

Mechanics of the Prey Capture Technique of the South African Grassland Bolas Spider, Cladomelea akermani

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:Simple Summary
Abstract
1. Introduction
2. Materials and Methods
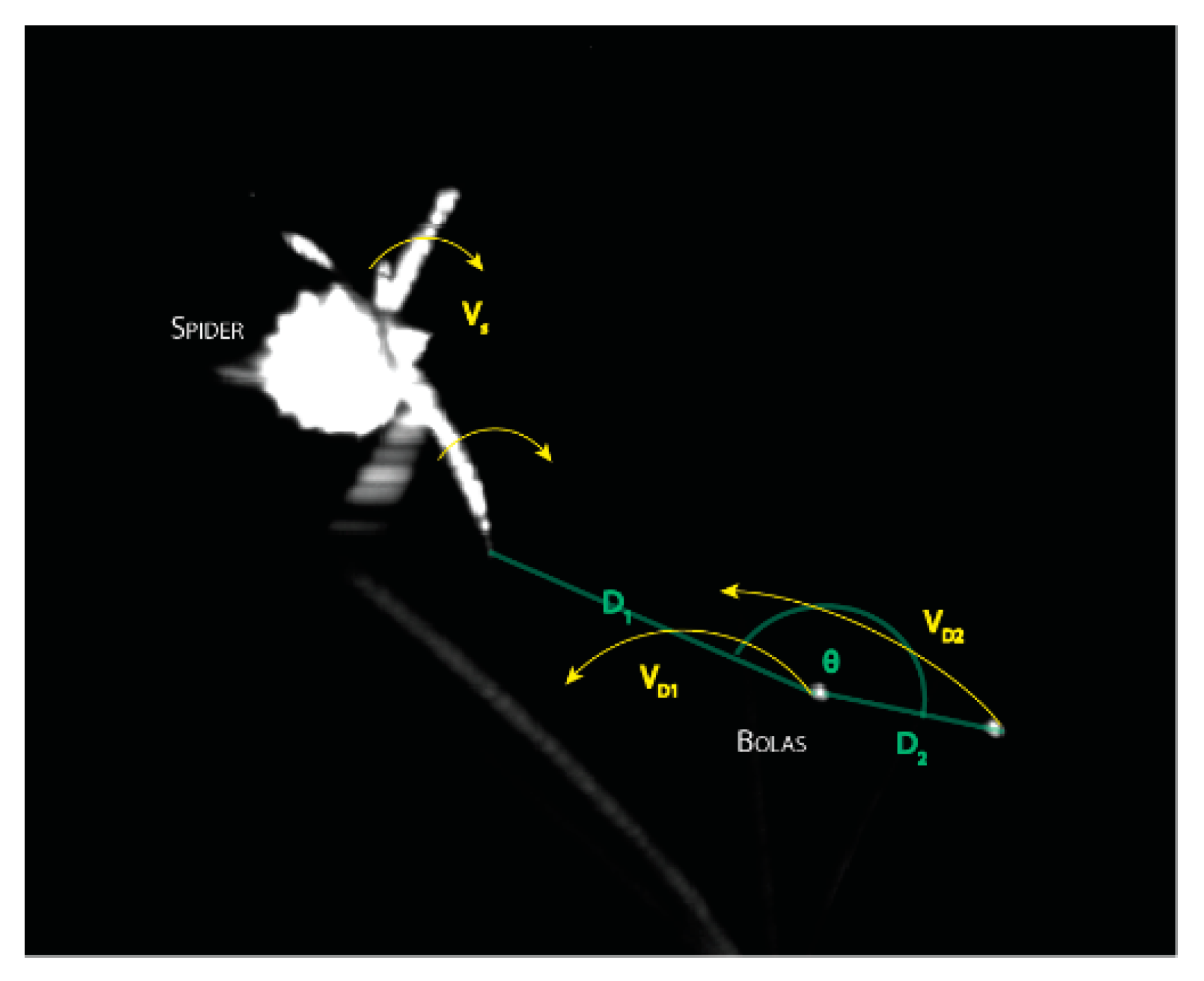
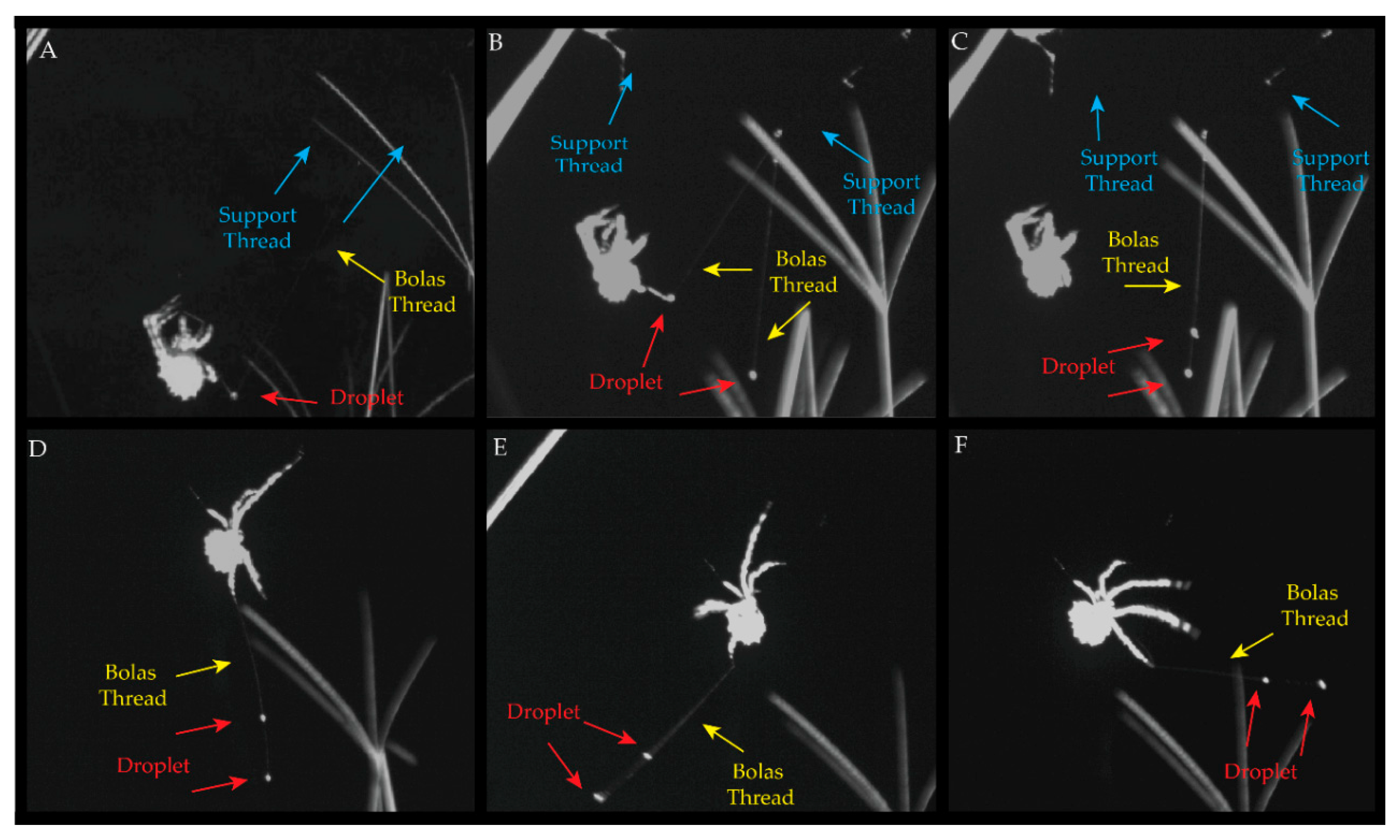
2.1. Field Observations and Kinematics of Prey Capture
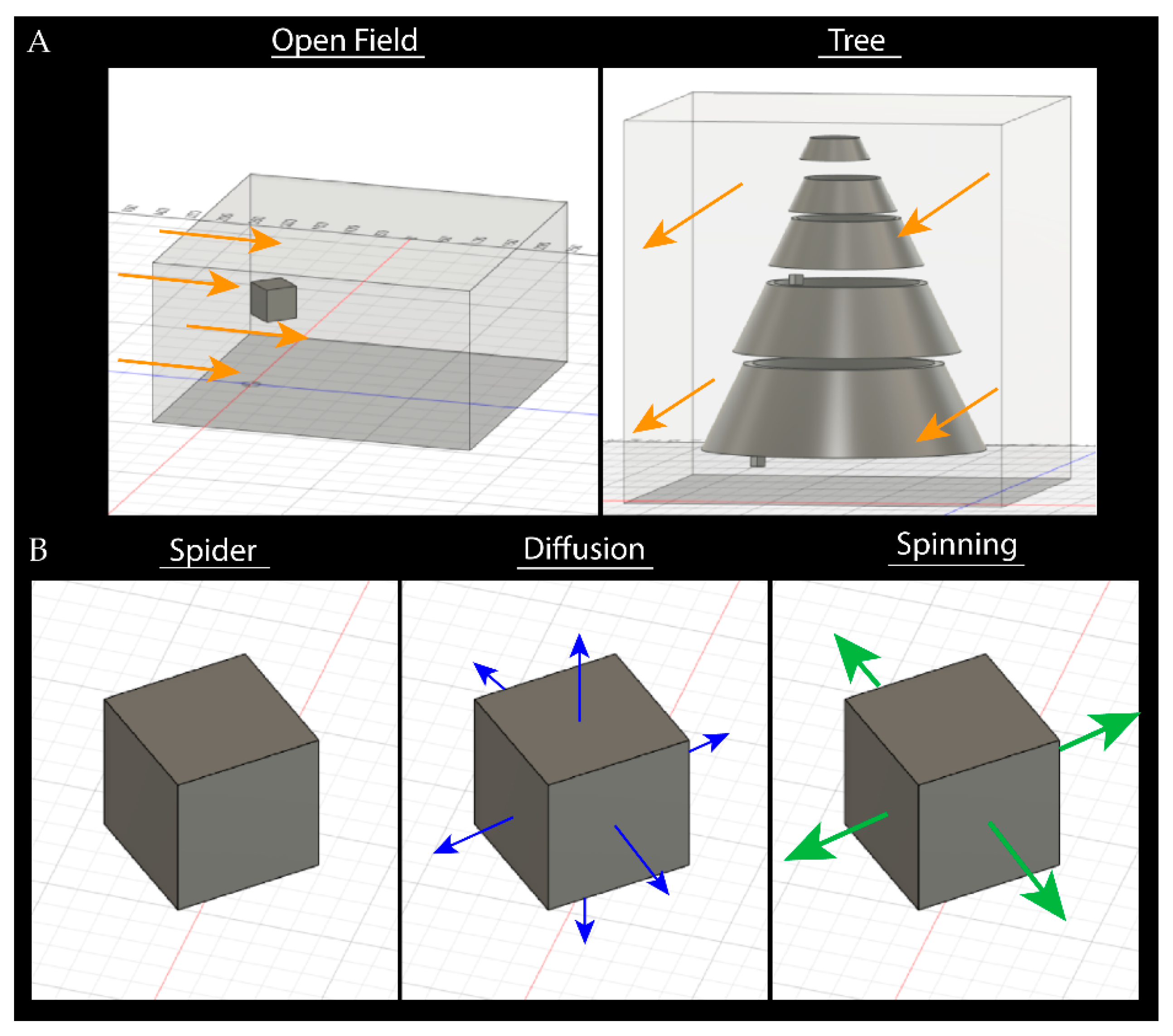
2.2. Pheromone Airflow—Computational Fluid Dynamics Model
3. Results
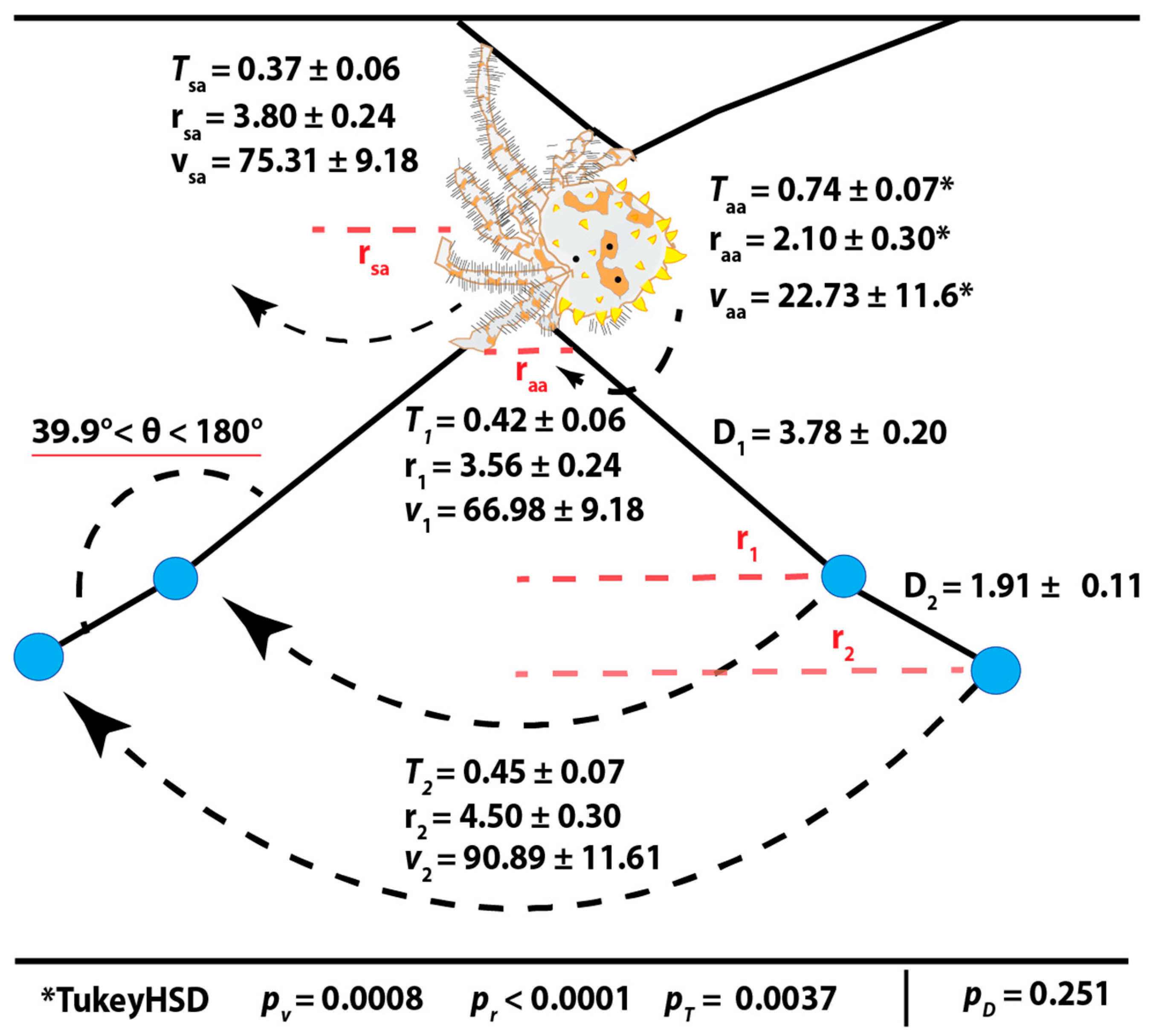
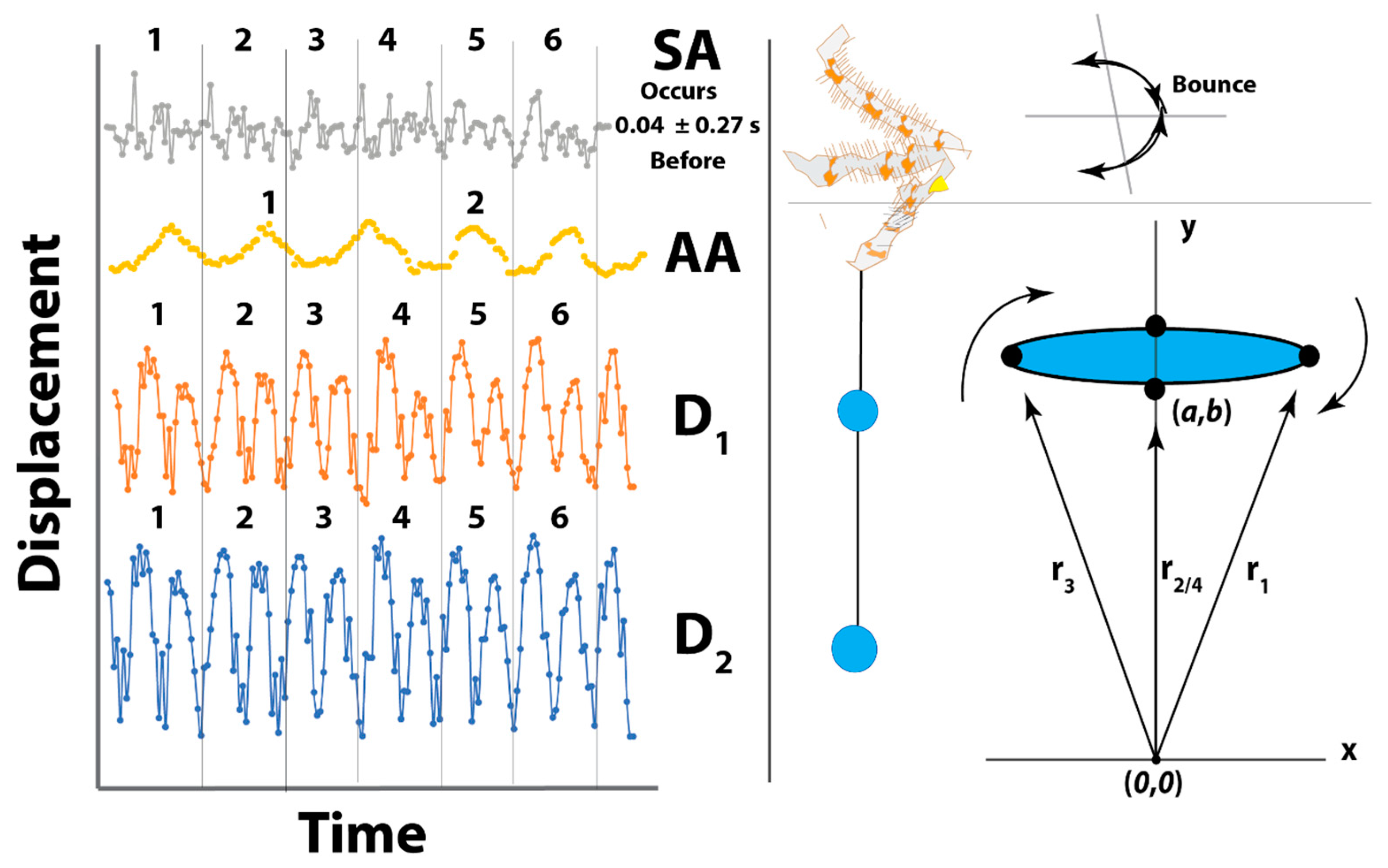
3.1. Observations on Bolas Building Behavior and Kinematics of Prey Capture
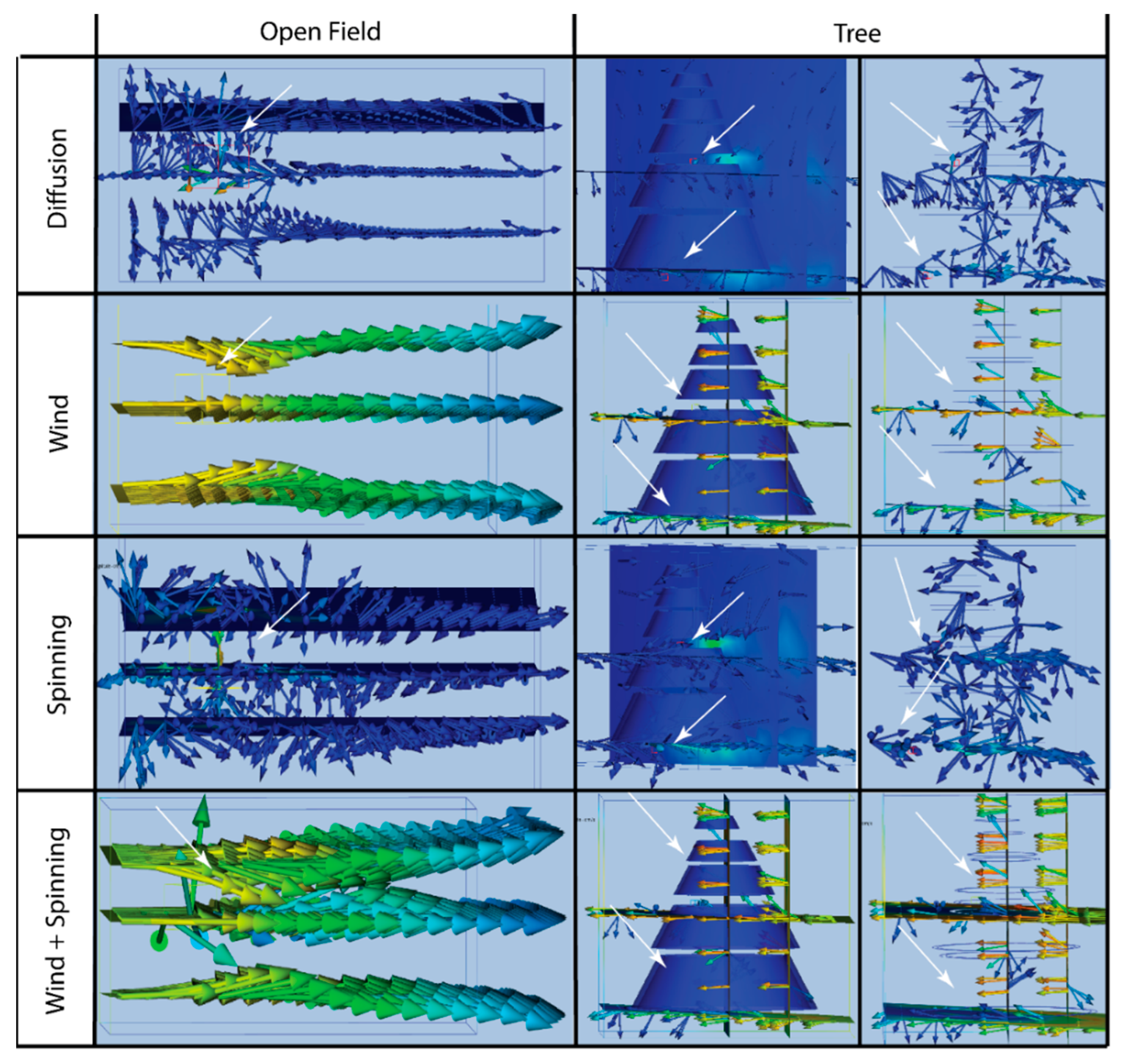
3.2. Pheromone Airflow—Computational Fluid Dynamics Model
4. Discussion
Supplementary Materials
Author Contributions
Funding
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Foelix, R.F. Biology of Spiders; Oxford University Press: New York, NY, USA, 2011. [Google Scholar]
