
Analysis of the Feeding Behavior and Life Table of Nilaparvata lugens and Sogatella furcifera (Hemiptera: Delphacidae) under Sublethal Concentrations of Imidacloprid and Sulfoxaflor
Abstract
:Simple Summary
Abstract
1. Introduction
2. Materials and Methods
2.1. Insects
2.2. Insecticides
2.3. Bioassays
2.4. Sublethal Effects of Two Insecticides on Reproduction
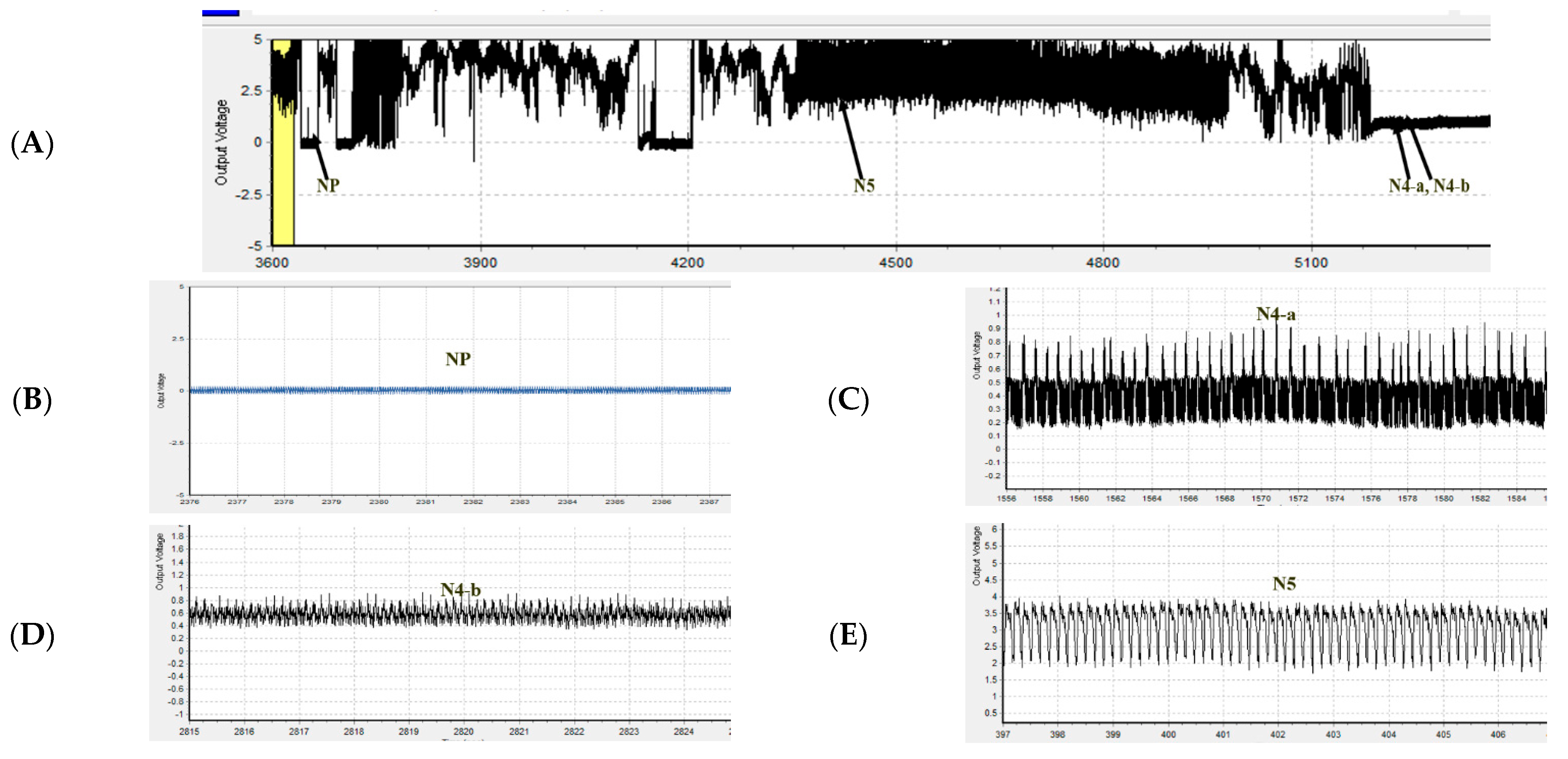
2.5. Electrical Penetration Graph (EPG)
3. Results
3.1. Toxicity of 10 Insecticides to the BPH and WBPH
3.2. Selection of Sublethal Concentrations
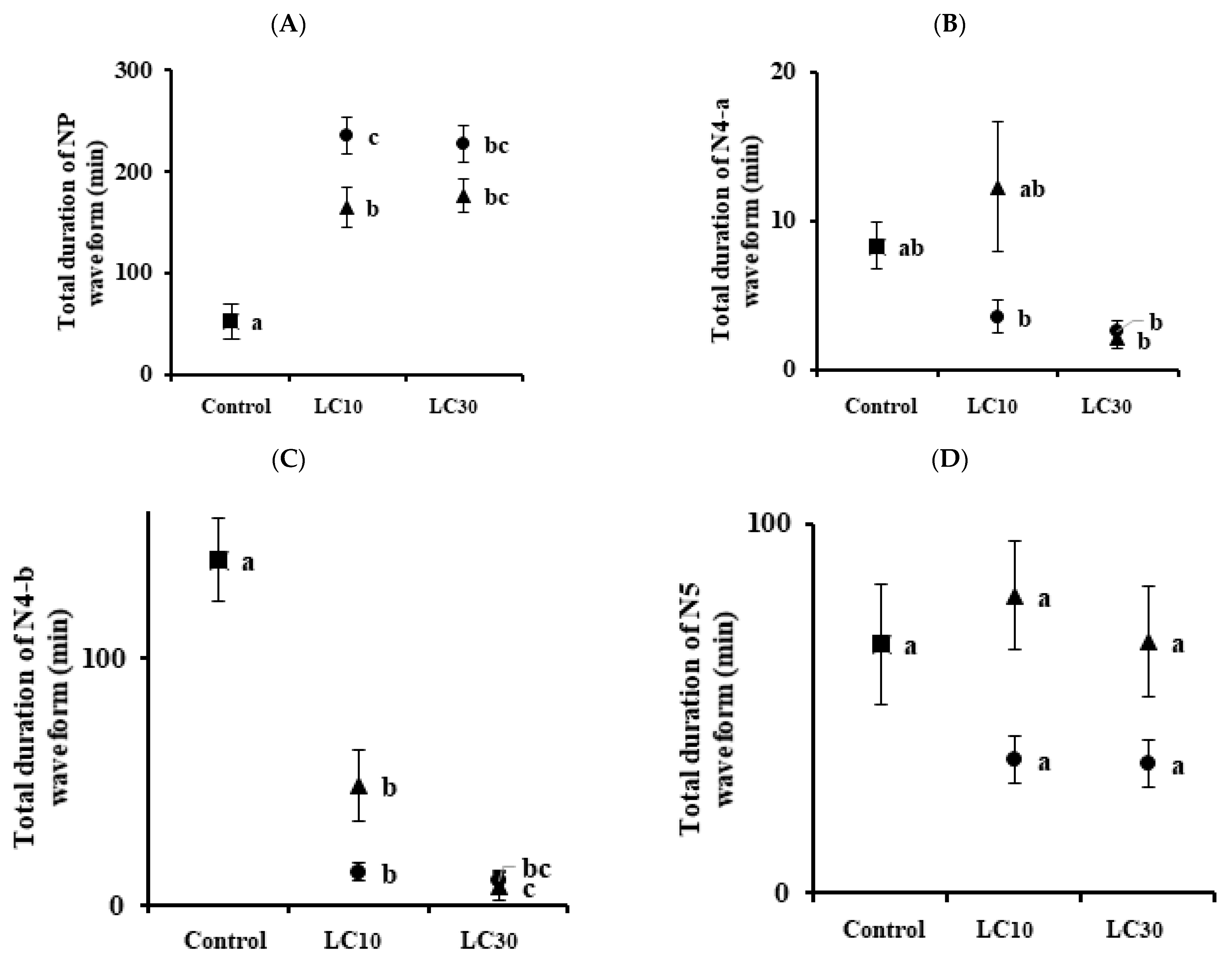
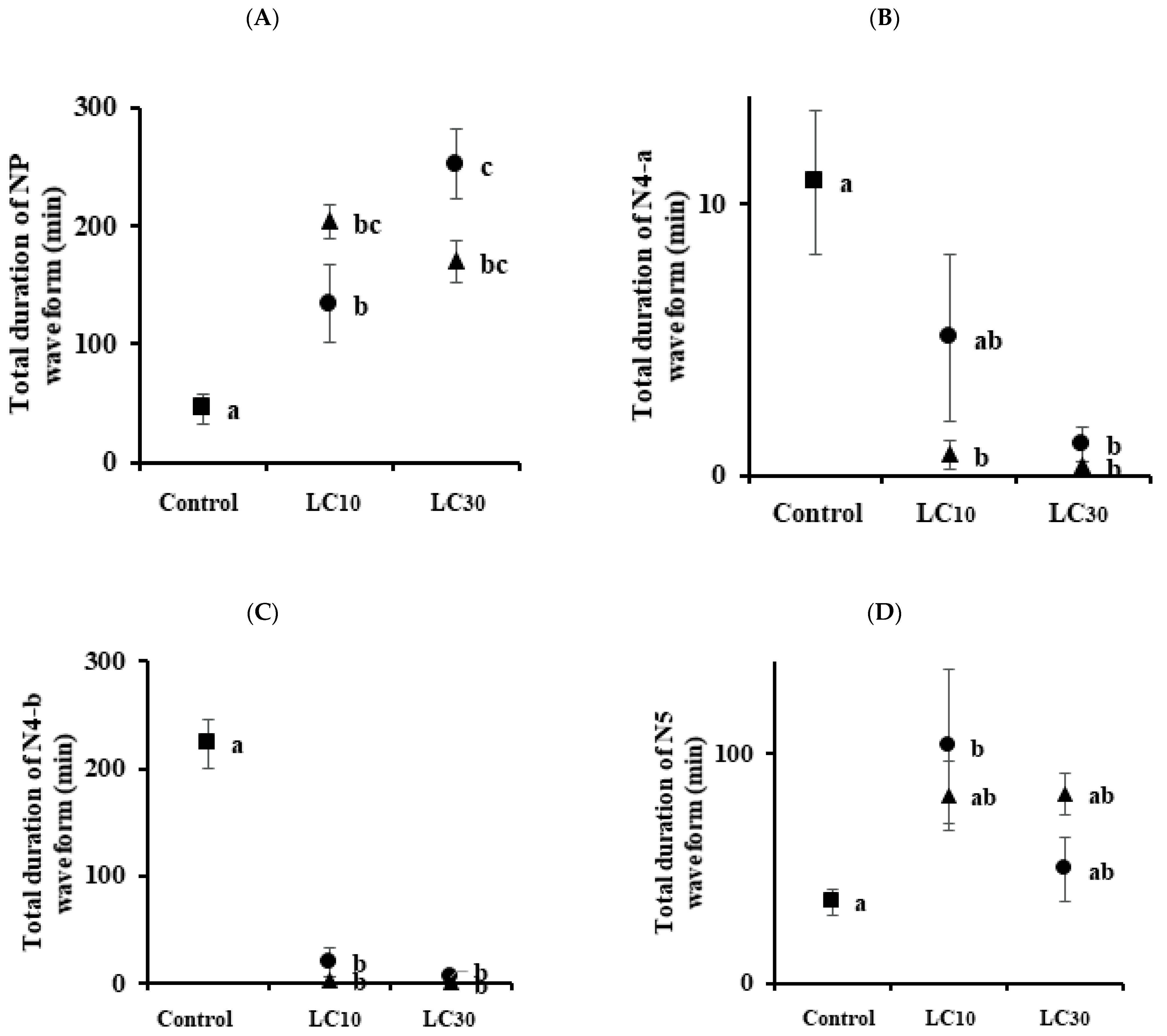
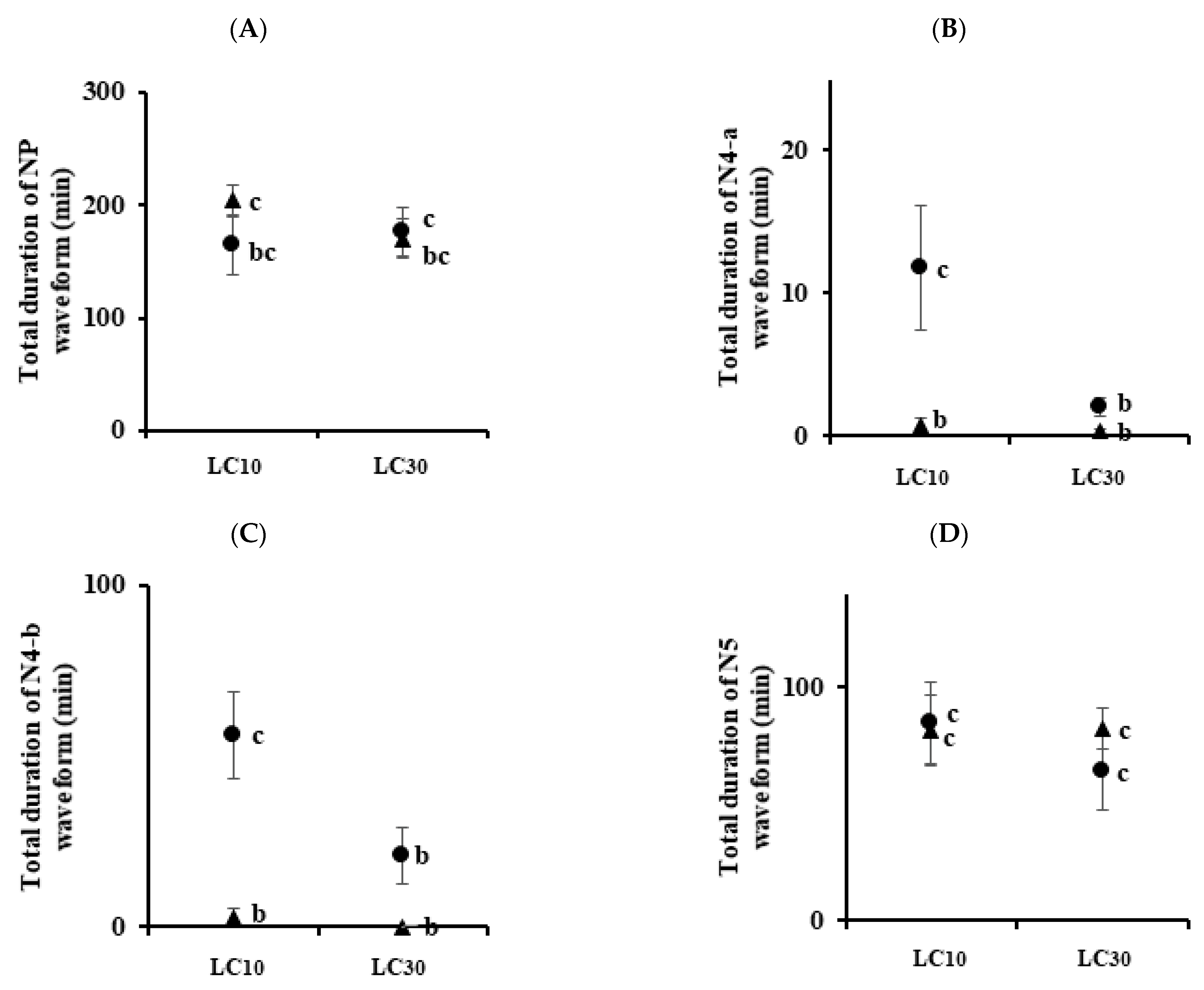
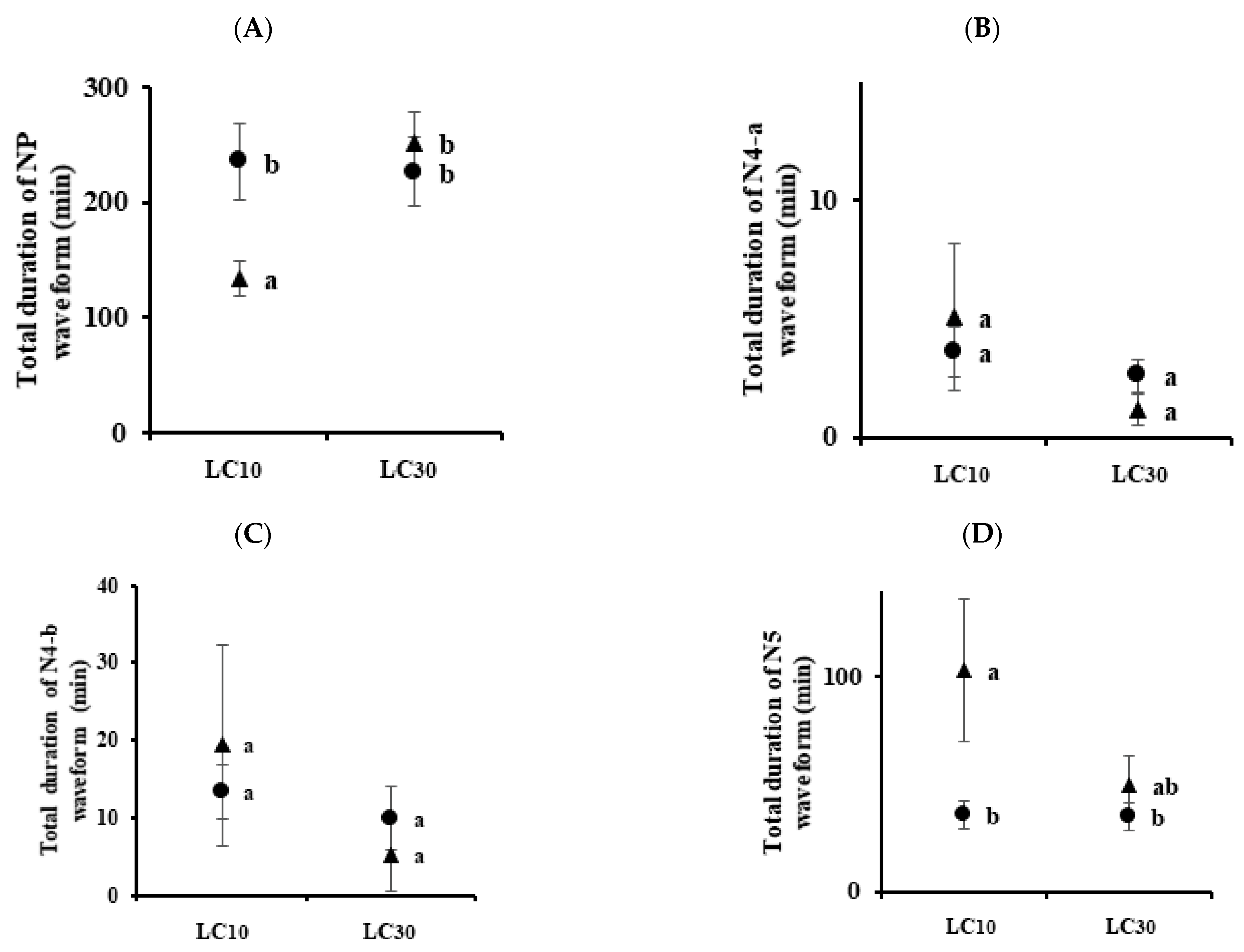
3.3. Analysis of Feeding Behaviors
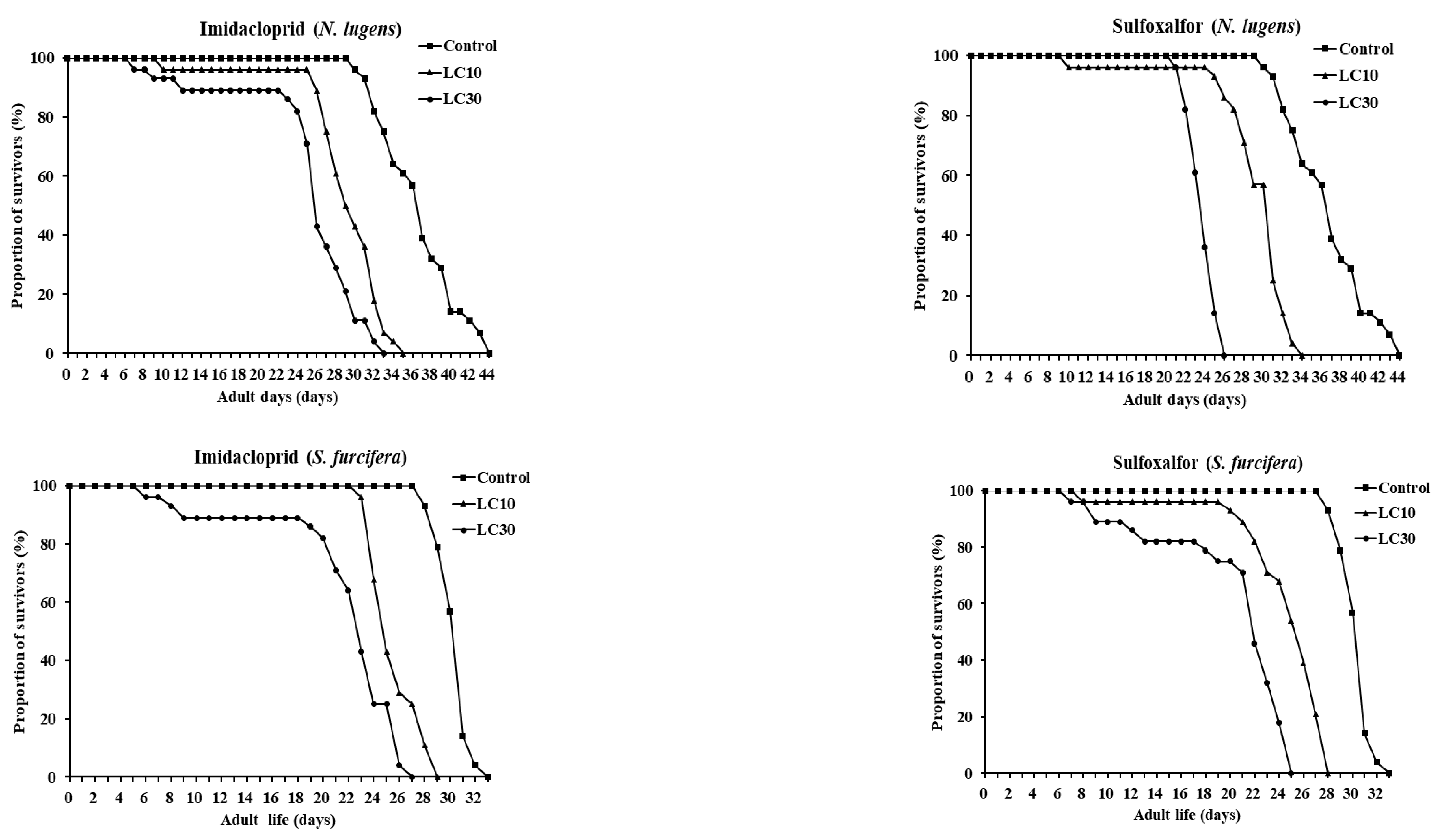
3.4. Influence of Imidacloprid and Sulfoxaflor at Sublethal Concentrations on Lifespan and Fecundity
3.5. Life Table
4. Discussion
5. Conclusions
Author Contributions
Funding
Data Availability Statement
Conflicts of Interest
References
- Yuan, L.-P. Development of hybrid rice to ensure food security. Rice Sci. 2014, 21, 1–2. [Google Scholar] [CrossRef]
- Kisimoto, R. Long-distance migration of rice insects. In Rice Insects: Management Strategies; Heinrichs, E.A., Miller, T.A., Eds.; Springer: New York, NY, USA, 1991; pp. 167–195. [Google Scholar]
- Kisimoto, R.; Sogawa, K. Migration of the brown planthopper Nilaparvata lugens and the white-backed Planthopper Sogatella furcifera in East Asia: The role of weather and climate. In Insect Migration: Tracking Resources through Space and Time; Gatehouse, A.G., Drake, V.A., Eds.; Cambridge University Press: Cambridge, UK, 1995; pp. 67–92. [Google Scholar]
- Otuka, A.; Dudhia, J.; Watanabe, T.; Furuno, A. A new trajectory analysis method for migratory planthoppers, Sogatella furcifera (Horváth) (Homoptera: Delphacidae) and Nilaparvata lugens (Stål), using an advanced weather forecast model. Agric. For. Entomol. 2005, 7, 1–9. [Google Scholar] [CrossRef]
- Lao, S.H.; Huang, X.H.; Huang, H.J.; Liu, C.W.; Zhang, C.X.; Bao, Y.Y. Genomic and transcriptomic insights into the cytochrome P450 monooxygenase gene repertoire in the rice pest brown planthopper, Nilaparvata lugens. Genomics 2015, 106, 301–309. [Google Scholar] [CrossRef]
