
New Synonym and New Species Record of Filchneria (Plecoptera: Perlodidae) from China with a Morphological, Phylogenetic and Biogeographic Study on This Genus
 , and
, and 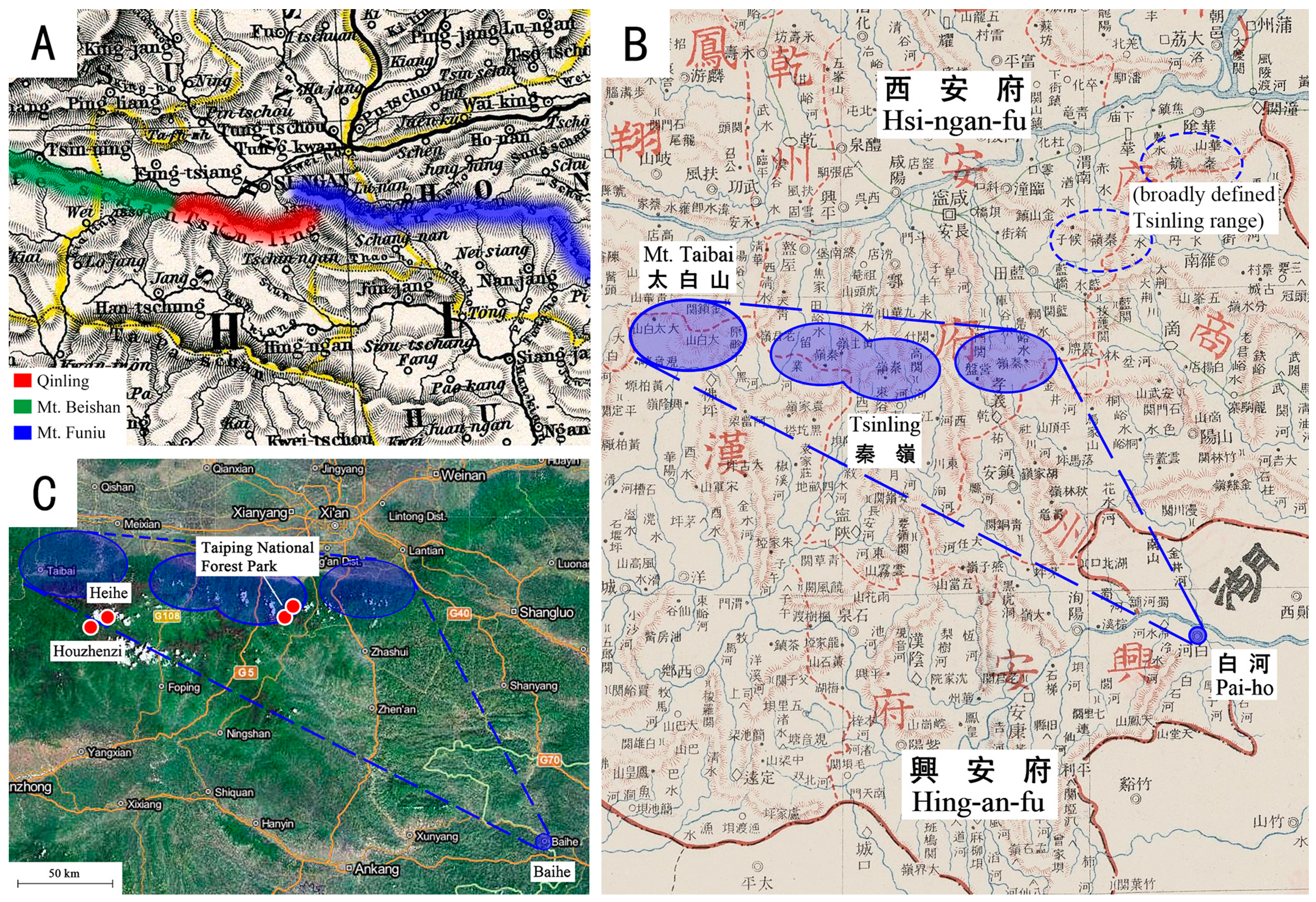{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:Simple Summary