- Blackledge, T.A.; Scharff, N.; Coddington, J.A.; Szüts, T.; Wenzel, J.W.; Hayashi, C.Y.; Agnarsson, I. Reconstructing web evolution and spider diversification in the molecular era. Proc. Natl. Acad. Sci. USA 2009, 106, 5229–5234. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Nentwig, W. Why do only certain insects escape from a spider’s web? Oecologia 1982, 53, 412–417. [Google Scholar] [CrossRef] [PubMed]
- Diaz, C.; Maksuta, D.; Amarpuri, G.; Tanikawa, A.; Miyashita, T.; Dhinojwala, A.; Blackledge, T.A. The moth specialist spider Cyrtarachne akirai uses prey scales to increase adhesion. J. R. Soc. Interface 2020, 17, 20190792. [Google Scholar] [CrossRef] [Green Version]
- Levi, H.W. The Bolas Spiders of the Genus Mastophora (Araneae: Araneidae); Harvard University: Cambridge, MA, USA, 2003. [Google Scholar]
- Tanikawa, A. Two new species of the genus Cyrtarachne (Araneae: Araneidae) from Japan hitherto identified as C. inaequalis. Acta Arachnol. 2013, 62, 95–101. [Google Scholar] [CrossRef] [Green Version]
- Baba, Y.G.; Kusahara, M.; Maezono, Y.; Miyashita, T. Adjustment of web-building initiation to high humidity: A constraint by humidity-dependent thread stickiness in the spider Cyrtarachne. Naturwissenschaften 2014, 101, 587–593. [Google Scholar] [CrossRef]
