
Insecticide Susceptibility Status of Anopheles and Aedes Mosquitoes in Malaria and Dengue Endemic Areas, Thai–Myanmar Border
 , ,
, ,  , ,
, ,  and
and
Abstract
:Simple Summary
Abstract
1. Introduction
2. Materials and Methods
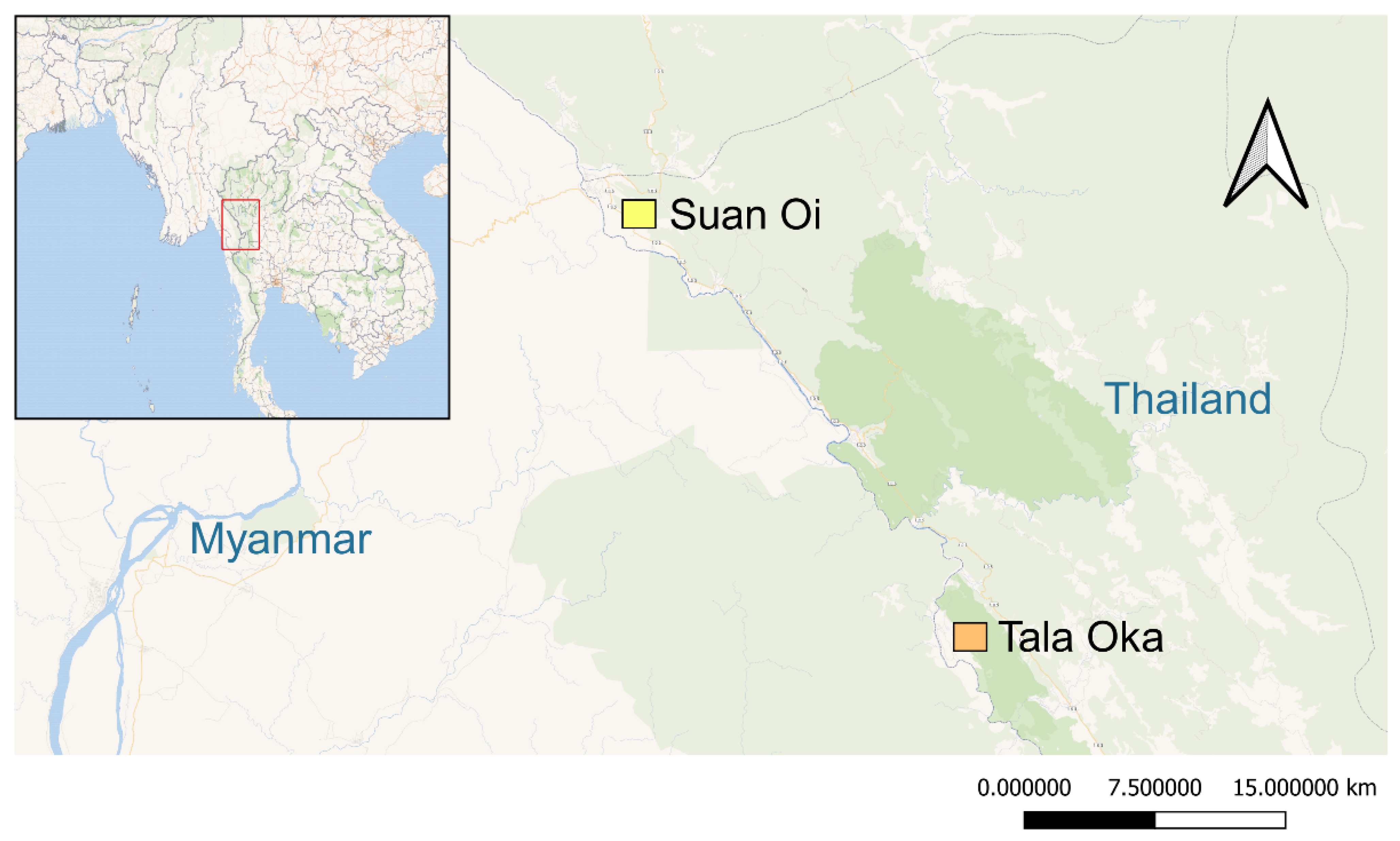
2.1. Study Sites and Mosquito Collections
2.2. Mosquito Collection
2.3. Mosquito Rearing
2.4. Insecticide Susceptibility Test
2.5. DNA Extraction and kdr Detection in Aedes aegypti
2.6. Data Analysis
3. Results
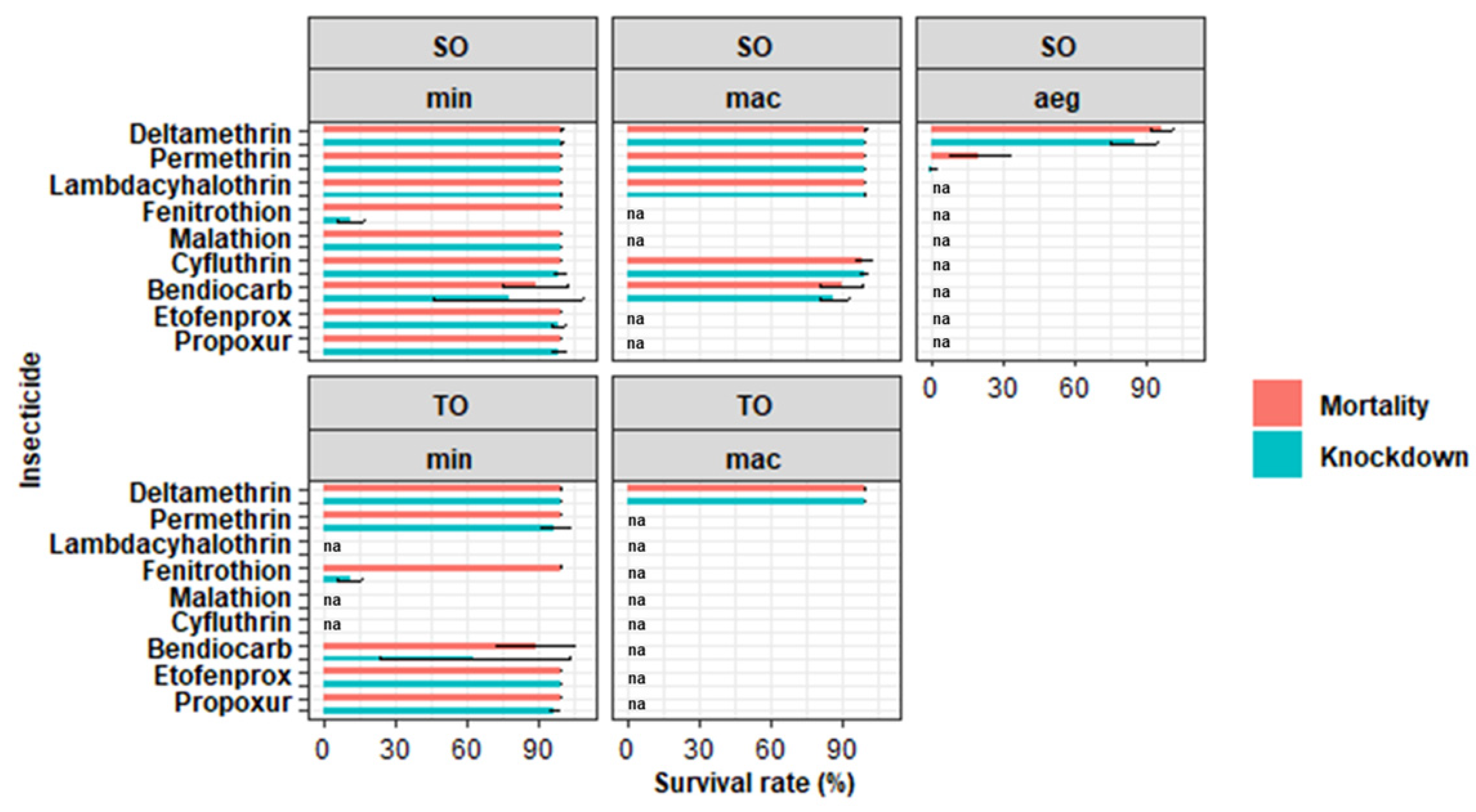
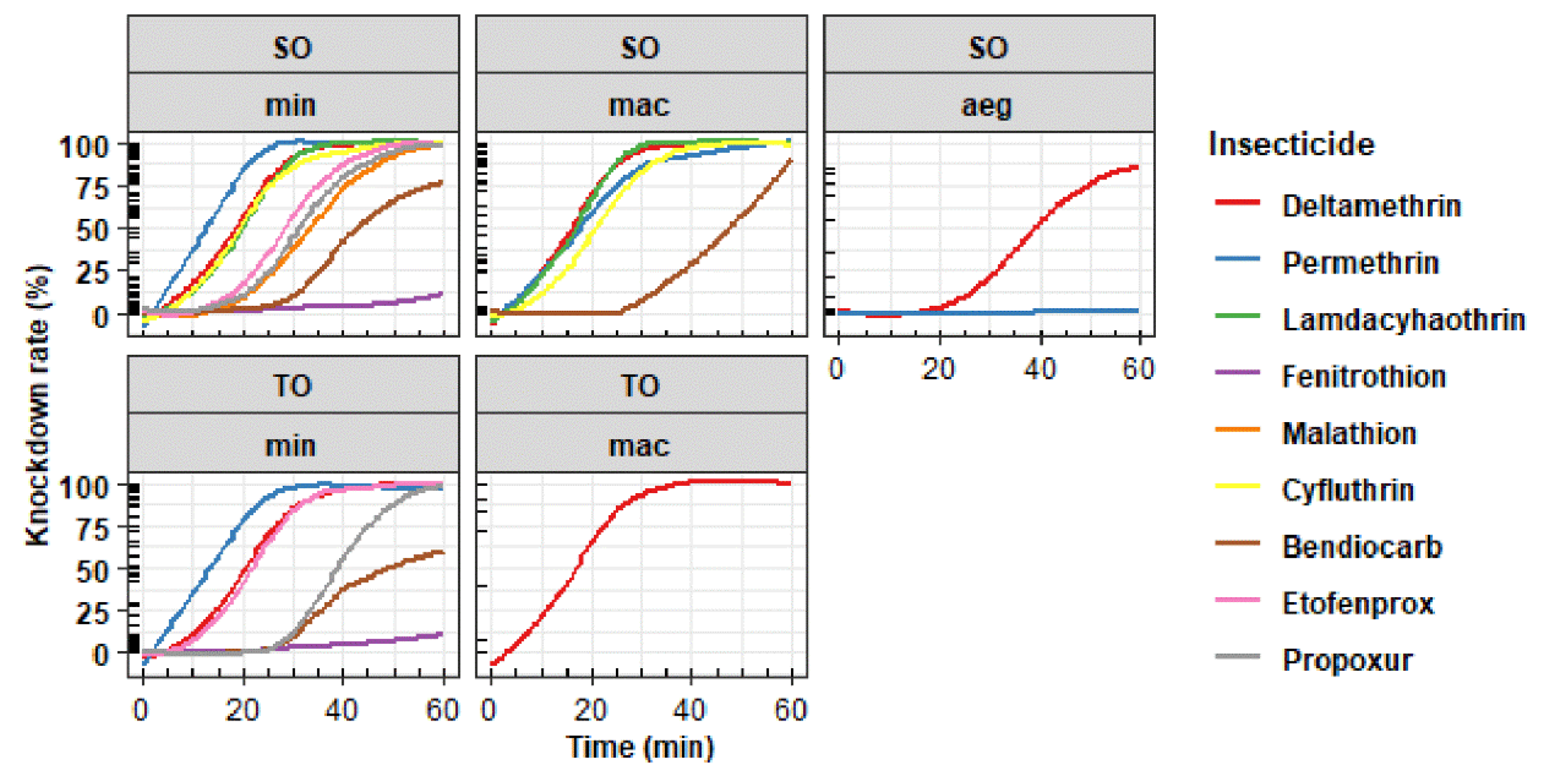
3.1. Bioassays
3.2. Prevalence of kdr Mutations in Ae. aegypti
4. Discussion
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- World Health Organization. World Malaria Report 2020: 20 Years of Global Progress and Challenges; World Health Organization: Geneva, Switzerland, 2020.
- World Health Organization. World Malaria Report 2021; World Health Organization: Geneva, Switzerland, 2021.
