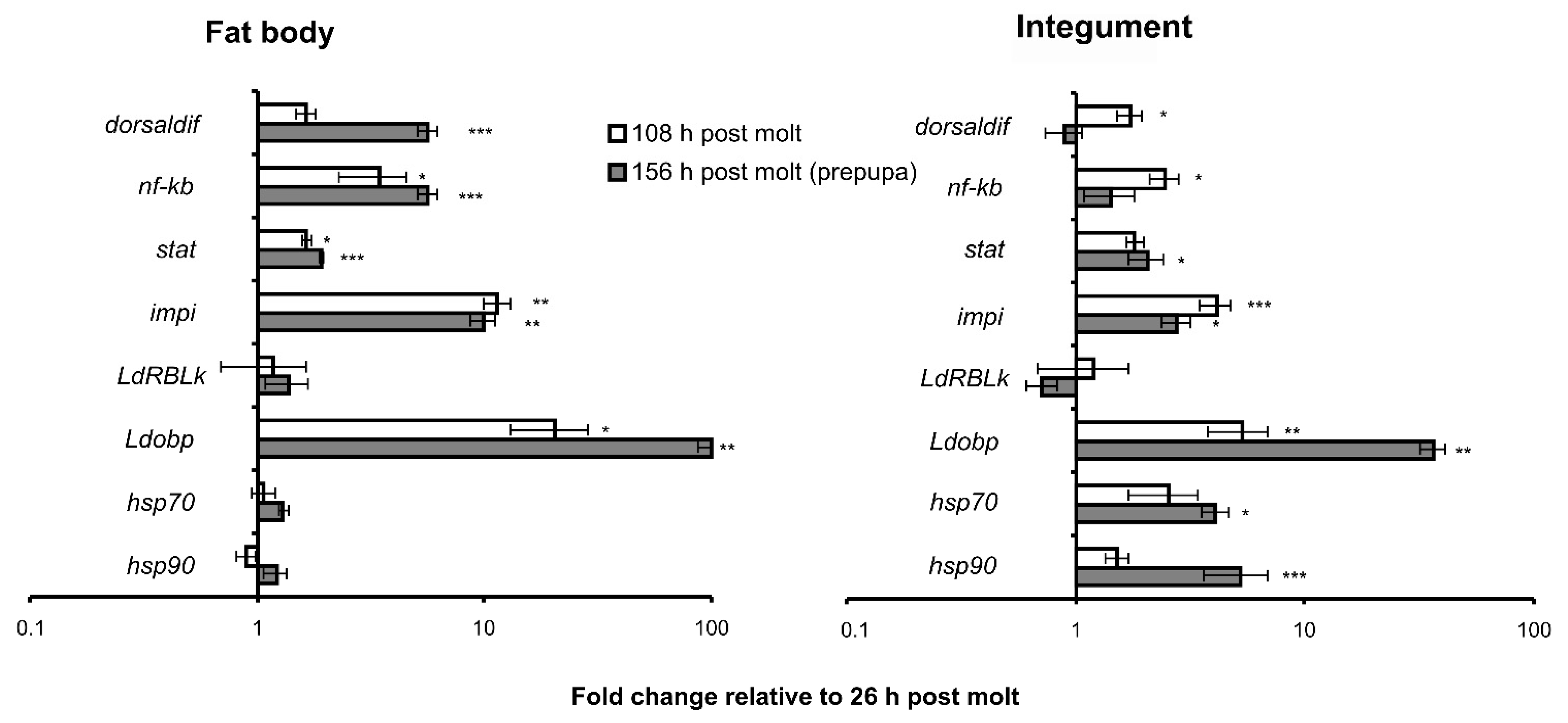
3.2. Changes in Gene Expression during Development
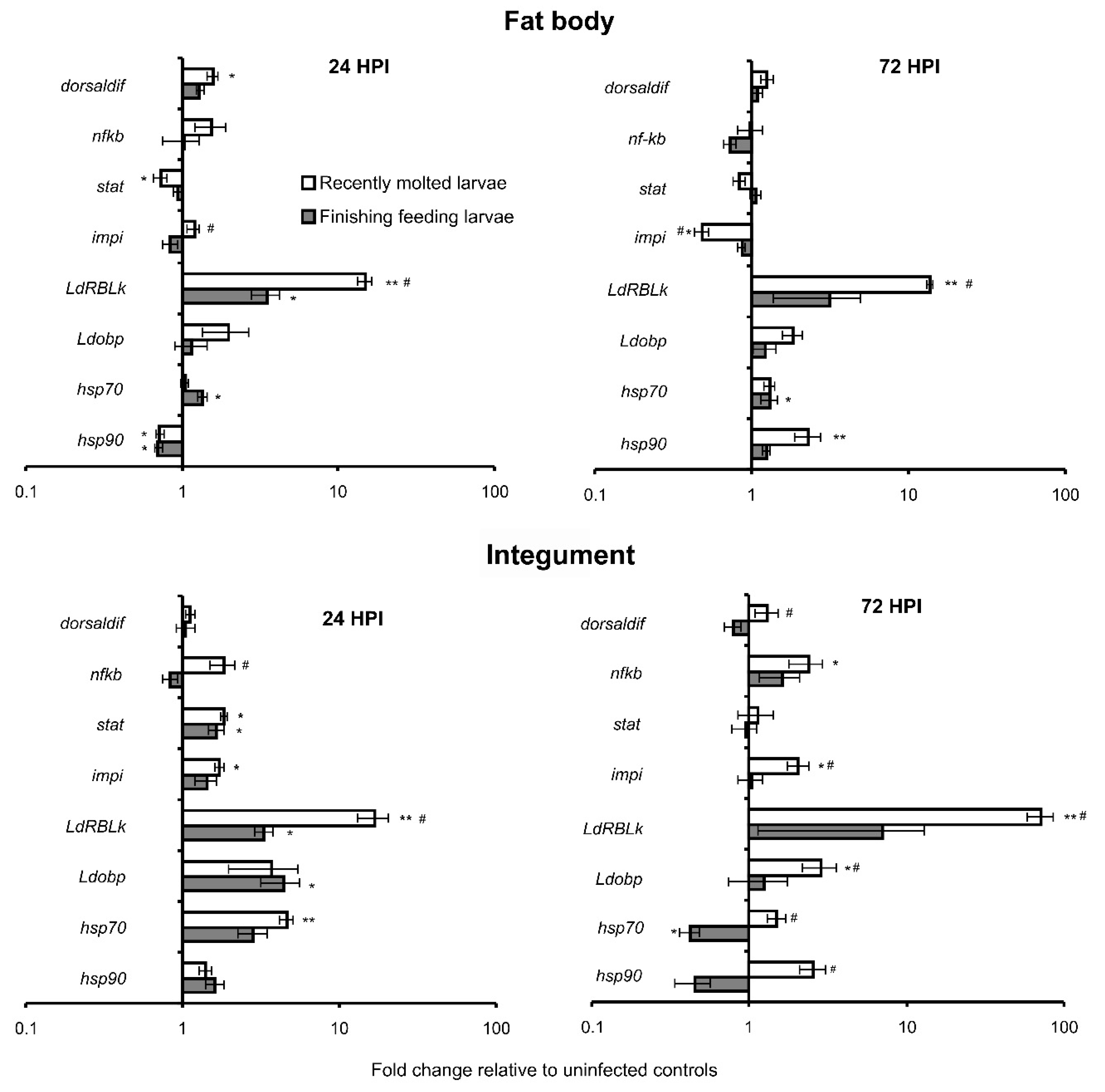
3.3. Alterations of Gene Expression in Response to M. robertsii Infection
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Critchlow, J.T.; Norris, A.; Tate, A.T. The legacy of larval infection on immunological dynamics over metamorphosis. Philos. Trans. R. Soc. B Biol. Sci. 2019, 374, 20190066. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Tate, A.T.; Graham, A.L. Dynamic Patterns of Parasitism and Immunity across Host Development Influence Optimal Strategies of Resource Allocation. Am. Nat. 2015, 186, 495–512. [Google Scholar] [CrossRef] [PubMed]
- Jaronski, S.T. Soil ecology of the entomopathogenic Ascomycetes: A critical examination of what we (think) we know. In Use of Entomopathogenic Fungi in Biological Pest Management; Ekesi, S., Maniania, N.K., Eds.; Research Signpost Inc.: Kerala, India, 2007; pp. 91–143. ISBN 978-81-308-0192-6. [Google Scholar]
- Scheepmaker, J.; Butt, T. Natural and released inoculum levels of entomopathogenic fungal biocontrol agents in soil in relation to risk assessment and in accordance with EU regulations. Biocontrol Sci. Technol. 2010, 20, 503–552. [Google Scholar] [CrossRef]
- Wraight, S.P.; Lacey, L.A.; Kabaluk, J.T.; Goettel, M.S. Potential for microbial biological control of coleopteran and hemipteran pests of potato. In Fruit, Vegetable and Cereal Science and Biotechnology 3 (Special Issue 1); Tennant, P., Benkeblia, N., Eds.; Global Science Book: Ikenobe, Japan, 2009; pp. 25–38. [Google Scholar]
- Weber, D.C.; Blackburn, M.B.; Jaronski, S.T. Biological and behavioral control of potato insect pests. In Insect Pests of Potato, 2nd ed.; Alyokhin, A., Rondon, S.I., Gao, Y., Eds.; Academic Press: Cambridge, MA, USA, 2022; pp. 231–276. [Google Scholar] [CrossRef]