- Diaz, C.; Tanikawa, A.; Miyashita, T.; Amarpuri, G.; Jain, D.; Dhinojwala, A.; Blackledge, T.A. Supersaturation with water explains the unusual adhesion of aggregate glue in the webs of the moth-specialist spider, Cyrtarachne akirai. R. Soc. Open Sci. 2018, 5, 181296. [Google Scholar] [CrossRef] [Green Version]
- Diaz, C.; Tanikawa, A.; Miyashita, T.; Dhinojwala, A.; Blackledge, T.A. Silk structure rather than tensile mechanics explains web performance in the moth-specialized spider, Cyrtarachne. J. Exp. Zool. A Ecol. Integr. Physiol. 2018, 329, 120–129. [Google Scholar] [CrossRef] [PubMed]
- Andersen, S.O. Biochemistry of insect cuticle. Annu. Rev. Entomol. 1979, 24, 29–59. [Google Scholar] [CrossRef]
- Cartan, C.K.; Miyashita, T. Extraordinary web and silk properties of Cyrtarachne (Araneae, Araneidae): A possible link between orb-webs and bolas. Biol. J. Linn. Soc. 2000, 71, 219–235. [Google Scholar] [CrossRef]
- Eberhard, W.G. Aggressive chemical mimicry by a bolas spider. Science 1977, 198, 1173–1175. [Google Scholar] [CrossRef]
- Eberhard, W.G. The natural history and behavior of the bolas spider Mastophora dizzydeani sp. n.(Araneidae). Psyche 1980, 87, 143–169. [Google Scholar] [CrossRef] [Green Version]
- Stowe, M.K. Prey specialization in the Araneidae. In Spiders-Webs, Behavior, and Evolution; Stanford University Press: Redwood City, CA, USA, 1986; pp. 101–131. [Google Scholar]
- Yeargan, K.V. Ecology of a bolas spider, Mastophora hutchinsoni: Phenology, hunting tactics, and evidence for aggressive chemical mimicry. Oecologia 1988, 74, 524–530. [Google Scholar] [CrossRef] [PubMed]
- Yeargan, K.V. Biology of bolas spiders. Annu. Rev. Entomol. 1994, 39, 81–99. [Google Scholar] [CrossRef]
- Tanikawa, A. Japanese Spiders of the Genus Ordgarius (Araneae: Araneidae). Acta Arachnol. 1997, 46, 101–110. [Google Scholar] [CrossRef] [Green Version]
- Diaz, C.; Long, J.H. Behavior and Bioadhesives: How Bolas Spiders, Mastophora hutchinsoni Catch Moths. Insect, 2022; submitted for review. [Google Scholar]