- Kobayashi, J.; Phompida, S.; Toma, T.; Looareensuwan, S.; Toma, H.; Miyagi, I. The effectiveness of impregnated bed net in malaria control in Laos. Acta Trop. 2004, 89, 299–308. [Google Scholar] [CrossRef] [PubMed]
- Sochantha, T.; Hewitt, S.; Nguon, C.; Okell, L.; Alexander, N.; Yeung, S.; Vannara, H.; Rowland, M.; Socheat, D. Insecticide-treated bednets for the prevention of Plasmodium falciparum malaria in Cambodia: A cluster-randomized trial. Trop. Med. Int. Health 2006, 11, 1166–1177. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Smithuis, F.M.; Kyaw, M.K.; Phe, U.O.; van der Broek, I.; Katterman, N.; Rogers, C.; Almeida, P.; Kager, P.A.; Stepniewska, K.; Lubell, Y. The effect of insecticide–treated bed nets on the incidence and prevalence of malaria in children in an area of unstable seasonal transmission in western Myanmar. Malar. J. 2013, 12, 363. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Edwards, H.M.; Sriwichai, P.; Kirabittir, K.; Prachumsri, J.; Chavez, I.F.; Hii, J. Transmission risk beyond the village: Entomological and human factors contributing to residual malaria transmission in an area approaching malaria elimination on the Thailand–Myanmar border. Malar. J. 2019, 18, 221. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Corbel, V.; N’Guessan, R. Distribution, mechanisms, impact and management of insecticide resistance in malaria vectors: A pragmatic review. In Anopheles Mosquitoes—New Insights into Malaria Vectors; IntechOpen: London, UK, 2013. [Google Scholar]
- Hougard, J.-M.; Duchon, S.; Darriet, F.; Zaim, M.; Rogier, C.; Guillet, P. Comparative performances, under laboratory conditions, of seven pyrethroid insecticides used for impregnation of mosquito nets. Bull. World Health Organ. 2003, 81, 324–333. [Google Scholar]
- World Health Organization. Global Malaria Plan. Global Plan for Insecticide Resistance Management in Malaria Vectors; WHO: Geneva, Switzerland, 2012.
