

Tangled in a Web: Management Type and Vegetation Shape the Occurrence of Web-Building Spiders in Protected Areas

Abstract
:Simple Summary
Abstract
1. Introduction
2. Materials and Methods
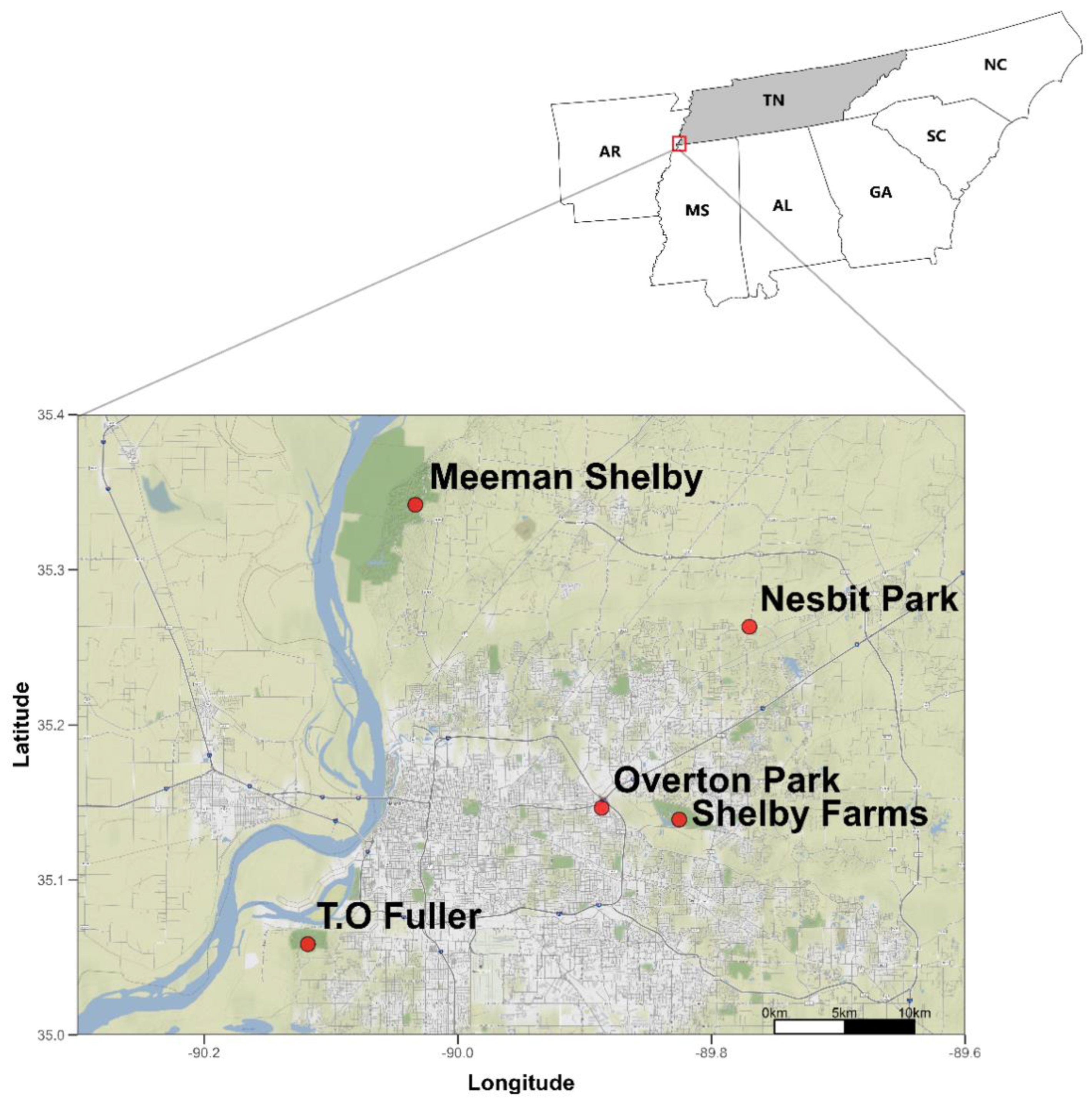
2.1. Study Area and Site Descriptions
2.2. Sample Collections and Laboratory Processing
2.3. Data Analyses
3. Results
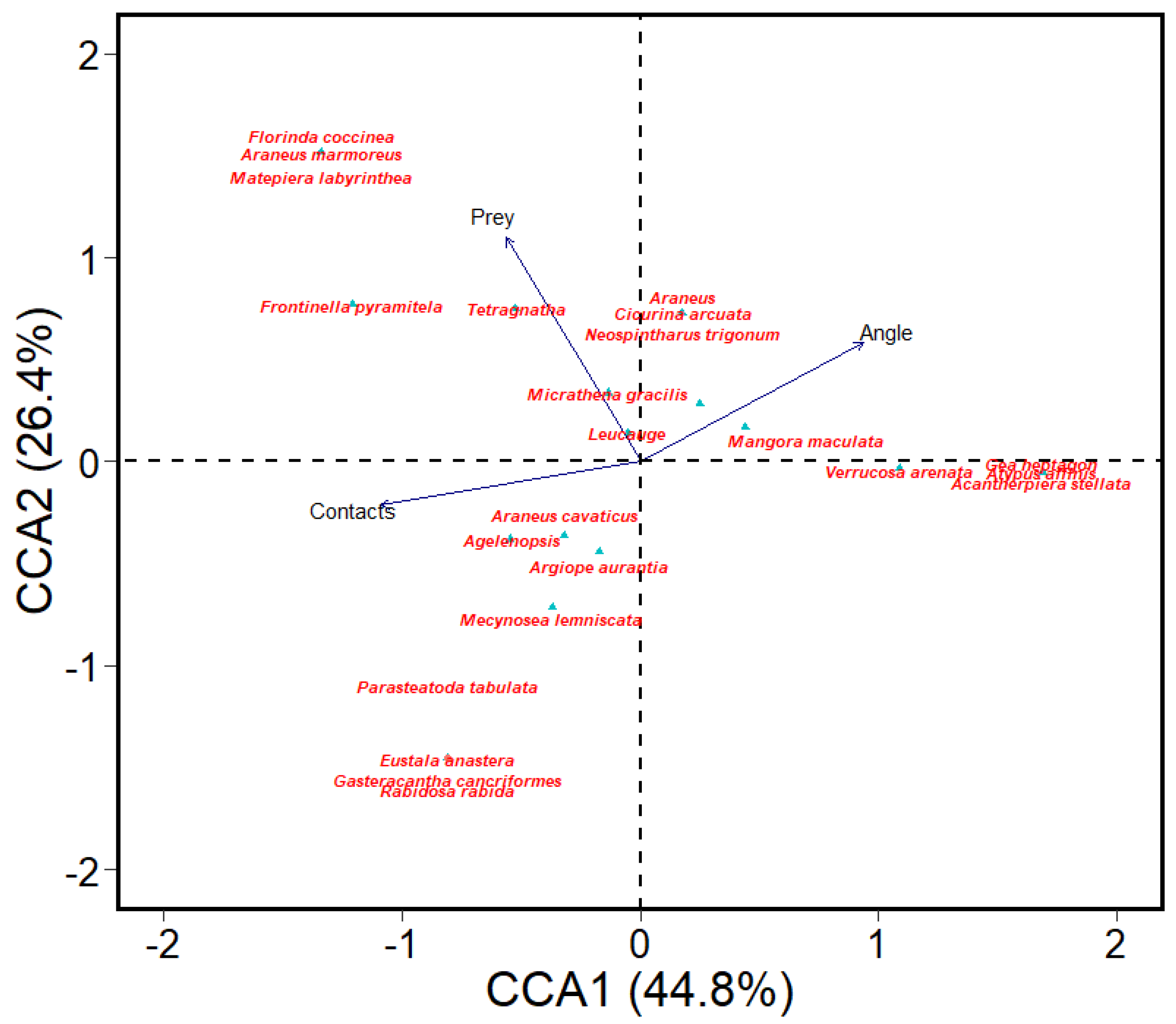
3.1. Species Occurrence and Predictors of Their Occurrence (Question 1)
3.2. Habitat Association and Indicator Species (Question 2)
4. Discussion
5. Conclusions
Author Contributions
Funding
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Nentwig, W.; Bacher, S.; Kumschick, S.; Pyšek, P.; Vilà, M. More than “100 worst” alien species in Europe. Biol. Invasions 2017, 20, 1611–1621. [Google Scholar] [CrossRef] [Green Version]
- Duncan, C.A.; Jachetta, J.J.; Brown, M.L.; Carrithers, V.F.; Clark, J.K.; Ditomaso, J.M.; Lym, R.G.; McDaniel, K.C.; Renz, M.J.; Rice, P.M. Assessing the Economic, Environmental, and Societal Losses from Invasive Plants on Rangeland and Wildlands. Weed Technol. 2004, 18, 1411–1416. [Google Scholar] [CrossRef]
- Brunson, M.W.; Tanaka, J. Economic and Social Impacts of Wildfires and Invasive Plants in American Deserts: Lessons From the Great Basin. Rangel. Ecol. Manag. 2011, 64, 463–470. [Google Scholar] [CrossRef] [Green Version]
- Bellard, C.; Cassey, P.; Blackburn, T.M. Alien species as a driver of recent extinctions. Biol. Lett. 2016, 12, 20150623. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Simberloff, D.; Martin, J.-L.; Genovesi, P.; Maris, V.; Wardle, D.A.; Aronson, J.; Courchamp, F.; Galil, B.; García-Berthou, E.; Pascal, M.; et al. Impacts of biological invasions: What’s what and the way forward. Trends Ecol. Evol. 2013, 28, 58–66. [Google Scholar] [CrossRef] [Green Version]
- Hoffmann, B.D.; Broadhurst, L.M. The economic cost of managing invasive species in Australia. NeoBiota 2016, 31, 1–18. [Google Scholar] [CrossRef] [Green Version]
- Seebens, H.; Blackburn, T.M.; Dyer, E.E.; Genovesi, P.; Hulme, P.E.; Jeschke, J.M.; Pagad, S.; Pyšek, P.; Winter, M.; Arianoutsou, M.; et al. No saturation in the accumulation of alien species worldwide. Nat. Commun. 2017, 8, 14435. [Google Scholar] [CrossRef]
- Hulme, P.E. Trade, transport and trouble: Managing invasive species pathways in an era of globalization. J. Appl. Ecol. 2009, 46, 10–18. [Google Scholar] [CrossRef]