Abstract
1. Introduction
2. Materials and Methods
2.1. Sampling and Identification
2.2. Preparation of Molecular Data
3. Morphological Taxonomy
3.1. Genus Filchneria Klapálek, 1908
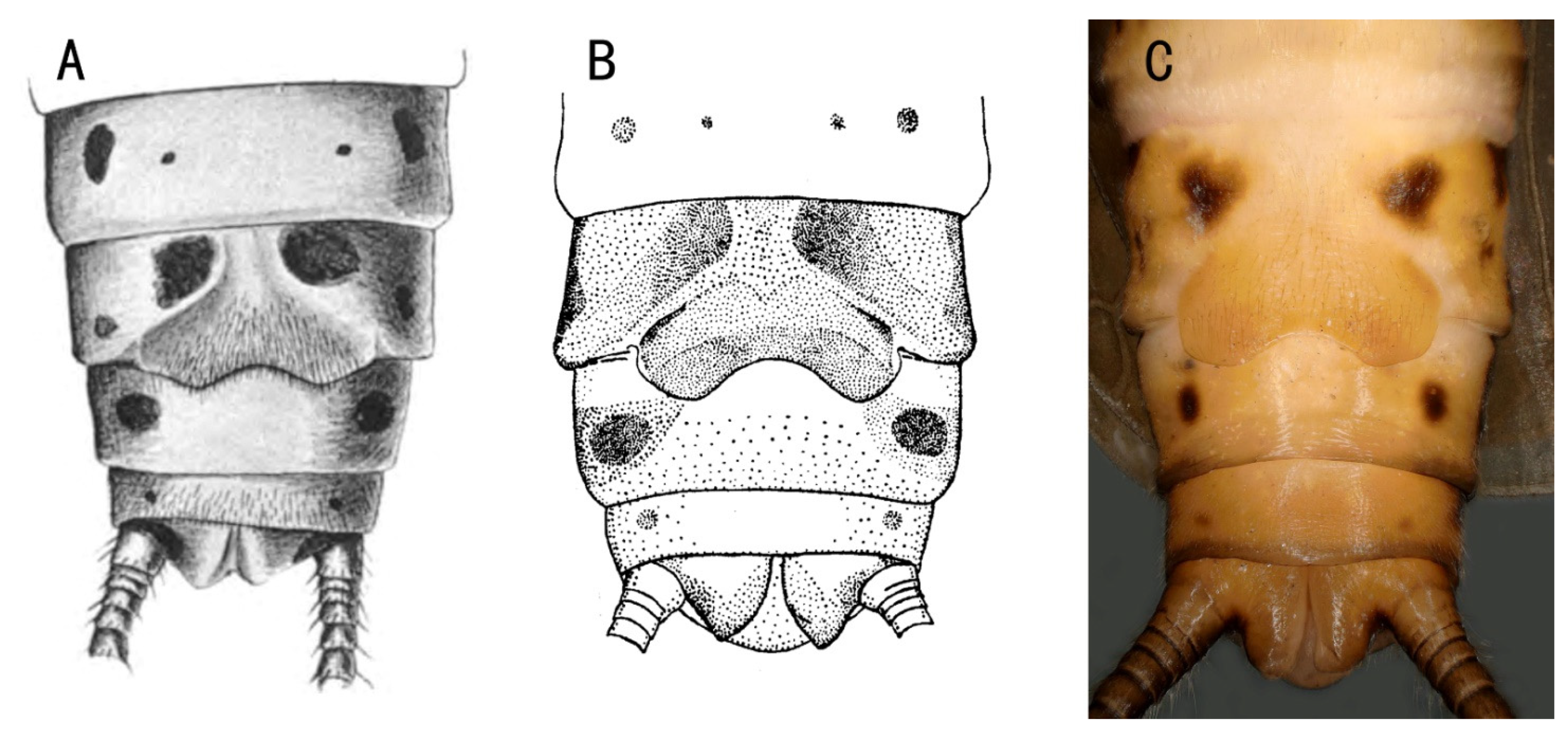
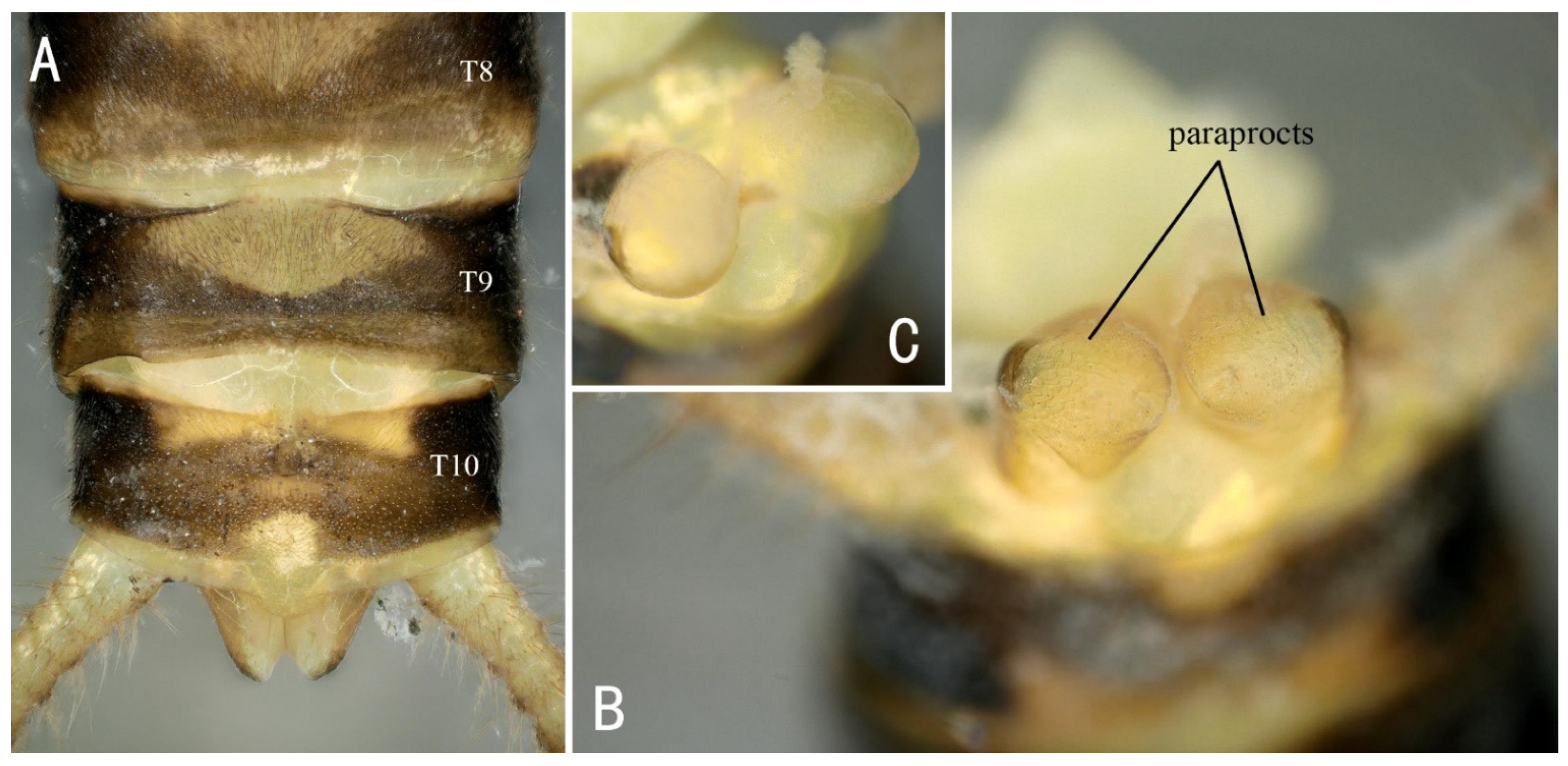
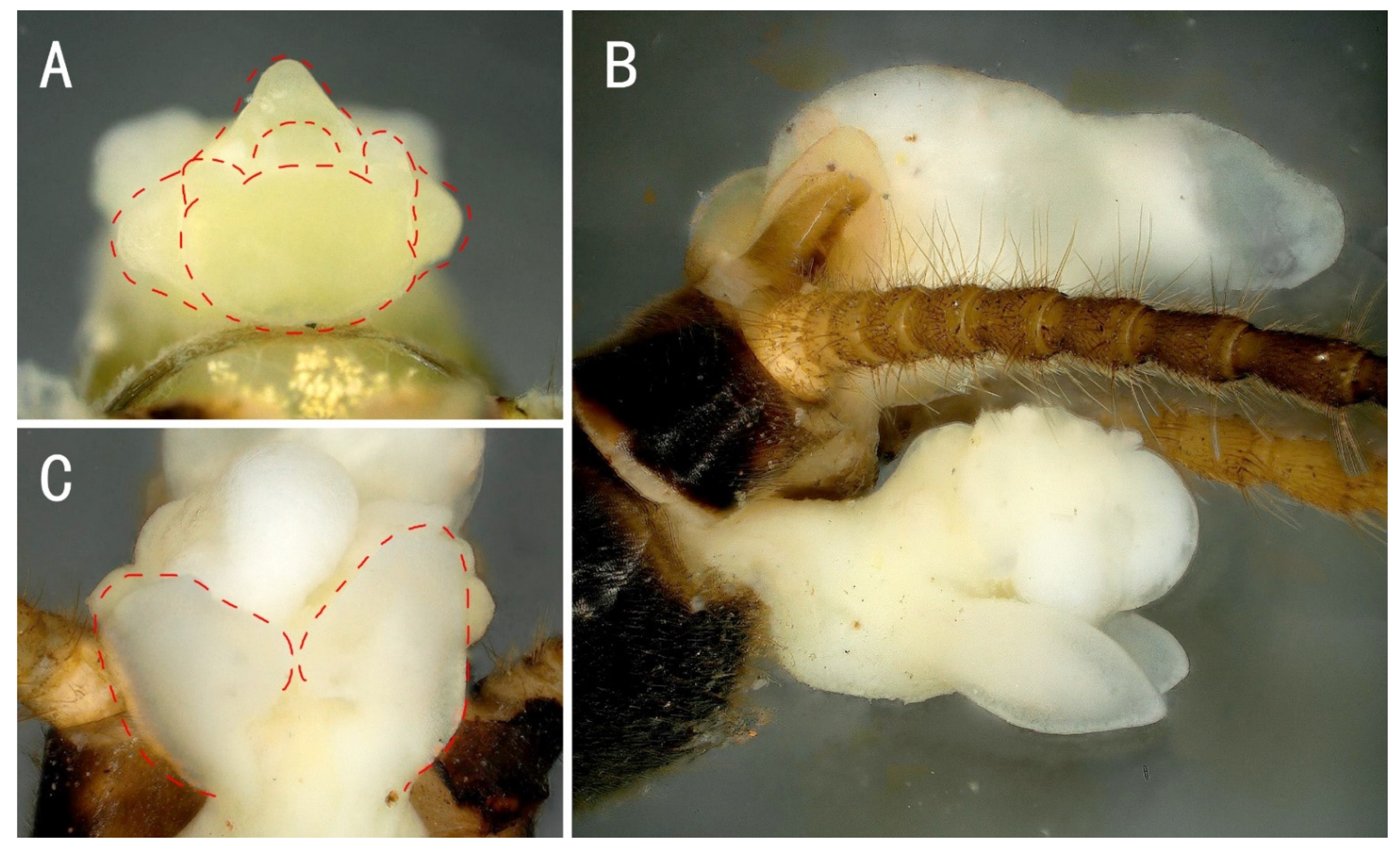
3.2. Filchneria mongolica (Klapálek, 1901) (New Record in China)