- Alyokhin, A.; Kryukov, V. Ecology of a potato field. In Insect Pests of Potato, 2nd ed.; Alyokhin, A., Rondon, S.I., Gao, Y., Eds.; Academic Press: Cambridge, MA, USA, 2022; pp. 451–462. ISBN 9780128212370. [Google Scholar] [CrossRef]
- Tomilova, O.G.; Yaroslavtseva, O.N.; Ganina, M.D.; Tyurin, M.V.; Chernyak, E.I.; Senderskiy, I.V.; Noskov, Y.A.; Polenogova, O.V.; Akhanaev, Y.B.; Kryukov, V.Y.; et al. Changes in antifungal defence systems during the intermoult period in the Colorado potato beetle. J. Insect Physiol. 2019, 116, 106–117. [Google Scholar] [CrossRef] [PubMed]
- Butt, T.M.; Coates, C.J.; Dubovskiy, I.M.; Ratcliffe, N.A. Entomopathogenic fungi: New insights into host–pathogen interactions. In Advances in Genetics; Lovett, B., St. Leger, R.J., Eds.; Academic Press: Cambridge, MA, USA, 2016; Volume 94, pp. 307–364. ISBN 9780128046944. [Google Scholar]
- Geng, T.; Lv, D.-D.; Huang, Y.-X.; Hou, C.-X.; Qin, G.-X.; Guo, X.-J. JAK/STAT signaling pathway-mediated immune response in silkworm (Bombyx mori) challenged by Beauveria bassiana. Gene 2016, 595, 69–76. [Google Scholar] [CrossRef] [PubMed]
- Ramirez, J.L.; Muturi, E.J.; Barletta, A.B.; Rooney, A.P. The Aedes aegypti IMD pathway is a critical component of the mosquito antifungal immune response. Dev. Comp. Immunol. 2018, 95, 1–9. [Google Scholar] [CrossRef] [PubMed]
- Wei, G.; Lai, Y.; Wang, G.; Chen, H.; Li, F.; Wang, S. Insect pathogenic fungus interacts with the gut microbiota to accelerate mosquito mortality. Proc. Natl. Acad. Sci. USA 2017, 114, 5994–5999. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Vilcinskas, A. Coevolution between pathogen-derived proteinases and proteinase inhibitors of host insects. Virulence 2010, 1, 206–214. [Google Scholar] [CrossRef] [PubMed]