- Elton, C.S. The Ecology of Invasions by Animals and Plants, 2nd ed.; Springer Nature: Cham, Switzerland, 2020; ISBN 978-3-030-34720-8. [Google Scholar]
- Doody, J.S.; Green, B.; Rhind, D.; Castellano, C.M.; Sims, R.; Robinson, T. Population-level declines in Australian predators caused by an invasive species. Anim. Conserv. 2009, 12, 46–53. [Google Scholar] [CrossRef]
- Nsengimana, V.; Kaplin, A.B.; Nsabimana, D.; Dekoninck, W.; Francis, F. Diversity and abundance of soil-litter arthropods and their relationships with soil physicochemical properties under different land uses in Rwanda. Biodiversity 2021, 22, 41–51. [Google Scholar] [CrossRef]
- Diagne, C.; Leroy, B.; Vaissière, A.-C.; Gozlan, R.E.; Roiz, D.; Jarić, I.; Salles, J.-M.; Bradshaw, C.J.A.; Courchamp, F. High and rising economic costs of biological invasions worldwide|Nature. Nature 2021, 592, 571–576. [Google Scholar] [CrossRef] [PubMed]
- Crowley, S.L.; Hinchliffe, S.; McDonald, R.A. Conflict in invasive species management. Front. Ecol. Evol. 2017, 15, 133–141. [Google Scholar] [CrossRef] [Green Version]
- Cordell, S.; Ostertag, R.; Michaud, J.; Warman, L. Quandaries of a decade-long restoration experiment trying to reduce invasive species: Beat them, join them, give up, or start over? Restor. Ecol. 2016, 24, 139–144. [Google Scholar] [CrossRef]
- Gleditsch, J.M.; Carlo, T.A. Fruit quantity of invasive shrubs predicts the abundance of common native avian frugivores in central Pennsylvania. Divers. Distrib. 2010, 17, 244–253. [Google Scholar] [CrossRef]
- Landsman, A.P.; Schmit, J.P.; Matthews, E.R. Invasive Plants Differentially Impact Forest Invertebrates, Providing Taxon-Specific Benefits by Enhancing Structural Complexity. Front. Ecol. Evol. 2021, 9, 682140. [Google Scholar] [CrossRef]
- Loomis, J.D.; Cameron, G.N.; Uetz, G.W. Impact of the Invasive Shrub Lonicera maackii on Shrub-Dwelling Araneae in a Deciduous Forest in Eastern North America. Am. Midl. Nat. 2014, 171, 204–218. [Google Scholar] [CrossRef]
- Langellotto, G.A.; Denno, R.F. Responses of Invertebrate Natural Enemies to Complex-Structured Habitats: A Meta-Analytical Synthesis. Available online: https://link.springer.com/article/10.1007/s00442-004-1497-3 (accessed on 12 October 2022).
- Goodenough, A. Are the ecological impacts of alien species misrepresented? A review of the “native good, alien bad” philosophy. Community Ecol. 2010, 11, 13–21. [Google Scholar] [CrossRef]