- Gertsch, W.J. The North American bolas spiders of the genera Mastophora and Agatostichus. Bull. Am. Mus. 1955, 106, 225–254. [Google Scholar]
- Dippenaar-Schoeman, A.S.; Jocqué, R. African Spiders: An Identification Manual; ARC-Plant Protection Research Institute: Pretoria, South Africa, 1997; Volume 9. [Google Scholar]
- Leroy, J.M.; Jocqué, R.; Leroy, A. On the behaviour of the African bolas-spider Cladomelea akermani Hewitt (Araneae, Araneidae, Cyrtarachninae), with description of the male. Ann. Natal Mus. Pietermaritzbg. 1998, 39, 1–9. [Google Scholar]
- Haynes, K.F.; Yeargan, K.V.; Gemeno, C. Detection of prey by a spider that aggressively mimics pheromone blends. J. Insect Behav. 2001, 14, 535–544. [Google Scholar] [CrossRef]
- Hutchinson, C.E. A bolas-throwing spider. Sci. Am. 1903, 89, 172. [Google Scholar] [CrossRef] [Green Version]
- Shinkai, A.; Shinkai, E. The natural history, bolas construction, and hunting behavior of the bolas spider, Ordgarius sexspinosus (Thorell) (Araneae: Araneidae). Acta Arachnol. 2002, 51, 149–154. [Google Scholar] [CrossRef]