- Zaim, M.; Aitio, A.; Nakashima, N. Safety of pyrethroid-treated mosquito nets. Med. Vet. Entomol. 2000, 14, 1–5. [Google Scholar] [CrossRef]
- Ranson, H.; N’guessan, R.; Lines, J.; Moiroux, N.; Nkuni, Z.; Corbel, V. Pyrethroid resistance in African anopheline mosquitoes: What are the implications for malaria control? Trends Parasitol. 2011, 27, 91–98. [Google Scholar] [CrossRef]
- Hemingway, J.; Ranson, H. Insecticide resistance in insect vectors of human disease. Annu. Rev. Entomol. 2000, 45, 371–391. [Google Scholar] [CrossRef]
- Scott, J.G. Cytochromes P450 and insecticide resistance. Insect Biochem. Mol. Biol. 1999, 29, 757–777. [Google Scholar] [CrossRef]
- Smith, L.B.; Tyagi, R.; Kasai, S.; Scott, J.G. CYP–mediated permethrin resistance in Aedes aegypti and evidence for trans-regulation. PLoS Negl. Trop. Dis 2018, 12, e0006933. [Google Scholar] [CrossRef] [Green Version]
- Karunaratne, S.H.P.P.; Hemingway, J.; Jayawardena, K.G.I.; Dassanayaka, V.; Vaughan, A. Kinetic and molecular differences in the amplified and non–amplified esterases from insecticide–resistant and susceptible Culex quinquefasciatus mosquitoes. J. Biol. Chem. 1995, 270, 31124–31128. [Google Scholar] [CrossRef] [PubMed]
- Prasad, K.M.; Raghavendra, K.; Verma, V.; Velamuri, P.S.; Pande, V. Esterases are responsible for malathion resistance in Anopheles stephensi: A proof using biochemical and insecticide inhibition studies. J. Vector Borne Dis. 2017, 54, 226. [Google Scholar] [PubMed]
- Ranson, H.; Hemingway, J. Mosquito glutathione transferases. Methods Enzymol. 2005, 401, 226–241. [Google Scholar] [PubMed]
- Liu, N. Insecticide resistance in mosquitoes: Impact, mechanisms, and research directions. Annu. Rev. Entomol. 2015, 60, 537–559. [Google Scholar] [CrossRef]
- Chang, C.; Huang, X.Y.; Chang, P.C.; Wu, H.H.; Dai, S.M. Inheritance and stability of sodium channel mutations associated with permethrin knockdown resistance in Aedes aegypti. Pestic. Biochem. Physiol. 2012, 104, 136–142. [Google Scholar] [CrossRef]
- Assogba, B.S.; Djogbénou, L.S.; Saizonou, J.; Milesi, P.; Djossou, L.; Djegbe, I.; Oumbouke, W.A.; Chandre, F.; Baba–Moussa, L.; Weill, M. Phenotypic effects of concomitant insensitive acetylcholinesterase (ace–1 R) and knockdown resistance (kdr R) in Anopheles gambiae: A hindrance for insecticide resistance management for malaria vector control. Parasit. Vectors 2014, 7, 548. [Google Scholar]
- Taylor-Wells, J.; Brooke, B.D.; Bermudez, I.; Jones, A.K. The neonicotinoid imidacloprid, and the pyrethroid deltamethrin, are antagonists of the insect Rdl GABA receptor. J. Neurochem. 2015, 135, 705–713. [Google Scholar] [CrossRef] [Green Version]
- Ministry of Public Health. The Status of Malaria Cases in Thailand. Available online: http://malaria.ddc.moph.go.th/malariaR10/index_newversion.php (accessed on 20 December 2021).
- Ministry of Public Health. The Status of Dengue and Dengue Haemorrhagic Fever Cases in Thailand. Available online: http://doe.moph.go.th/surdata/index.php (accessed on 20 December 2021).