- Reddy, B.N.; Lakshmi, V.J.; Maheswari, T.; Ramulamma, A.; Katti, G. Studies on antibiosis and tolerance mechanism of resistance to brown planthopper, Nilaparvata lugens (Stal)(Hemiptera: Delphacidae) in the selected rice entries. Ecoscan 2016, 10, 269–275. [Google Scholar]
- Heinrichs, E.A. Impact of insecticides on the resistance and resurgence of rice planthoppers. In Planthoppers: Their Ecology and Management; Denno, R.F., Perfect, T.J., Eds.; Springer: Boston, MA, USA, 1994; pp. 571–598. [Google Scholar]
- Cheng, J.A. Rice planthopper problems and relevant causes in China. In Planthoppers: New Threats to the Sustainability of Intensive Rice Production Systems in Asia; Heong, K.L., Hardy, B., Eds.; International Rice Research Institute: Los Baños, Philippines, 2009; pp. 157–178. [Google Scholar]
- Iamba, K.; Dono, D. A review on brown planthopper (Nilaparvata lugens Stål), a major pest of rice in Asia and Pacific. Asian J. Res. Crop Sci. 2021, 6, 7–19. [Google Scholar] [CrossRef]
- Xu, H.; He, X.; Zheng, X.; Yang, Y.; Tian, J.; Lu, Z. Southern rice black-streaked dwarf virus (SRBSDV) directly affects the feeding and reproduction behavior of its vector, Sogatella furcifera (Horváth)(Hemiptera: Delphacidae). Virol. J. 2014, 11, 1–6. [Google Scholar]
- Cabauatan, P.Q.; Cabunagan, R.C.; Choi, I.R. Rice viruses transmitted by the brown planthopper Nilaparvata lugens Stål. In Planthoppers: New Threats to the Sustainability of Intensive Rice Production Systems in Asia; International Rice Research Institute: Los Baños, Philippines, 2009; pp. 357–368. [Google Scholar]
- Kim, J.D.; Kim, H.J.; Rho, S.P.; Bae, S.H. Analysis of damage on rice by brown planthopper (Nilaparvata lugens Stal) I. Effects of infested stages of rice and appearance days of hopper-burn on yield loss. Korean J. Appl. Entomol. 1984, 23, 153–157. [Google Scholar]
- KCPA. Available online: https://www.koreacpa.org/ko/use-book/ (accessed on 30 April 2022).
- Mullins, J.W. Imidacloprid: A new nitroguanidine insecticide. ACS Symp. Ser. 1993, 524, 183–198. [Google Scholar] [CrossRef]
- Watson, G.B.; Loso, M.R.; Babcock, J.M.; Hasler, J.M.; Letherer, T.J.; Young, C.D.; Zhu, Y.; Casida, J.E.; Sparks, T.C. Novel nicotinic action of the sulfoximine insecticide sulfoxaflor. Insect Biochem. Mol. Biol. 2011, 41, 432–439. [Google Scholar] [CrossRef]
- Desneux, N.; Decourtye, A.; Delpuech, J.-M. The sublethal effects of pesticides on beneficial arthropods. Annu. Rev. Entomol. 2007, 52, 81–106. [Google Scholar] [CrossRef]