- Didham, R.K.; Tylianakis, J.M.; Hutchison, M.A.; Ewers, R.M.; Gemmell, N.J. Are invasive species the drivers of ecological change? Trends Ecol. Evol. 2005, 20, 470–474. [Google Scholar] [CrossRef]
- Schirmel, J.; Bundschuh, M.; Entling, M.H.; Kowarik, I.; Buchholz, S. Impacts of invasive plants on resident animals across ecosystems, taxa, and feeding types: A global assessment. Glob. Change Biol. 2016, 22, 594–603. [Google Scholar] [CrossRef]
- Brown, J.H.; Sax, D.F. An Essay on Some Topics Concerning Invasive Species. Austral Ecol. 2004, 29, 530–536. [Google Scholar] [CrossRef]
- Cash, J.; Anderson, C.; Gulsby, W. The ecological effects of Chinese privet (Ligustrum sinense) invasion. Invasive Plant Sci. Manag. 2020, 13, 3–13. [Google Scholar] [CrossRef]
- Birkhofer, K.; Nyffeler, M. An estimated 400–800 million tons of prey are annually killed by the global spider community. Sci. Nat. 2017, 104, 30. [Google Scholar] [CrossRef] [Green Version]
- Buchholz, S. Ground spider assemblages as indicators for habitat structure in inland sand ecosystems. Biodivers. Conserv. 2010, 19, 2565–2595. [Google Scholar] [CrossRef]
- Ameline, C.; Puzin, C.; Bowden, J.J.; Lambeets, K.; Vernon, P.; Pétillon, J. Habitat specialization and climate affect arthropod fitness: A comparison of generalist vs. specialist spider species in Arctic and temperate biomes. Biol. J. Linn. Soc. 2017, 121, 592–599. [Google Scholar] [CrossRef]
- Buchholz, S.; Tietze, H.; Kowarik, I.; Schirmel, J. Effects of a Major Tree Invader on Urban Woodland Arthropods. PLoS ONE 2015, 10, e0137723. [Google Scholar] [CrossRef] [Green Version]
- Harry, I.; Höfer, H.; Schielzeth, H.; Assmann, T. Protected habitats of Natura 2000 do not coincide with important diversity hotspots of arthropods in mountain grasslands. Insect Conserv. Divers. 2019, 12, 329–338. [Google Scholar] [CrossRef]
- Clausen, I.H.S. The use of spiders Araneaeas ecological indicators*. Br. Arachnol. Soc. 1986, 7, 83–86. [Google Scholar]
- Hore, U.; Uniyal, V.; Nentwig, W.; Entling, M.; Kropf, C. Influence of space, vegetation structure, and microclimate on spider (Araneae) species composition in Terai Conservation Area, India. Eur. Arachnol. 2010, 2008, 71–77. [Google Scholar]
- Vasconcellos-Neto, J.; Romero, G.Q.; Santos, A.J.; Dippenaar-Schoeman, A.S. Associations of Spiders of the Genus Peucetia (Oxyopidae) with Plants Bearing Glandular Hairs. Biotropica 2007, 39, 221–226. [Google Scholar] [CrossRef]