- Scharff, N.; Hormiga, G. First evidence of aggressive chemical mimicry in the Malagasy orb weaving spider Exechocentrus lancearius Simon, 1889 (Arachnida: Araneae: Araneidae) and description of a second species in the genus. Arthropod Syst. Phylogeny 2012, 70, 107–118. [Google Scholar]
- Hewitt, J. On certain South African Arachnida, with descriptions of three new species. Ann. Natal Mus. 1923, 5, 55–66. [Google Scholar]
- Roff, J.; Dippenaar-Schoeman, A. Description of a new species of Cladomelea bolas-spider from South Africa, with notes on its behaviour (Araneae: Araneidae). Afr. Invertebr. 2004, 45, 1–6. [Google Scholar]
- Chlegant, J. Kinovea (0.8.15) [Computer Software]. Joan Chlegant and Contributions. 2011. Available online: Kinveo.org (accessed on 20 December 2021).
- Autodesk. Autodesk Fusion 360; Version 2.0.13881; Autodesk: San Rafael, CA, USA, 2022. [Google Scholar]
- Autodesk. Autodesk CFD 2023; Version 23.0.0.0; Autodesk: San Rafael, CA, USA, 2022. [Google Scholar]
- Fick, A. Ueber diffusion. Ann. Phys. 1855, 170, 59–86. [Google Scholar] [CrossRef]
- Magnus, G. Ueber die Abweichung der Geschosse, und: Ueber eine auffallende Erscheinung bei rotirenden Körpern. Ann. Phys. (Berl.) 1853, 164, 1–29. [Google Scholar] [CrossRef] [Green Version]
- Cardé, R.T.; Mafra-Neto, A. Mechanisms of flight of male moths to pheromone. In Insect Pheromone Research; Springer: Boston, MA, USA, 1997; pp. 275–290. [Google Scholar]
- Cardé, R.T. Navigation along windborne plumes of pheromone and resource-linked odors. Annu. Rev. Entomol. 2021, 66, 317–336. [Google Scholar] [CrossRef]
- Sahni, V.; Blackledge, T.A.; Dhinojwala, A. Viscoelastic solids explain spider web stickiness. Nat. Commun. 2010, 1, 19. [Google Scholar] [CrossRef] [Green Version]
- Sahni, V.; Blackledge, T.A.; Dhinojwala, A. Changes in the adhesive properties of spider aggregate glue during the evolution of cobwebs. Sci. Rep. 2011, 1, 41. [Google Scholar] [CrossRef] [Green Version]