- De França, S.M.; Breda, M.O.; Barbosa, D.R.S.; Araujo, A.M.N.; Guedes, C.A. The sublethal effects of insecticides in insects. In Biological Control of Pest and Vector Insects; Vonnie, D.C.S., Ed.; IntechOpen: Rijeka, Croatia, 2017; pp. 23–39. [Google Scholar]
- McLean, D.L.; Kinsey, M.G. Probing behavior of the pea aphid, Acyrthosiphon pisum. I. Definitive correlation of electronically recorded waveforms with aphid probing activities. Ann. Entomol. Soc. Am. 1967, 60, 400–405. [Google Scholar] [CrossRef]
- Tjallingii, W.F. Electronic recording of penetration behaviour by aphids. Entomol. Exp. Appl. 1978, 24, 721–730. [Google Scholar] [CrossRef]
- Pearson, C.C.; Backus, E.A.; Shugart, H.J.; Munyaneza, J.E. Characterization and correlation of EPG waveforms of Bactericera Cockerelli (Hemiptera: Triozidae): Variability in waveform appearance in relation to applied signal. Ann. Entomol. Soc. Am. 2014, 107, 650–666. [Google Scholar] [CrossRef]
- Zhang, Z.; Cui, B.; Zhang, Y. Electrical penetration graphs indicate that tricin is a key secondary metabolite of rice, inhibiting phloem feeding of brown planthopper, Nilaparvata lugens. Entomol. Exp. Appl. 2015, 156, 14–27. [Google Scholar] [CrossRef]
- Hao, Z.P.; Hou, S.M.; Hu, B.C.; Huang, F.; Dang, X.L. Assessment of probing behavior of the cabbage aphid, Brevicoryne brassicae (Hemiptera: Aphididae), on Three Brassica napus cultivars at three developmental stages using electropenetrography (EPG). J. Kans. Entomol. Soc. 2017, 90, 11–23. [Google Scholar] [CrossRef]
- Baldin, E.L.L.; Stamm, M.D.; Bentivenha, J.P.F.; Koch, K.G.; Heng-Moss, T.M.; Hunt, T.E. Feeding Behavior of Aphis glycines (Hemiptera: Aphididae) on Soybeans Exhibiting Antibiosis, Antixenosis, and Tolerance Resistance. Fla. Entomol. 2018, 101, 223–228. [Google Scholar] [CrossRef] [Green Version]
- Chesnais, Q.; Mauck, K.E. Choice of tethering material influences the magnitude and significance of treatment effects in whitefly electrical penetration graph recordings. J. Insect Behav. 2018, 31, 656–671. [Google Scholar] [CrossRef] [Green Version]
- Xu, C.; Lu, C.; Piao, J.; Wang, Y.; Zhou, T.; Zhou, Y.; Li, S. Rice virus release from the planthopper salivary gland is independent of plant tissue recognition by the stylet. Pest Manag. Sci. 2020, 76, 3208–3216. [Google Scholar] [CrossRef]