- Scheidler, M. Influence of habitat structure and vegetation architecture on spiders. Zool. Anz. 1990, 225, 333–340. [Google Scholar]
- Knight, D.B.; Davis, R.E. Climatology of Tropical Cyclone Rainfall in the Southeastern United States. Phys. Geogr. 2007, 28, 126–147. [Google Scholar] [CrossRef]
- Schmid, P.E. Observing and Modeling Urban Thunderstorm Modification Due to Land Surface and Aerosol Effects. Ph.D. Thesis, Purdue University Graduate School, West Lafayette, IN, USA, 2020. [Google Scholar] [CrossRef]
- Kahle, D.; Wickham, H. ggmap: Spatial Visualization with ggplot2. R J. 2013, 5, 144–161. [Google Scholar] [CrossRef]
- Meeman-Shelby Forest State Park Meeman-Shelby Forest State Park Business & Management Plan 2018. Available online: https://tnstateparks.com/assets/pdf/park-plans/June2018MeemanShelbyBP.pdf (accessed on 24 August 2022).
- West, C.V. Meeman-Shelby Forest State Park. Available online: https://tennesseeencyclopedia.net/entries/meeman-shelby-forest-state-park (accessed on 3 October 2022).
- Trust for Public Land Tennessee Office The Parks of Memphis—Past, Present & Future 2014. Available online: https://www.tpl.org/wp-content/uploads/2015/07/ccpe-memphis-parks-report.pdf (accessed on 10 September 2022).
- Green, J. Sampling Method and Time Determines Composition of Spider Collections. J. Arachnol. 1999, 27, 176–182. [Google Scholar]
- Ausden, M.; Drake, M. Invertebrates. In Ecological Census Techniques; Sutherland, W., Ed.; Cambridge University Press: Cambridge, UK, 2006; pp. 214–249. [Google Scholar]
- McCaffrey, J.P.; Parrella, M.P.; Horsburgh, R.L. Evaluation of the Limb-Beating Sampling Method for Estimating Spider (Araneae) Populations on Apple Trees. J. Arachnol. 1983, 11, 363–368. [Google Scholar]
- Whitmore, C.; Slotow, R.; Crouch, T.E.; Dippenaar-Schoeman, A.S. Diversity of spiders (Araneae) in a Savanna reserve, Northern province, South Africa. Arac 2002, 30, 344–356. [Google Scholar] [CrossRef]
- Bishop, L.; Connolly, S.R. Web Orientation, Thermoregulation, and Prey Capture Efficiency in a Tropical Forest Spider. J. Arachnol. 1992, 20, 173–178. [Google Scholar]
- Eberhard, W.G. The ecology of the web of Uloborus diversus (Araneae: Uloboridae). Oecologia 1971, 6, 328–342. [Google Scholar] [CrossRef]
- Wu, C.-C.; Blamires, S.J.; Wu, C.-L.; Tso, I.-M. Wind induces variations in spider web geometry and sticky spiral droplet volume. J. Exp. Biol. 2013, 216, 3342–3349. [Google Scholar] [CrossRef] [Green Version]