- Elettro, H.; Neukirch, S.; Vollrath, F.; Antkowiak, A. In-drop capillary spooling of spider capture thread inspires hybrid fibers with mixed solid–liquid mechanical properties. Proc. Natl. Acad. Sci. USA 2016, 113, 6143–6147. [Google Scholar] [CrossRef] [Green Version]
- Miyashita, T.; Sakamaki, Y.; Shinkai, A. Evidence against moth attraction by Cyrtarachne, a genus related to bolas spiders. Acta Arachnol. 2001, 50, 1–4. [Google Scholar] [CrossRef]






Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Diaz, C., Jr.; Roff, J. Mechanics of the Prey Capture Technique of the South African Grassland Bolas Spider, Cladomelea akermani. Insects 2022, 13, 1118. https://doi.org/10.3390/insects13121118
Diaz C Jr., Roff J. Mechanics of the Prey Capture Technique of the South African Grassland Bolas Spider, Cladomelea akermani. Insects. 2022; 13(12):1118. https://doi.org/10.3390/insects13121118
Chicago/Turabian StyleDiaz, Candido, Jr., and John Roff. 2022. "Mechanics of the Prey Capture Technique of the South African Grassland Bolas Spider, Cladomelea akermani" Insects 13, no. 12: 1118. https://doi.org/10.3390/insects13121118