- Velusamy, R.; Heinrichs, E.A. Electronic monitoring of feeding behavior of Nilaparvata lagens (Homoptera: Delphacidae) on resistant and susceptible rice cultivars. Environ. Entomol. 1986, 15, 678–682. [Google Scholar] [CrossRef]
- Kim, N.; Seo, M.; Youn, Y. Characteristics of feeding behavior of the rice brown planthopper, Nilaparvata lugens, using electrical penetration graph (EPG) technique on different rice varieties. Korean J. Appl. Entomol. 2005, 44, 177–187. [Google Scholar]
- Soffan, A.; Aldawood, A. Electrical penetration graph monitored feeding behavior of cowpea aphid, Aphis craccivora, on faba bean, Vicia faba, cultivars. Turk. Entomol. Derg. 2015, 39, 401–411. [Google Scholar] [CrossRef] [Green Version]
- Ding, X.; Huang, X.; Sun, L.; Wu, J.; Liu, J. Influence of abscisic acid-biosynthesis inhibitor fluridone on the feeding behavior and fecundity of Nilaparvata lugens. Insects 2019, 10, 57. [Google Scholar] [CrossRef] [Green Version]
- SAS Institute. SAS User’s Guide, Statistics Version 9, 1st ed.; SAS Institute: Cary, NC, USA, 2009. [Google Scholar]
- Maia, A.H.N.; Luiz, A.J.B.; Campanhola, C. Statistical influence on associated fertility life table parameters using jackknife technique: Computational aspects. J. Econ. Entomol. 2000, 93, 511–518. [Google Scholar]
- Tjallingii, W.F. Electronic Recordings of Stylet Penetration Activities. Aphids: Their Biology, Natural Enemies and Control; Minks, A.K., Harrewijn, P., Eds.; Elsevier Science Publishers: Amsterdam, The Netherlands, 1988; Volume 2, pp. 95–108. [Google Scholar]
- Tjallingii, W.F.; Hogen Esch, T.H. Fine-structure of aphid stylet routes in plant-tissues in correlation with EPG signals. Physiol. Entomol. 1993, 18, 317328. [Google Scholar] [CrossRef]
- Cho, S.-R.; Koo, H.-N.; Yoon, C.; Kim, G.-H. Sublethal effects of flonicamid and thiamethoxam on green peach aphid, Myzus persicae and feeding behavior analysis. J. Korean Soc. Appl. Biol. Chem. 2011, 54, 889–898. [Google Scholar] [CrossRef]
- Marcic, D.; Petronijevic, S.; Drobnjakovic, T.; Prijovic, M.; Peric, P.; Milenkovic, S. The effects of spirotetramat on life history traits and population growth of Tetranychus urticae (Acari: Tetranychidae). Exp. Appl. Acarol. 2012, 56, 113–122. [Google Scholar] [CrossRef]