- Somboon, P.; Prapanthadara, L.-A.; Suwonkerd, W. Insecticide susceptibility tests of Anopheles minimus sl, Aedes aegypti, Aedes albopictus, and Culex quinquefasciatus in northern Thailand. Southeast Asian J. Trop. Med. Public Health 2003, 34, 87–93. [Google Scholar]
- Stenhouse, S.A.; Plernsub, S.; Yanola, J.; Lumjuan, N.; Dantrakool, A.; Choochote, W.; Somboon, P. Detection of the V1016G mutation in the voltage-gated sodium channel gene of Aedes aegypti (Diptera: Culicidae) by allele–specific PCR assay, and its distribution and effect on deltamethrin resistance in Thailand. Parasit. Vectors 2013, 6, 253. [Google Scholar] [CrossRef] [Green Version]
- Thongwat, D.; Bunchu, N. Susceptibility to temephos, permethrin and deltamethrin of Aedes aegypti (Diptera: Culicidae) from Muang district, Phitsanulok Province, Thailand. Asian Pac. J. Trop. Med. 2015, 8, 14–18. [Google Scholar] [CrossRef] [Green Version]
- Plernsub, S.; Saingamsook, J.; Yanola, J.; Lumjuan, N.; Tippawangkosol, P.; Walton, C.; Somboon, P. Temporal frequency of knockdown resistance mutations, F1534C and V1016G, in Aedes aegypti in Chiang Mai city, Thailand and the impact of the mutations on the efficiency of thermal fogging spray with pyrethroids. Acta Trop. 2016, 162, 125–132. [Google Scholar] [CrossRef] [PubMed]
- Saingamsook, J.; Saeung, A.; Yanola, J.; Lumjuan, N.; Walton, C.; Somboon, P. A multiplex PCR for detection of knockdown resistance mutations, V1016G and F1534C, in pyrethroid–resistant Aedes aegypti. Parasit. Vectors 2017, 10, 465. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sathantriphop, S.; Paeporn, P.; Ya-Umphan, P.; Mukkhun, P.; Thanispong, K.; Chansang, C.; Bangs, M.J.; Chareonviriyaphap, T.; Tainchum, K. Behavioral Action of deltamethrin and cypermethrin in pyrethroid–resistant Aedes aegypti (Diptera: Culicidae): Implications for control strategies in Thailand. J. Med. Entomol. 2020, 57, 1157–1167. [Google Scholar] [CrossRef]
- Nachaiwieng, W.; Yanola, J.; Chamnanya, S.; Lumjuan, N.; Somboon, P. Efficacy of five commercial household insecticide aerosol sprays against pyrethroid resistant Aedes aegypti and Culex quinquefasciatus mosquitoes in Thailand. Pestic. Biochem. Physiol. 2021, 178, 104911. [Google Scholar] [CrossRef] [PubMed]
- Hirata, K.; Komagata, O.; Itokawa, K.; Yamamoto, A.; Tomita, T.; Kasai, S. A single crossing–over event in voltage–sensitive Na+ channel genes may cause critical failure of dengue mosquito control by insecticides. PLoS Negl. Trop. Dis. 2014, 8, e3085. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Plernsub, S.; Saingamsook, J.; Yanola, J.; Lumjuan, N.; Tippawangkosol, P.; Sukontason, K.; Walton, C.; Somboon, P. Additive effect of knockdown resistance mutations, S989P, V1016G and F1534C, in a heterozygous genotype conferring pyrethroid resistance in Aedes aegypti in Thailand. Parasit. Vectors 2016, 9, 417. [Google Scholar] [CrossRef] [Green Version]
- Chaumeau, V.; Cerqueira, D.; Zadrozny, J.; Kittiphanakun, P.; Andolina, C.; Chareonviriyaphap, T.; Nosten, F.; Corbel, V. Insecticide resistance in malaria vectors along the Thailand–Myanmar border. Parasit. Vectors 2017, 10, 165. [Google Scholar] [CrossRef] [Green Version]
- Sumarnrote, A.; Overgaard, H.J.; Marasri, N.; Fustec, B.; Thanispong, K.; Chareonviriyaphap, T.; Corbel, V. Status of insecticide resistance in Anopheles mosquitoes in Ubon Ratchathani province, Northeastern Thailand. Malar. J. 2017, 16, 299. [Google Scholar] [CrossRef] [Green Version]
- Stevenson, B.J.; Pignatelli, P.; Nikou, D.; Paine, M.J. Pinpointing P450s associated with pyrethroid metabolism in the dengue vector, Aedes aegypti: Developing new tools to combat insecticide resistance. PLoS Negl. Trop. Dis. 2012, 6, e1595. [Google Scholar] [CrossRef] [Green Version]
- Poupardin, R.; Srisukontarat, W.; Yunta, C.; Ranson, H. Identification of carboxylesterase genes implicated in temephos resistance in the dengue vector Aedes aegypti. PLoS Negl. Trop. Dis. 2014, 8, e2743. [Google Scholar] [CrossRef] [Green Version]
- Rodpradit, P.; Boonsuepsakul, S.; Chareonviriyaphap, T.; Bangs, M.J.; Rongnoparut, P. Cytochrome P450 genes: Molecular cloning and overexpression in a pyrethroid–resistant strain of Anopheles minimus mosquito. J. Am. Mosq. Control Assoc. 2005, 21, 71–79. [Google Scholar] [CrossRef]
- Duangkaew, P.; Kaewpa, D.; Rongnoparut, P. Protective efficacy of Anopheles minimus CYP6P7 and CYP6AA3 against cytotoxicity of pyrethroid insecticides in Spodoptera frugiperda (Sf9) insect cells. Trop. Biomed. 2011, 28, 293–301. [Google Scholar]
- Rattanarithikul, R.; Harrison, B.A.; Panthusiri, P.; Peyton, E.; Coleman, R.E. Illustrated keys to the mosquitoes of Thailand III. Genera Aedeomyia, Ficalbia, Mimomyia, Hodgesia, Coquillettidia, Mansonia, and Uranotaenia. Southeast Asian J. Trop. Med. Public Health 2006, 37, 1. [Google Scholar] [PubMed]
- Choochote, W.; Saeung, A. Systematic techniques for the recognition of Anopheles species complexes. In Anopheles Mosquitoes—New Insights into Malaria Vectors; IntechOpen: London, UK, 2013. [Google Scholar]
- World Health Organization. Test Procedures for Insecticide Resistance Monitoring in Malaria Vector Mosquitoes, 2nd ed.; World Health Organization: Geneva, Switzerland, 2016.