- Buskirk, R.E. Coloniality, Activity Patterns and Feeding in a Tropical Orb-Weaving Spider. Ecology 1975, 56, 1314–1328. [Google Scholar] [CrossRef]
- Eberhard, W.G. Function and Phylogeny of Spider Webs. Annu. Rev. Ecol. Syst. 1990, 21, 341–372. [Google Scholar] [CrossRef]
- Balfour, R.A.; Rypstra, A.L. The Influence of Habitat Structure on Spider Density in a No-Till Soybean Agroecosystem. J. Arachnol. 1998, 26, 221–226. [Google Scholar]
- Mcnett, B.J.; Rypstra, A.L. Habitat selection in a large orb-weaving spider: Vegetational complexity determines site selection and distribution. Ecol. Entomol. 2000, 25, 423–432. [Google Scholar] [CrossRef]
- Ubick, D.; Cushing, P.E. Spiders of North America: An Identification Manual, Second Edition, 2nd ed.; American Arachnological Society: Keene, NH, USA, 2017; ISBN 978-0-9980146-0-9. [Google Scholar]
- Bradley, R.A. Common Spiders of North America, 1st ed.; University of California Press: Berkeley, CA, USA, 2019; ISBN 978-0-520-31531-0. [Google Scholar]
- Legendre, P.; Gallagher, E.D. Ecologically meaningful transformations for ordination of species data. Oecologia 2001, 129, 271–280. [Google Scholar] [CrossRef]
- Oksanen, J.; Blanchet, F.G.; Kindt, R.; Legendre, P.; O’Hara, R.; Simpson, G.; Solymos, P.; Stevens, M.; Wagner, H. Package ‘vegan’: Community Ecology Package. Version 1.17–2. 2010. Available online: https://www.researchgate.net/publication/282247921_Package_%27vegan%27_Community_Ecology_Package_Version_117--2 (accessed on 22 November 2022).
- Oksanen, J.; Blanchet, F.G.; Kindt, R.; Legendre, P.; Minchin, P.; O’Hara, B.; Simpson, G.; Solymos, P.; Stevens, H.; Wagner, H. Vegan: Community Ecology Package. R Package Version 2.2–1. 2015, Volume 2, pp. 1–2. Available online: http://www.pelagicos.net/MARS6910_spring2015/manuals/R_vegan.pdf (accessed on 22 November 2022).
- Dufrêne, M.; Legendre, P. Species Assemblages and Indicator Species:the Need for a Flexible Asymmetrical Approach. Ecol. Monogr. 1997, 67, 345–366. [Google Scholar] [CrossRef]
- Wildi, O.; Feldmeyer-Christe, E. Indicator values (IndVal) mimic ranking by F-ratio in real-world vegetation data. Community Ecol. 2013, 14, 139–143. [Google Scholar] [CrossRef]
- R Core Team. A Language and Environment for Statistical Computing; R Foundation for Statistical Computing: Vienna, Austria, 2022. [Google Scholar]
- Ledin, A.E.; Styrsky, J.D.; Styrsky, J.N. Friend or Foe? Orb-Weaver Spiders Inhabiting Ant–Acacias Capture Both Herbivorous Insects and Acacia Ant Alates. J. Insect Sci. 2020, 20, 16. [Google Scholar] [CrossRef]