- Zhang, R.-M.; Dong, J.-F.; Chen, J.-H.; Ji, Q.-E.; Cui, J.-J. The sublethal effects of chlorantraniliprole on Helicoverpa armigera (Lepidoptera: Noctuidae). J. Integr. Agric. 2013, 12, 457–466. [Google Scholar] [CrossRef]
- Koo, H.-N.; Lee, S.-W.; Yun, S.-H.; Kim, H.K.; Kim, G.-H. Feeding response of the cotton aphid, Aphis gossypii, to sublethal rates of flonicamid and imidacloprid. Entomol. Exp. Appl. 2015, 154, 110–119. [Google Scholar] [CrossRef]
- Chen, X.D.; Ashfaq, M.; Stelinski, L.L. Susceptibility of Asian citrus psyllid, Diaphorina citri (Hemiptera: Liviidae), to the insecticide afidopyropen: A new and potent modulator of insect transient receptor potential channels. Appl. Entomol. Zool. 2018, 53, 453–461. [Google Scholar] [CrossRef] [Green Version]
- Liang, P.-Z.; Ma, K.-S.; Chen, X.-W.; Tang, C.-Y.; Xia, J.; Chi, H.; Gao, X.-W. Toxicity and sublethal effects of flupyradifurone, a novel butenolide insecticide, on the development and fecundity of Aphis gossypii (Hemiptera: Aphididae). J. Econ. Entomol. 2019, 112, 852–858. [Google Scholar] [CrossRef]
- Wang, L.; Wang, Q.; Wang, Q.; Rui, C.; Cui, L. The feeding behavior and life history changes in imidacloprid-resistant Aphis gossypii glover (Homoptera: Aphididae). Pest Manag. Sci. 2020, 76, 1402–1412. [Google Scholar] [CrossRef]
- Butler, C.D.; Walker, G.P.; Trumble, J.T. Feeding disruption of potato psyllid, Bactericera cockerelli, by imidacloprid as measured by electrical penetration graphs. Entomol. Exp. Appl. 2012, 142, 247–257. [Google Scholar] [CrossRef]
- Garzo, E.; Moreno, A.; Hernando, S.; Mariño, V.; Torne, M.; Santamaria, E.; Díaz, I.; Fereres, A. Electrical penetration graph technique as a tool to monitor the early stages of aphid resistance to insecticides. Pest Manag. Sci. 2016, 72, 707–718. [Google Scholar] [CrossRef]
- Boina, D.R.; Onagbola, E.O.; Salyani, M.; Stelinski, L.L. Antifeedant and sublethal effects of imidacloprid on Asian citrus psyllid, Diaphorina citri. Pest Manag. Sci. 2009, 65, 870–877. [Google Scholar] [CrossRef]
- Qin, X.; Zhang, J.; Liu, Q.; Chen, Y.; Zhang, R. Sublethal effects of Paichongding on Nilaparvata lugens (Homoptera: Delphacidae). J. Econ. Entomol. 2013, 106, 10–15. [Google Scholar] [CrossRef]
- Liao, X.; Ali, E.; Li, W.; He, B.; Gong, P.; Xu, P.; Li, J.; Wan, H. Sublethal effects of sulfoxaflor on the development and reproduction of the brown planthopper, Nilaparvata lugens (Stål). Crop Prot. 2019, 118, 6–14. [Google Scholar] [CrossRef]