- Sriwichai, P.; Karl, S.; Samung, Y.; Kiattibutr, K.; Sirichaisinthop, J.; Mueller, I.; Cui, L.; Sattabongkot, J. Imported Plasmodium falciparum and locally transmitted Plasmodium vivax: Cross–border malaria transmission scenario in northwestern Thailand. Malar. J. 2017, 16, 258. [Google Scholar] [CrossRef] [PubMed]
- Aïkpon, R.; Agossa, F.; Ossè, R.; Oussou, O.; Aïzoun, N.; Oké-Agbo, F.; Akogbéto, M. Bendiocarb resistance in Anopheles gambiae sl. populations from Atacora department in Benin, West Africa: A threat for malaria vector control. Parasit. Vectors 2013, 6, 192. [Google Scholar] [CrossRef] [Green Version]
- Keïta, M.; Kané, F.; Thiero, O.; Traoré, B.; Zeukeng, F.; Sodio, A.B.; Traoré, S.F.; Djouaka, R.; Doumbia, S.; Sogoba, N. Acetylcholinesterase (ace–1 R) target site mutation G119S and resistance to carbamates in Anopheles gambiae (sensu lato) populations from Mali. Parasit. Vectors 2020, 13, 1–9. [Google Scholar] [CrossRef]
- Kpanou, C.D.; Sagbohan, H.W.; Dagnon, F.; Padonou, G.G.; Ossè, R.; Salako, A.S.; Sidick, A.; Sewadé, W.; Sominahouin, A.; Condo, P. Characterization of resistance profile (intensity and mechanisms) of Anopheles gambiae in three communes of northern Benin, West Africa. Malar. J. 2021, 20, 328. [Google Scholar] [CrossRef]
- Yusuf, M.A.; Vatandoost, H.; Oshaghi, M.A.; Hanafi-Bojd, A.A.; Yayo, A.; Enayati, A.; Abduljalal, A.; Abdullahi, A.S.; Jalo, R.I.; Firdausi, A. Biochemical mechanism of insecticide resistance in malaria vector, Anopheles gambiae s.l in Nigeria. Iran. J. Public Health 2021, 50, 101–110. [Google Scholar] [CrossRef]
- Edi, C.V.; Djogbénou, L.; Jenkins, A.M.; Regna, K.; Muskavitch, M.A.T.; Poupardin, R.; Jones, C.M.; Essandoh, J.; Kétoh, G.K.; Paine, M.J.I.; et al. CYP6 P450 enzymes and ACE–1 duplication produce extreme and multiple insecticide resistance in the malaria mosquito Anopheles gambiae. PLoS Genet. 2014, 10, e1004236. [Google Scholar] [CrossRef]
- Saha, P.; Chatterjee, M.; Ballav, S.; Chowdhury, A.; Basu, N.; Maji, A.K. Prevalence of kdr mutations and insecticide susceptibility among natural population of Aedes aegypti in West Bengal. PLoS ONE 2019, 14, e0215541. [Google Scholar] [CrossRef]
- Pareja-Loaiza, P.X.; Santacoloma Varon, L.; Rey Vega, G.; Gómez–Camargo, D.; Maestre-Serrano, R.; Lenhart, A. Mechanisms associated with pyrethroid resistance in populations of Aedes aegypti (Diptera: Culicidae) from the Caribbean coast of Colombia. PLoS ONE 2020, 15, e0228695. [Google Scholar] [CrossRef] [PubMed]
- Yanola, J.; Somboon, P.; Walton, C.; Nachaiwieng, W.; Prapanthadara, L.-A. A novel F1552/C1552 point mutation in the Aedes aegypti voltage–gated sodium channel gene associated with permethrin resistance. Pestic. Biochem. Physiol. 2010, 96, 127–131. [Google Scholar] [CrossRef]
- Endersby-Harshman, N.M.; Ali, A.; Alhumrani, B.; Alkuriji, M.A.; Al-Fageeh, M.B.; Al-Malik, A.; Alsuabeyl, M.S.; Elfekih, S.; Hoffmann, A.A. Voltage-sensitive sodium channel (Vssc) mutations associated with pyrethroid insecticide resistance in Aedes aegypti (L.) from Jeddah, Kingdom of Saudi Arabia-baseline information for a Wolbachia release program. Parasit. Vectors 2021, 14, 361. [Google Scholar] [CrossRef] [PubMed]