- Hesselberg, T.; Triana, E. The web of the acacia orb-spider Eustala illicita (Araneae: Araneidae) with notes on its natural history. J. Arachnol. 2010, 38, 21–26. [Google Scholar] [CrossRef]
- Clough, Y.; Kruess, A.; Kleijn, D.; Tscharntke, T. Spider diversity in cereal fields: Comparing factors at local, landscape and regional scales. J. Biogeogr. 2005, 32, 2007–2014. [Google Scholar] [CrossRef]
- Smallwood, P.D. Web-Site Tenure in the Long-Jawed Spider: Is It Risk-Sensitive Foraging, or Conspecific Interactions? Ecology 1993, 74, 1826–1835. [Google Scholar] [CrossRef]
- Whitney, K.D. Experimental Evidence That Both Parties Benefit in a Facultative Plant-Spider Mutualism. Ecology 2004, 85, 1642–1650. [Google Scholar] [CrossRef] [Green Version]
- Dennis, P.; Young, M.R.; Gordon, I.J. Distribution and abundance of small insects and arachnids in relation to structural heterogeneity of grazed, indigenous grasslands. Ecol. Entomol. 1998, 23, 253–264. [Google Scholar] [CrossRef]
- Malumbres-Olarte, J.; Vink, C.J.; Ross, J.G.; Cruickshank, R.H.; Paterson, A.M. The role of habitat complexity on spider communities in native alpine grasslands of New Zealand. Insect Conserv. Divers. 2013, 6, 124–134. [Google Scholar] [CrossRef]
- Sieg, C.H.; Phillips, B.G.; Moser, L.P. Exotic invasive plants. In Ecological Restoration of Southwestern Ponderosa Pine Forests; Frederici, P., Ed.; Island Press: Washington, DC, USA, 2003; pp. 251–267. [Google Scholar]
- Norbury, G.; Heyward, R.; Parkes, J. Skink and invertebrate abundance in relation to vegetation, rabbits and predators in a New Zealand dryland ecosystem on JSTOR. N. Z. J. Ecol. 2009, 33, 24–31. [Google Scholar]
- Hayes, S.J.; Holzmueller, E.J. Relationship between Invasive Plant Species and Forest Fauna in Eastern North America. Forests 2012, 3, 840–852. [Google Scholar] [CrossRef] [Green Version]
- Wang, H.-H.; Grant, W.E. Invasion of Eastern Texas Forestlands by Chinese Privet: Efficacy of Alternative Management Strategies. Diversity 2014, 6, 652–664. [Google Scholar] [CrossRef] [Green Version]
- Gaertner, M.; Breeyen, A.D.; Hui, C.; Richardson, D.M. Impacts of alien plant invasions on species richness in Mediterranean-type ecosystems: A meta-analysis. Prog. Phys. Geogr. Earth Environ. 2009, 33, 319–338. [Google Scholar] [CrossRef]