- Rotskaya, U.; Kryukov, V.; Kosman, E.; Tyurin, M.; Glupov, V. Identification of the Ricin-B-Lectin LdRBLk in the Colorado Potato Beetle and an Analysis of Its Expression in Response to Fungal Infections. J. Fungi 2021, 7, 364. [Google Scholar] [CrossRef]
- Scott, I.M.; Hatten, G.; Tuncer, Y.; Clarke, V.C.; Jurcic, K.; Yeung, K.K.-C. Proteomic Analyses Detect Higher Expression of C-Type Lectins in Imidacloprid-Resistant Colorado Potato Beetle Leptinotarsa decemlineata Say. Insects 2020, 12, 3. [Google Scholar] [CrossRef]
- Wei, Z.; Ortiz-Urquiza, A.; Keyhani, N.O. Altered Expression of Chemosensory and Odorant Binding Proteins in Response to Fungal Infection in the Red Imported Fire Ant, Solenopsis invicta. Front. Physiol. 2021, 12. [Google Scholar] [CrossRef] [PubMed]
- Kausar, S.; Abbas, M.N.; Yang, L.; Cui, H. Biotic and abiotic stress induces the expression of Hsp70/90 organizing protein gene in silkworm, Bombyx mori. Int. J. Biol. Macromol. 2020, 143, 610–618. [Google Scholar] [CrossRef] [PubMed]
- King, A.M.; MacRae, T.H. Insect Heat Shock Proteins During Stress and Diapause. Annu. Rev. Èntomol. 2015, 60, 59–75. [Google Scholar] [CrossRef] [PubMed]
- Wrońska, A.K.; Boguś, M.I. Heat shock proteins (HSP 90, 70, 60, and 27) in Galleria mellonella (Lepidoptera) hemolymph are affected by infection with Conidiobolus coronatus (Entomophthorales). PLoS ONE 2020, 15, e0228556. [Google Scholar] [CrossRef] [Green Version]
- Takov, D.; Ostoich, P.; Zubrik, M.; Pilarska, D. Coleopterans as model organisms in insect immunity: A review. North-West. J. Zool. 2022, 18, 104–111. [Google Scholar]
- Shi, X.-Q.; Guo, W.-C.; Wan, P.-J.; Zhou, L.-T.; Ren, X.-L.; Ahmat, T.; Fu, K.-Y.; Li, G.-Q. Validation of reference genes for expression analysis by quantitative real-time PCR in Leptinotarsa decemlineata (Say). BMC Res. Notes 2013, 6, 93. [Google Scholar] [CrossRef] [Green Version]