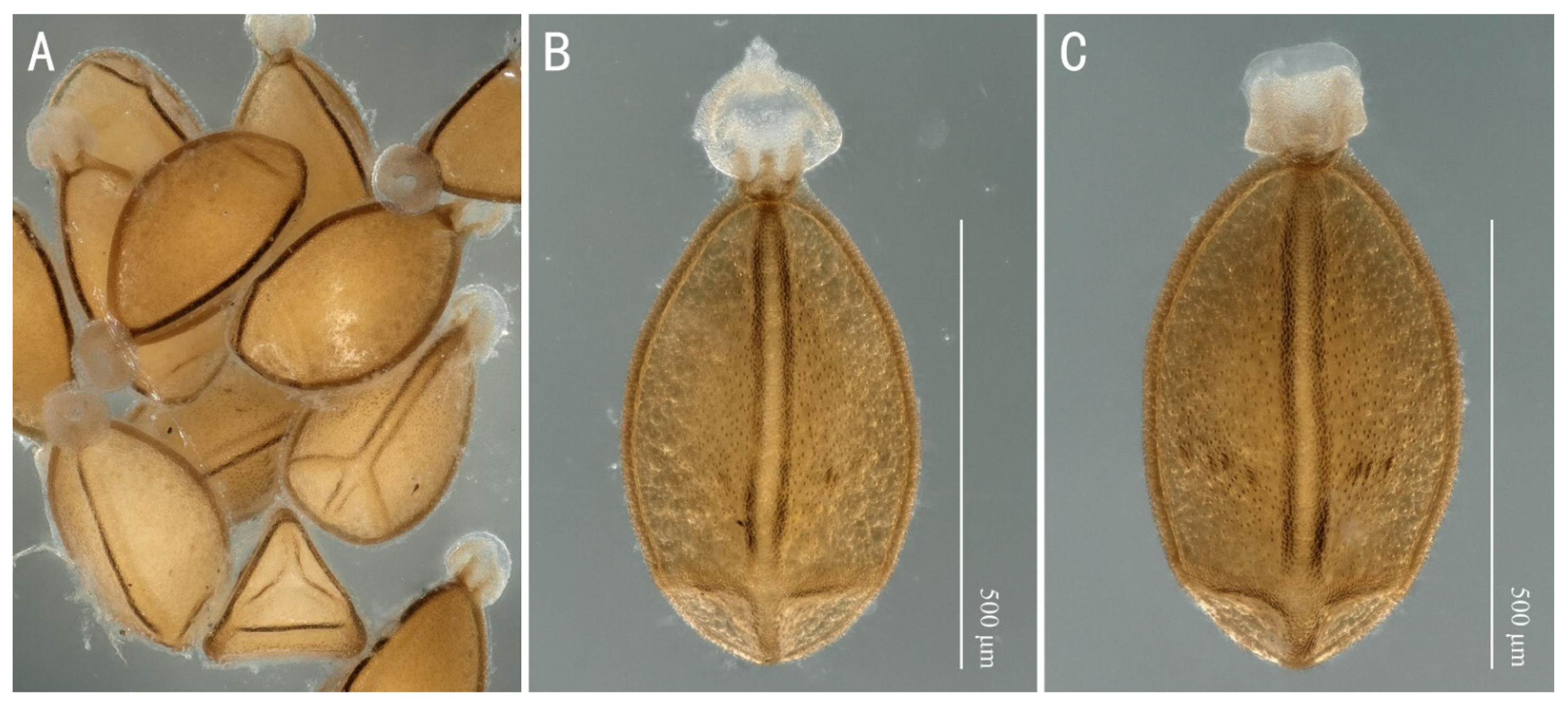
3.3. Filchneria songi Chen, 2019
3.4. Filchneria zhouchangfai (Chen, 2020) (comb. nov.)
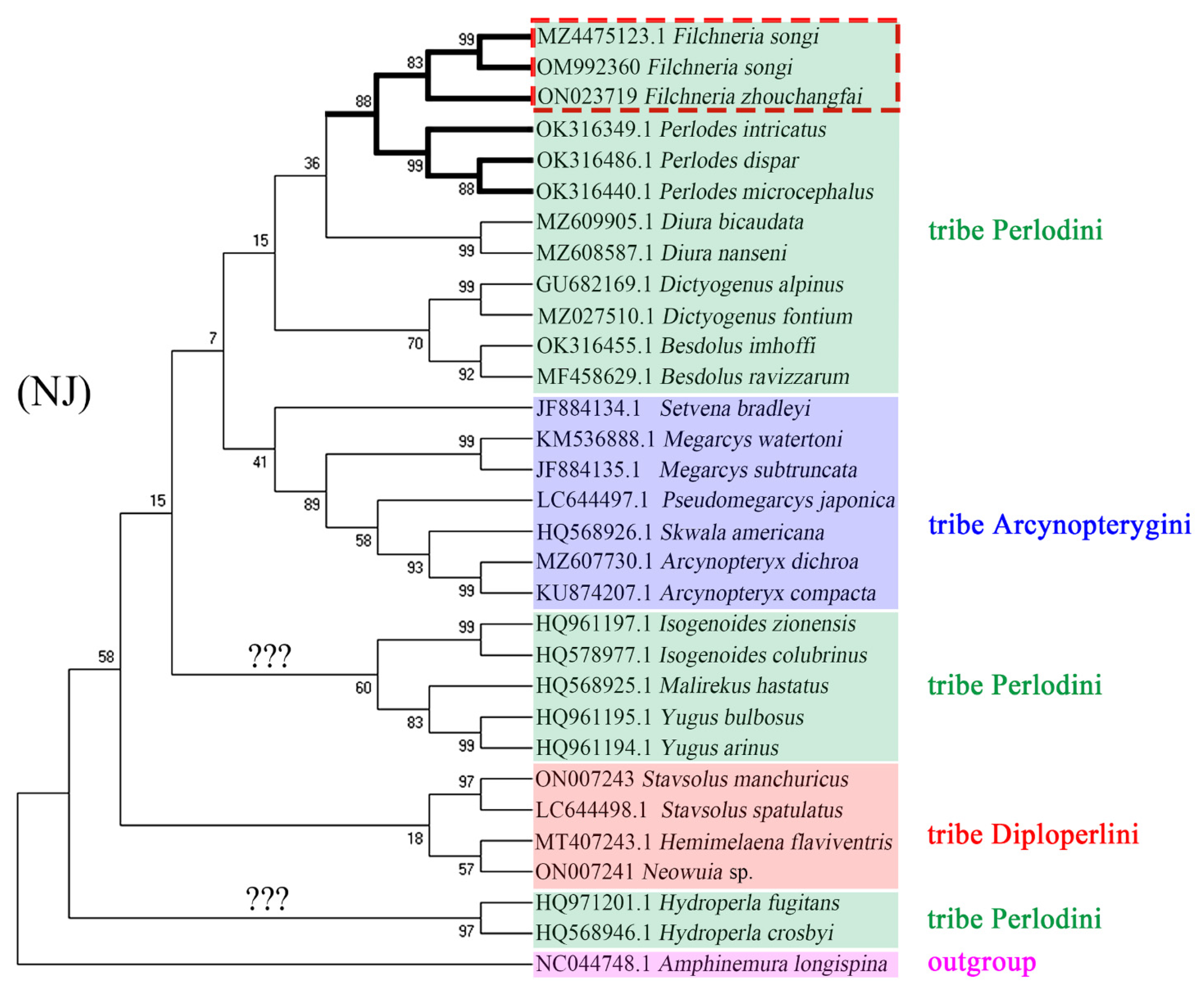
4. Phylogenetic Analysis
5. Discussion