- Mostafiz, M.M.; Hassan, E.; Shim, J.K.; Lee, K.Y. Lethal and sublethal effects of methyl benzoate on the predatory bug Nesidiocoris tenuis. Insects 2020, 11, 377. [Google Scholar] [CrossRef]







{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Insect | Insecticides | RC (ppm) | n (a) | Mortality (%) | |||
|---|---|---|---|---|---|---|---|
| 24 h | 48 h | 72 h | 96 h | ||||
| Nilaparvata lugens | Imidacloprid (8%, SC) | 16 | 58 | 28.1 ± 14.1 | 39.8 ± 12.9 | 50.6 ± 19.5 | 57.5 ± 8.4 |
| Sulfoxaflor (7%, SC) | 14 | 59 | 62.2 ± 3.8 | 85.8 ± 5.5 | 93.1 ± 11.5 | 93.1 ± 11.5 | |
| Dinotefuran (50%, SG) | 100 | 58 | 90.0 ± 10.0 | 96.6 ± 5.8 | 100 | 100 | |
| Carbosulfan (20%, SC) | 200 | 60 | 47.0 ± 18.5 | 59.8 ± 9.6 | 63.6 ± 14.0 | 63.6 ± 14.0 | |
| Clothianidin (8%, SG) | 16 | 60 | 63.9 ± 21.6 | 66.1 ± 23.6 | 69.3 ± 26.8 | 76.2 ± 15.7 | |
| Etofenprox (10%, EW) | 100 | 59 | 67.8 ± 11.3 | 70.5 ± 5.7 | 70.5 ± 5.7 | 70.5 ± 5.7 | |
| Fenobucarb (50%, EC) | 500 | 62 | 100 | 100 | 100 | 100 | |
| Flonicamid (10%, WG) | 20 | 63 | 10.0 | 17.2 ± 17.3 | 20.7 ± 20.8 | 29.3 ± 7.6 | |
| Pymetrozine (49%, WG) | 98 | 61 | 32.1 ± 10.7 | 56.1 ± 16.5 | 66.5 ± 15.6 | 69.9 ± 10.1 | |
| Thiamethoxam (10% WG) | 10 | 60 | 50.0 ± 10.0 | 48.3 ± 10.0 | 55.2 ± 15.3 | 58.6 ± 10.0 | |
| Sogatella furcifera | Imidacloprid (8%, SC) | 16 | 60 | 56.7 ± 5.8 | 66.7 ± 5.8 | 80.0 | 96.6 ± 5.8 |
| Sulfoxaflor (7%, SC) | 14 | 60 | 40.0 ± 10.0 | 60.0 ± 10.0 | 83.3 ± 5.8 | 100 | |
| Dinotefuran (50%, SG) | 100 | 60 | 93.3 ± 11.5 | 100 | 100 | 100 | |
| Carbosulfan (20%, SC) | 200 | 60 | 90.0 ± 10.0 | 90.0 ± 10.0 | 90.0 ± 10.0 | 89.7 ± 10.0 | |
| Clothianidin (8%, SG) | 16 | 60 | 86.7 ± 5.8 | 93.3 ± 5.8 | 100 | 100 | |
| Etofenprox (10%, EW) | 100 | 60 | 70.0 ± 10.0 | 70.0 ± 10.0 | 70.0 ± 10.0 | 89.7 ± 10.0 | |
| Fenobucarb (50%, EC) | 500 | 60 | 100 | 100 | 100 | 100 | |
| Flonicamid (10%, WG) | 20 | 60 | 6.7 ± 11.5 | 13.3 ± 5.8 | 30.0 ± 10.0 | 44.8 ± 20.8 | |
| Pymetrozine (49%, WG) | 98 | 60 | 13.3 ± 5.8 | 36.7 ± 11.5 | 50.0 ± 17.3 | 65.5 ± 15.3 | |
| Thiamethoxam (10% WG) | 10 | 60 | 30.0 ± 17.3 | 56.7 ± 15.3 | 70.0 ± 30.0 | 96.6 ± 5.8 | |
| Insect | Insecticides | n(a) | Mortality (%) | LC (b)10 | LC30 | Slope ± SE (c) |
|---|---|---|---|---|---|---|