- Ahmad, N.A.; Endersby-Harshman, N.M.; Mohd Mazni, N.R.; Mohd Zabari, N.Z.A.; Amran, S.N.S.; Ridhuan Ghazali, M.K.; Abdul Karim, M.A.; Cheong, Y.L.; Sinkins, S.P.; Ahmad, N.W. Characterization of sodium channel mutations in the dengue vector mosquitoes Aedes aegypti and Aedes albopictus within the context of ongoing Wolbachia releases in Kuala Lumpur, Malaysia. Insects 2020, 11, 529. [Google Scholar] [CrossRef]
- Sene, N.M.; Mavridis, K.; Ndiaye, E.H.; Diagne, C.T.; Gaye, A.; Ngom, E.H.M.; Ba, Y.; Diallo, D.; Vontas, J.; Dia, I.; et al. Insecticide resistance status and mechanisms in Aedes aegypti populations from Senegal. PLoS Negl. Trop. Dis. 2021, 15, e0009393. [Google Scholar] [CrossRef] [PubMed]
- Seck, M.C.; Thwing, J.; Fall, F.B.; Gomis, J.F.; Deme, A.; Ndiaye, Y.D.; Daniels, R.; Volkman, S.K.; Ndiop, M.; Ba, M.; et al. Malaria prevalence, prevention and treatment seeking practices among nomadic pastoralists in northern Senegal. Malar. J. 2017, 16, 413. [Google Scholar] [CrossRef]
- Verhaeghen, K.; Van Bortel, W.; Trung, H.D.; Sochantha, T.; Coosemans, M. Absence of knockdown resistance suggests metabolic resistance in the main malaria vectors of the Mekong region. Malar. J. 2009, 8, 84. [Google Scholar] [CrossRef] [Green Version]
- Bonizzoni, M.; Afrane, Y.; Dunn, W.A.; Atieli, F.K.; Zhou, G.; Zhong, D.; Li, J.; Githeko, A.; Yan, G. Comparative transcriptome analyses of deltamethrin-resistant and -susceptible Anopheles gambiae mosquitoes from Kenya by RNA-Seq. PLoS ONE 2012, 7, e44607. [Google Scholar] [CrossRef] [Green Version]
- Xu, J.; Su, X.; Bonizzoni, M.; Zhong, D.; Li, Y.; Zhou, G.; Nguyen, H.; Tong, S.; Yan, G.; Chen, X.-G. Comparative transcriptome analysis and RNA interference reveal CYP6A8 and SNPs related to pyrethroid resistance in Aedes albopictus. PLoS Negl. Trop. Dis. 2018, 12, e0006828. [Google Scholar] [CrossRef]




{kind=link}
{kind=link}
{kind=link}
| Species | Insecticide | N 1 | R 2 | % Mortality | % Knockdown Rate | KDT50 3 (min) | Status 4 |
|---|---|---|---|---|---|---|---|
| An. minimus s.l. | 0.05% Deltamethrin | 1562 | 65 | 99.93 (99.80–100.07) | 99.93 (99.80–100.07) | 19.12 (14.45–23.55) | S |
| 0.75% Permethrin | 96 | 4 | 100 (na) | 100 (na) | 13.72 (11.18–16.17) | S | |
| 0.05% Lambdacyhalothrin | 293 | 12 | 100 (na) | 100 (na) | 20.12 (18.16–20.06) | S | |
| 1.0% Fenitrothion | 195 | 8 | 100 (na) | 11.25 (6.34–16.17) | 105.47 (88.66–140.01) | S | |
| 5.0% Malathion | 50 | 2 | 100 (na) | 100 (na) | 34.10 (32.14–36.05) | S | |
| 1.5% Cyfluthrin | 75 | 3 | 100 (na) | 98.67 (92.93–104.40) | 21.47 (8.44–32.01) | S | |
| 0.1% Bendiocarb | 125 | 5 | 88.80 (71.81–105.79) | 77.60 (38.87–116.33) | 45.63 (43.36–48.14) | R | |
| 0.5% Etofenprox | 246 | 10 | 100 (na) | 98.80 (96.87–100.73) | 29.22 (28.18–30.26) | S | |
| 0.1% Propoxur | 444 | 18 | 100 (na) | 98.89 (97.56–100.22) | 31.72 (30.61–32.83) | S | |
| An. maculatus s.l. | 0.05% Deltamethrin | 809 | 34 | 99.87 (99.61–100.13) | 100 (na) | 17.16 (7.80–25.23) | S |
| 0.75% Permethrin | 100 | 4 | 100 (na) | 100 (na) | 19.77 (16.87–22.47) | S | |