- Green, S.J.; Grosholz, E.D. Functional eradication as a framework for invasive species control. Front. Ecol. Environ. 2021, 19, 98–107. [Google Scholar] [CrossRef]



{kind=link}
{kind=link}
| Site | Description | Dominant Plants | Invasive |
|---|---|---|---|
| Shelby Farms Park | 4500-acre public park that was opened for public recreation by the Shelby County government in the 1970s; is heavily managed, including forest trail upkeep. | Winged Elm Slippery Elm Privet | No No Yes |
| Overton Park Conservatory | 184-acre public park opened in 2011 with heavy maintenance, including invasive species removals and trail upkeep. | Hornwort Pawpaw Box Elder Sweetgum | No No No No |
| Nesbit Park | 333.75-acre park owned by cycling club company Stanking Creek Cycling open for free public use. No explicit plant management other than trail upkeep. | Jumpseed Pawpaw Winged Elm Sweetgum | No No No No |
| Meeman-Shelby State Park | 13,469-acre state park initially built in the 1930s by the National Park Service; had land clearing and replanting efforts when built but currently has less spending management than Shelby Farms or Overton Parks. | Hophornbeam Beech Hophornbeam Sugar Maple | No No No No |
| T.O. Fuller State Park | 1138-acre state park opened in 1938; has public recreation options but significantly lower revenues and spending than Shelby Farms and Overton Parks in recent years. | Pawpaw Privet Winged Elm Slippery Elm | No Yes No No |
| Park | Species | IndVal | p | Freq |
|---|---|---|---|---|
| Shelby Farms | Eustala anastera | 1.00 | 0.013 | 3 |
| Shelby Farms | Parasteatoda tabulata | 1.00 | 0.01 | 3 |
| Shelby Farms | Gasteracantha cancriformes | 1.00 | 0.014 | 3 |
| Shelby Farms | Rabidosa rabida | 1.00 | 0.009 | 3 |
| Shelby Farms | Mecynogea lemniscata | 0.58 | 0.012 | 9 |
| Shelby Farms | Agelenopsis sp. | 0.53 | 0.012 | 12 |
| Overton Park | Cicurina arcuata | 1.00 | 0.012 | 3 |
| Overton Park | Araneid sp. | 1.00 | 0.013 | 3 |
| Overton Park | Neospintharus trigonum | 1.00 | 0.016 | 3 |
| Overton Park | Leucauge venusta | 0.36 | 0.014 | 15 |
| Meeman-Shelby | Florinda coccinea | 1.00 | 0.013 | 3 |
| Meeman-Shelby | Araneus marmoreus | 1.00 | 0.009 | 3 |
| Meeman-Shelby | Melpomene sp. | 1.00 | 0.011 | 3 |
| Meeman-Shelby | Metepiera labyrinthea | 1.00 | 0.022 | 3 |
| Meeman-Shelby | Frontinella pyramitela | 0.75 | 0.013 | 6 |
| T.O Fuller | Atypus affinis | 1.00 | 0.01 | 3 |
| T.O Fuller | Acantherpiera stellata | 1.00 | 0.009 | 3 |