| Nilaparvata lugens | Imidacloprid | 244 | 39.8 | 4.04 (2.73–5.47) | 12.72 (10.01–15.64) | 0.18 ± 0.12 |
| Sulfoxaflor | 240 | 85.8 | 0.83 (0.60–1.08) | 2.30 (1.86–2.75) | 0.10 ± 0.12 | |
| Sogatella furcifera | Imidacloprid | 207 | 66.7 | 1.32 (0.89–1.80) | 3.55 (2.75–4.37) | 0.15 ± 0.14 |
| Sulfoxaflor | 167 | 60.0 | 0.43 (0.14–0.87) | 1.77 (0.87–2.91) | 0.14 ± 0.13 |
| Insect | Insecticides | Conc. | NP | N4-a | N4-b | N5 |
|---|---|---|---|---|---|---|
| Nilaparvata lugens | Control | 9.88 ± 0.97 a (a) | 6.25 ± 1.0 a | 5.25 ± 0.88 a | 2.75 ± 0.53 a | |
| Imidacloprid | LC10 | 11.13 ± 1.01 a | 3.0 ± 0.93 ab | 2.38 ± 0.73 b | 5.38 ± 0.84 a | |
| LC30 | 9.50 ± 1.16 a | 0.88 ± 0.40 b | 0.88 ± 0.40 b | 5.55 ± 0.80 a | ||
| Sulfoxaflor | LC10 | 10.0 ± 1.46 a | 3.0 ± 0.89 ab | 1.38 ± 0.32 b | 4.88 ± 0.95 a | |
| LC30 | 9.38 ± 1.16 a | 2.38 ± 0.68 b | 0.88 ± 0.35 b | 4.75 ± 0.82 a | ||
| Sogatella furcifera | Control | 11.63 ± 1.97 a | 4.88 ± 1.06 a | 2.50 ± 0.57 a | 4.50 ± 0.68 a | |
| Imidacloprid | LC10 | 14.13 ± 1.42 ab | 1.38 ± 0.32 b | 0.63 ± 0.26 b | 9.50 ± 1.52 b | |
| LC30 | 11.25 ± 1.10 a | 0.75 ± 0.31 b | 0.0 b | 7.50 ± 1.24 ab | ||
| Sulfoxaflor | LC10 | 18.63 ± 1.31 b | 5.0 ± 1.02 a | 1.25 ± 0.25 ab | 8.38 ± 1.0 ab | |
| LC30 | 10.88 ± 2.21 a | 2.25 ± 0.62 ab | 0.50 ± 0.19 b | 4.38 ± 1.03 a | ||
| Stage | Nilaparvata lugens | Sogatella furcifera | ||||||||
|---|---|---|---|---|---|---|---|---|---|---|
| Control | Imidacloprid | Sulfoxaflor | Control | Imidacloprid | Sulfoxaflor | |||||
| LC10 | LC30 | LC10 | LC30 | LC10 | LC30 | LC10 | LC30 | |||
| 1st to 5th instar (days) | 20.43 ± 0.39 a (a) | 19.57 ± 0.60 ab | 18.0 ± 0.91 bc | 18.86 ± 0.49 ab | 15.93 ± 0.10 c | 15.82 ± 0.09 a | 15.54 ± 0.11 a | 14.5 ± 0.61 a | 15.04 ± 0.45 a | 14.39 ± 0.59 a |
| Adult longevity (days) | 15.29 ± 0.74 a | 9.0 ± 0.20 b | 6.2 ± 0.41 c | 9.52 ± 0.26 b | 6.96 ± 0.26 c | 13.57 ± 0.19 a | 9.18 ± 0.31 b | 6.67 ± 0.34 c | 8.96 ± 0.34 b | 6.14 ± 0.34 c |
| Total longevity (days) | 35.71 ± 0.74 a | 28.61 ± 0.60 b | 25.64 ± 0.60 c | 28.04 ± 0.59 b | 22.89 ± 0.26 d | 29.39 ± 0.26 a | 24.71 ± 0.34 b | 20.21 ± 1.08 cd | 23.36 ± 0.93 bc | 19.21 ± 1.09 d |
| Pre-oviposition (days) | 5.5 ± 0.14 a | 7.64 ± 0.09 c | 7.54 ± 0.10 c | 7.32 ± 0.09 c | 5.96 ± 0.11 b | 5.82 ± 0.07 a | 5.64 ± 0.09 ab | 6.04 ± 0.13 ab | 6.14 ± 0.07 ab | 6.18 ± 0.07 ab |