| 0.05% Lambdacyhalothrin | 50 | 2 | 100 (na) | 100 (na) | 17.17 (16.36–17.97) | S | |
| 1.5% Cyfluthrin | 129 | 6 | 98.66 (95.24–102.09) | 99.33 (97.62–101.05) | 22.17 (17.48–26.48) | S | |
| 0.1% Bendiocarb | 66 | 3 | 89.92 (68.40–111.43) | 86.50 (71.44–101.56) | 46.95 (44.76–49.37) | R | |
| Ae. aegypti | 0.05% Deltamethrin | 225 | 9 | 96.89 (93.19–100.58) | 85.33 (77.65–93.02) | 41.09 (39.87–42.33) | R* |
| 0.75% Permethrin | 200 | 8 | 20 (9.43–30.57) | 0.5 (–0.68–1.68) | 144.33 (na) | R |
| Species | Insecticide | N 1 | R 2 | % Mortality | % Knockdown Rate | KDT50 3 (min) | Status 4 |
|---|---|---|---|---|---|---|---|
| An. minimus s.l. | 0.05% Deltamethrin | 575 | 25 | 100 (na) | 100 (na) | 21.96 (18.96–24.83) | S |
| 0.75% Permethrin | 100 | 4 | 100 (na) | 97 (87.45–106.55) | 15.44 (–14.67–28.78) | S | |
| 1.0% Fenitrothion | 100 | 4 | 100 (na) | 11 (2.99–19.01) | 104.69 (88.37–137.32) | S | |
| 0.1% Bendiocarb | 125 | 6 | 88.77 (71.88–105.66) | 63.31 (21.18–105.44) | 50.81 (47.60–54.95) | R | |
| 0.5% Etofenprox | 50 | 2 | 100 (na) | 100 (na) | 22.93 (19.59–26.14) | S | |
| 0.1% Propoxur | 99 | 4 | 100 (na) | 96.96 (93.73–100.19) | 39.56 (37.52–41.61) | S | |
| An. maculatus s.l. | 0.05% Deltamethrin | 50 | 2 | 100 (na) | 100 (na) | 18.01 (16.59–19.40) | S |
| Insecticide | Status 1 | Total PCR | Kdr Genotype | G Allele Frequency (95% CI) | C Allele Frequency (95% CI) | ||
|---|---|---|---|---|---|---|---|
| VV/CC | VG/FC | GG/FF | |||||
| 0.05% Deltamethrin | R | 7 | 0 | 4 | 3 | 0.714 (0.454–0.883) | 0.286 (0.117–0.546) |
| S | 30 | 22 | 6 | 2 | 0.167 (0.093–0.280) | 0.833 (0.720–0.907) | |
| 0.75% Permethrin | R | 30 | 22 | 7 | 1 | 0.150 (0.081–0.261) | 0.850 (0.739–0.919) |
| S | 30 | 24 | 6 | 0 | 0.100 (0.047–0.201) | 0.900 (0.799–0.953) | |
| Mutant Allele | Insecticide | Odd Ratio (95% CI) | Fisher’s Exact Test |
|---|---|---|---|
| 1016G | 0.05% Deltamethrin | 12.500 (3.489–39.980) | 0.0001 * |
| 0.75% Permethrin | 1.588 (0.534–4.904) | 1.588 | |
| 1534C | 0.05% Deltamethrin | 0.080 (0.025–0.287) | 0.0001 * |
| 0.75% Permethrin | 0.630 (0.204–1.873) | 0.583 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Pusawang, K.; Sattabongkot, J.; Saingamsook, J.; Zhong, D.; Yan, G.; Somboon, P.; Wongpalee, S.P.; Cui, L.; Saeung, A.; Sriwichai, P. Insecticide Susceptibility Status of Anopheles and Aedes Mosquitoes in Malaria and Dengue Endemic Areas, Thai–Myanmar Border. Insects 2022, 13, 1035. https://doi.org/10.3390/insects13111035
Pusawang K, Sattabongkot J, Saingamsook J, Zhong D, Yan G, Somboon P, Wongpalee SP, Cui L, Saeung A, Sriwichai P. Insecticide Susceptibility Status of Anopheles and Aedes Mosquitoes in Malaria and Dengue Endemic Areas, Thai–Myanmar Border. Insects. 2022; 13(11):1035. https://doi.org/10.3390/insects13111035
Chicago/Turabian StylePusawang, Kanchon, Jetsumon Sattabongkot, Jassada Saingamsook, Daibin Zhong, Guiyun Yan, Pradya Somboon, Somsakul Pop Wongpalee, Liwang Cui, Atiporn Saeung, and Patchara Sriwichai. 2022. "Insecticide Susceptibility Status of Anopheles and Aedes Mosquitoes in Malaria and Dengue Endemic Areas, Thai–Myanmar Border" Insects 13, no. 11: 1035. https://doi.org/10.3390/insects13111035