- National Library of Medicine. BLAST: Basic Local Alignment Search Tool. Available online: https://www.ncbi.nlm.nih.gov/tools/primer-blast/ (accessed on 13 December 2022).
- Integrated DNA Technologies. OligoAnalyzer Tool—Primer Analysis|IDT. Available online: https://eu.idtdna.com/pages/tools/oligoanalyzer (accessed on 13 December 2022).
- Hammer, Ø.; Harper, D.A.T.; Ryan, P.D. PAST: Paleontological statistics software package for education and data analysis. Palaeontol. Electron. 2001, 4, 1–9. [Google Scholar]
- Qu, S.; Wang, S. Interaction of entomopathogenic fungi with the host immune system. Dev. Comp. Immunol. 2018, 83, 96–103. [Google Scholar] [CrossRef]
- Hultmark, D. Drosophila immunity: Paths and patterns. Curr. Opin. Immunol. 2003, 15, 12–19. [Google Scholar] [CrossRef]
- Sorrentino, R.P.; Melk, J.P.; Govind, S. Genetic Analysis of Contributions of Dorsal Group and JAK-Stat92E Pathway Genes to Larval Hemocyte Concentration and the Egg Encapsulation Response in Drosophila. Genetics 2004, 166, 1343–1356. [Google Scholar] [CrossRef]
- Ramirez, J.L.; Dunlap, C.A.; Muturi, E.J.; Barletta, A.B.F.; Rooney, A.P. Entomopathogenic fungal infection leads to temporospatial modulation of the mosquito immune system. PLoS Negl. Trop. Dis. 2018, 12, e0006433. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ramirez, J.L.; Schumacher, M.K.; Ower, G.; Palmquist, D.E.; Juliano, S.A. Impacts of fungal entomopathogens on survival and immune responses of Aedes albopictus and Culex pipiens mosquitoes in the context of native Wolbachia infections. PLoS Negl. Trop. Dis. 2021, 15, e0009984. [Google Scholar] [CrossRef] [PubMed]
- Kryukov, V.Y.; Kosman, E.S. Institute of Systematics and Ecology of Animals, SB RAS, Novosibirsk, Russia. Unpublished work. 2022. Available online: http://reznikova.net/wp-content/uploads/2017/02/Reznikova_Insight2016-1.pdf (accessed on 13 December 2022).