| Fecundity (eggs/female) | 129.07 ± 9.83 a | 72.27 ± 3.83 b | 47.93 ± 3.58 cd | 66.0 ± 3.54 bc | 36.67 ± 3.09 d | 102.33 ± 6.11 a | 39.93 ± 2.83 cd | 28.13 ± 1.96 d | 56.07 ± 2.81 b | 49.07 ± 2.61 bc |
| Insect | Insecticides | R0 (a) | rm(b) | T(c) | λ(d) | DT(e) | |
|---|---|---|---|---|---|---|---|
| Nilaparvata lugens | Control | 148.05 ± 10.19 a (f) | 0.182 ± 0.004 a | 27.23 ± 0.30 a | 1.202 ± 0.004 a | 3.80 ± 0.08 a | |
| Imidacloprid | LC10 | 56.98 ± 3.07 b | 0.140 ± 0.007 b | 27.34 ± 0.56 ab | 1.149 ± 0.008 bc | 5.09 ± 0.26 c | |
| LC30 | 38.84 ± 1.60 bc | 0.121 ± 0.005 c | 25.23 ± 0.25 c | 1.129 ± 0.005 c | 5.81 ± 0.20 d | ||
| Sulfoxaflor | LC10 | 46.20 ± 4.96 b | 0.157 ± 0.003 b | 25.94 ± 0.17 bc | 1.167 ± 0.003 b | 4.48 ± 0.09 bc | |
| LC30 | 21.67 ± 2.20 c | 0.148 ± 0.003 b | 20.74 ± 0.20 d | 1.162 ± 0.004 b | 4.12 ± 0.07 b | ||
| Sogatella furcifera | Control | 100.83 ± 5.27 a | 0.19 ± 0.003 a | 24.24 ± 0.18 a | 1.209 ± 0.003 a | 3.65 ± 0.05 a | |
| Imidacloprid | LC10 | 31.67 ± 2.13 bc | 0.151 ± 0.004 c | 22.91 ± 0.24 b | 1.163 ± 0.004 c | 4.65 ± 0.13 c | |
| LC30 | 21.25 ± 1.19 c | 0.145 ± 0.003 c | 21.06 ± 0.23 c | 1.156 ± 0.005 c | 4.82 ± 0.13 c | ||
| Sulfoxaflor | LC10 | 42.15 ± 1.76 b | 0.169 ± 0.003 b | 22.12 ± 0.30 b | 1.185 ± 0.004 b | 4.12 ± 0.08 b | |
| LC30 | 30.66 ± 2.00 c | 0.169 ± 0.005 b | 20.29 ± 0.19 c | 1.183 ± 0.005 b | 4.15 ± 0.10 b | ||
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Kang, Y.; Koo, H.-N.; Kim, H.-K.; Kim, G.-H. Analysis of the Feeding Behavior and Life Table of Nilaparvata lugens and Sogatella furcifera (Hemiptera: Delphacidae) under Sublethal Concentrations of Imidacloprid and Sulfoxaflor. Insects 2022, 13, 1130. https://doi.org/10.3390/insects13121130
Kang Y, Koo H-N, Kim H-K, Kim G-H. Analysis of the Feeding Behavior and Life Table of Nilaparvata lugens and Sogatella furcifera (Hemiptera: Delphacidae) under Sublethal Concentrations of Imidacloprid and Sulfoxaflor. Insects. 2022; 13(12):1130. https://doi.org/10.3390/insects13121130
Chicago/Turabian StyleKang, Yeolgyu, Hyun-Na Koo, Hyun-Kyung Kim, and Gil-Hah Kim. 2022. "Analysis of the Feeding Behavior and Life Table of Nilaparvata lugens and Sogatella furcifera (Hemiptera: Delphacidae) under Sublethal Concentrations of Imidacloprid and Sulfoxaflor" Insects 13, no. 12: 1130. https://doi.org/10.3390/insects13121130