| T.O Fuller | Gea heptagon | 1.00 | 0.015 | 3 |
| T.O Fuller | Verrucosa arenata | 0.74 | 0.017 | 12 |
| Nesbit | Mangora maculata | 0.50 | 0.015 | 15 |
| Tree Species | Species | IndVal | p | Freq |
|---|---|---|---|---|
| Beech | Araneus marmoreus | 0.83 | 0.001 | 5 |
| Beech | Florinda coccinea | 0.43 | 0.002 | 9 |
| Black walnut | Frontinella pyramitela | 0.33 | 0.001 | 36 |
| Black walnut | Argiope aurantia | 0.27 | 0.001 | 61 |
| Black walnut | Mangora maculata | 0.08 | 0.001 | 122 |
| Black walnut | Mecynogea lemniscata | 0.07 | 0.001 | 151 |
| Boxelder | Micrathena gracilis | 0.11 | 0.001 | 120 |
| Boxelder | Verrucosa arenata | 0.09 | 0.001 | 121 |
| Deadwood | Parasteatoda tabulata | 0.74 | 0.001 | 17 |
| Deadwood | Rabidosa rabida | 0.74 | 0.001 | 17 |
| Deadwood | Agelenopsis pennsylvanica | 0.17 | 0.016 | 45 |
| Deadwood | Leucauge venusta | 0.06 | 0.001 | 172 |
| Green ash | Metepeira labyrinthea | 1.00 | 0.001 | 3 |
| Japanese honeysuckle | Agelenopsis aperta | 0.51 | 0.001 | 35 |
| Jumpseed and hornwort | Leucage sp. | 0.14 | 0.001 | 91 |
| Pawpaw | Gea heptagon | 1.00 | 0.001 | 17 |
| Pawpaw | Araneus cavaticus | 0.71 | 0.001 | 23 |
| Pawpaw | Micrathena sagittata | 0.34 | 0.001 | 32 |
| Chinese privet | Eustala anastera | 0.97 | 0.001 | 31 |
| Slippery elm | Acantherpeira stellata | 1.00 | 0.001 | 12 |
| Slippery elm | Gasteracantha cancriformis | 1.00 | 0.001 | 12 |
| Slippery elm | Tetragnatha sp. | 0.53 | 0.001 | 20 |
| Tulip poplar | Cicurina arcuata | 1.00 | 0.001 | 5 |
| Virginia creeper | Melpomene sp. | 1.00 | 0.001 | 5 |
| White oak | Araneus sp. | 0.60 | 0.001 | 9 |
| White oak | Atypus affinis | 0.40 | 0.009 | 6 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Ellsworth, E.; Li, Y.; Chari, L.D.; Kron, A.; Moyo, S. Tangled in a Web: Management Type and Vegetation Shape the Occurrence of Web-Building Spiders in Protected Areas. Insects 2022, 13, 1129. https://doi.org/10.3390/insects13121129
Ellsworth E, Li Y, Chari LD, Kron A, Moyo S. Tangled in a Web: Management Type and Vegetation Shape the Occurrence of Web-Building Spiders in Protected Areas. Insects. 2022; 13(12):1129. https://doi.org/10.3390/insects13121129
Chicago/Turabian StyleEllsworth, El, Yihan Li, Lenin D. Chari, Aidan Kron, and Sydney Moyo. 2022. "Tangled in a Web: Management Type and Vegetation Shape the Occurrence of Web-Building Spiders in Protected Areas" Insects 13, no. 12: 1129. https://doi.org/10.3390/insects13121129