- Tyurin, M.; Kabilov, M.; Smirnova, N.; Tomilova, O.; Yaroslavtseva, O.; Alikina, T.; Glupov, V.; Kryukov, V. Can Potato Plants Be Colonized with the Fungi Metarhizium and Beauveria under Their Natural Load in Agrosystems? Microorganisms 2021, 9, 1373. [Google Scholar] [CrossRef] [PubMed]
- Sikura, A.I.; Sikura, L.V. Entomopathogenous fungi, bacteria, protozoa, viroses. In Colorado Potato Beetle, Leptinotarsa decemlineata Say Phylogeny, Morphology, Physiology, Ecology, Adaptation, Natural Enemies; Nauka Publishers: Moscow, Russia, 1981; pp. 299–313. (In Russian) [Google Scholar]
- Dong, Y.; Morton, J.C.; Ramirez, J.L.; Souza-Neto, J.A.; Dimopoulos, G. The entomopathogenic fungus Beauveria bassiana activate toll and JAK-STAT pathway-controlled effector genes and anti-dengue activity in Aedes aegypti. Insect Biochem. Mol. Biol. 2012, 42, 126–132. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wang, J.-L.; Chen, L.; Tang, L.; Zhao, H.-B.; Liu, X.-S.; Wang, Y.-F. 20-hydroxyecdysone transcriptionally regulates humoral immunity in the fat body of Helicoverpa armigera. Insect Mol. Biol. 2014, 23, 842–856. [Google Scholar] [CrossRef]
- Grizanova, E.V.; Coates, C.J.; Butt, T.M.; Dubovskiy, I.M. RNAi-mediated suppression of insect metalloprotease inhibitor (IMPI) enhances Galleria mellonella susceptibility to fungal infection. Dev. Comp. Immunol. 2021, 122, 104126. [Google Scholar] [CrossRef]
- Zhang, D.; Lu, Y.; Chen, H.; Wu, C.; Zhang, H.; Chen, L.; Chen, X. Antifungal peptides produced by actinomycetes and their biological activities against plant diseases. J. Antibiot. 2020, 73, 265–282. [Google Scholar] [CrossRef]
- Tomilova, O.G.; Kryukov, V.Y.; Duisembekov, B.A.; Yaroslavtseva, O.N.; Tyurin, M.V.; Kryukova, N.A.; Skorokhod, V.; Dubovskiy, I.M.; Glupov, V.V. Immune-physiological aspects of synergy between avermectins and the entomopathogenic fungus Metarhizium robertsii in Colorado potato beetle larvae. J. Invertebr. Pathol. 2016, 140, 8–15. [Google Scholar] [CrossRef]
- EMBL’s European Bioinformatics Institute (EMBL-EBI). Pheromone/general odorant binding protein (IPR006170)—Inter Pro. Available online: https://www.ebi.ac.uk/interpro/entry/InterPro/IPR006170/ (accessed on 13 December 2022).
- Gaubert, A.; Amigues, B.; Spinelli, S.; Cambillau, C. Structure of odorant binding proteins and chemosensory proteins determined by X-ray crystallography. In Methodsin Enzymology; Pelosi, P., Knoll, W., Eds.; Academic Press: Cambridge, MA, USA, 2020; Volume 642, pp. 151–167. ISBN 9780128211571. [Google Scholar]
- Dubovskiy, I.; Whitten, M.; Kryukov, V.; Yaroslavtseva, O.N.; Grizanova, E.; Greig, C.; Mukherjee, K.; Vilcinskas, A.; Mitkovets, P.V.; Glupov, V.V.; et al. More than a colour change: Insect melanism, disease resistance and fecundity. Proc. R. Soc. B Boil. Sci. 2013, 280, 20130584. [Google Scholar] [CrossRef] [Green Version]
- Kryukov, V.Y.; Rotskaya, U.; Yaroslavtseva, O.; Polenogova, O.; Kryukova, N.; Akhanaev, Y.; Krivopalov, A.; Alikina, T.; Vorontsova, Y.L.; Slepneva, I.; et al. Fungus Metarhizium robertsii and neurotoxic insecticide affect gut immunity and microbiota in Colorado potato beetles. Sci. Rep. 2021, 11, 1299. [Google Scholar] [CrossRef]
- Geourjon, C.; Deléage, G. SOPMA: Significant improvement in protein secondary structure prediction by consensus prediction from multiple alignments. Comput. Appl. Biosci. 1995, 11, 681–684. [Google Scholar] [CrossRef] [PubMed]
- Cserzo, M.; Wallin, E.; Simon, I.; von Heijne, G.; Elofsson, A. Prediction of transmembrane alpha-helices in procaryotic membrane proteins: The Dense Alignment Surface method. Protein Eng. 1997, 10, 673–676. [Google Scholar] [CrossRef] [PubMed]




{kind=link}
{kind=link}
{kind=link}
| Gene Name | GenBank Accession Number | Gene Symbol | Primer Sequence (5′→3′) * | Product Size (bp) | PCR Efficiency (± SD) |
|---|---|---|---|---|---|
| 60S ribosomal protein L18 | XM_023172940.1 | rp18 | For TAGAATCCTCAAAGCAGGTGGC Rev CTGGACCAAAGTGTT CACTGC | 133 | 1.98 ± 0.02 |
| ADP-ribosylation factor-like protein 4 | KC190027.1 | arf2 | For GTGCTCGCGAACCATGTGA Rev AAACCTCCAATCCCTCGTGAAG | 140 | 1.94 ± 0.01 |
| ADP-ribosylation factor-like protein 1 | XM_023169879.1 | arf19 | For CGGTGCTGGTAAAACGACAATATT Rev TGACCTCCCAAATCCCAAACTT | 135 | 1.98 ± 0.02 |
| DorsalDif-like protein | XM_023158121.1 | dorsaldif | For TGTGCGAAAAGGTGGCTAAAG Rev ACTTGGGAGGGTTGGAAGTC | 94 | 1.95 ± 0.05 |
| NF-kappaB-like protein | XM_023174540.1 | nfkb | For AAGCAGCGGTTTGATTCGTTC Rev AACTCGTCCAAGTTCTCCAGG | 120 | 1.96 ± 0.06 |
| Signal transducer and activator of transcription protein | XM_023165198.1 | stat | For AGGAGCAGAACACAGGGTAC Rev TTTGCCTGGGAATTCTGTTGAC | 140 | 1.94± 0.03 |
| Inducible metalloproteinase inhibitor protein | XM_023167894.1 | impi | For GCACCCTGTACAAGACCGT Rev CCGTGCGTAACCAGGTATACA | 138 | 1.94 ± 0.02 |
| Ricin-like β-lectin k | GEEF01084863.1 | LdRLBLk | For GAACTGTTGATTGTCGTCACCATG Rev TGGAAAGTTTGGGAGATGGAACT | 140 | 1.96 ± 0.04 |
| Odorant binding protein | XM_023171367.1 | Ldobp | For AGGTGAATCATGAAGTGCATTGC Rev GTGGTACCTCTCTGCTTCCTC | 96 | 1.93 ± 0.02 |
| Heat shock protein 70 | KC544268.1 | hsp70 | For ACCCCGAAGAAGTCAGCTC Rev TCCAGGCCGTAAGCGATG | 214 | 1.96 ± 0.04 |
| Heat shock protein 90 | KC556802.1 | hsp90 | For GGGTGTAGTCGACTCTGAAGAC Rev AGAGCTCCTCAAACAGTTCCAA | 125 | 1.89 ± 0.01 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Kryukov, V.Y.; Rotskaya, U.N.; Yaroslavtseva, O.N.; Noskov, Y.A.; Glupov, V.V. Expression of Immunity- and Stress-Related Genes during an Intermolt Period in the Colorado Potato Beetle. Insects 2022, 13, 1168. https://doi.org/10.3390/insects13121168
Kryukov VY, Rotskaya UN, Yaroslavtseva ON, Noskov YA, Glupov VV. Expression of Immunity- and Stress-Related Genes during an Intermolt Period in the Colorado Potato Beetle. Insects. 2022; 13(12):1168. https://doi.org/10.3390/insects13121168
Chicago/Turabian StyleKryukov, Vadim Yu., Ulyana N. Rotskaya, Olga N. Yaroslavtseva, Yury A. Noskov, and Viktor V. Glupov. 2022. "Expression of Immunity- and Stress-Related Genes during an Intermolt Period in the Colorado Potato Beetle" Insects 13, no. 12: 1168. https://doi.org/10.3390/insects13121168