5.1. Morphology
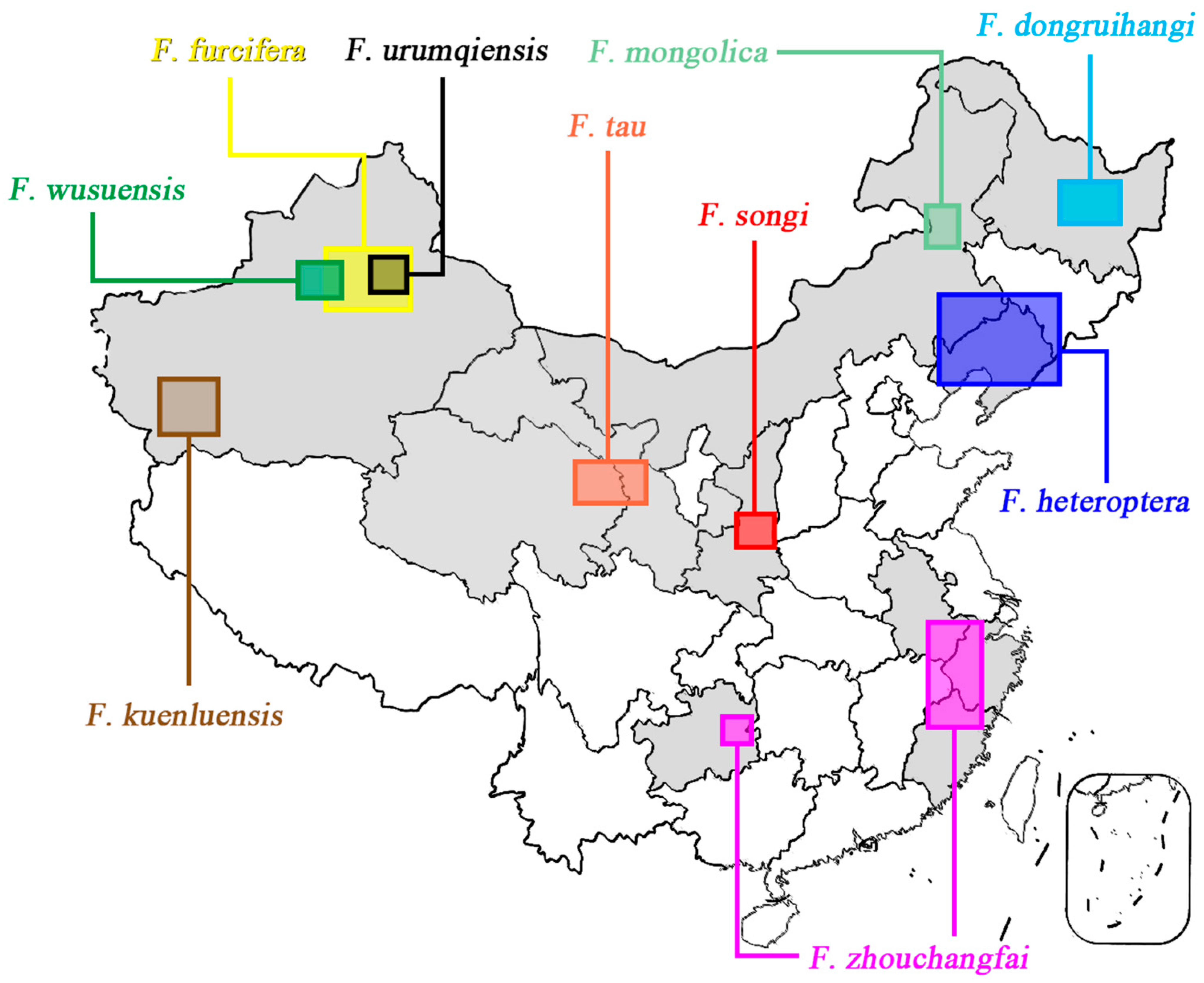
5.2. Biogeography
Author Contributions
Funding
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Klapálek, F. Onovych a málo známych druzích palaearktickỷch Neuropteroid. Rozpravy České Akademie Cís Františka Josefa R 1901, 10, 1–19. [Google Scholar]
- Klapálek, F. Plecoptera. Filchner Expedition China Tibet. Wiss. Zool.-Bot. Ergeb. 1908, 10, 59–64. [Google Scholar]
- Klapálek, F. Plécoptères I. Fam. Perlodidae. Collect. Zool. Baron Edm. Sélys-Longchamps Brux. 1912, 4, 1–66. [Google Scholar]
- Wu, C.F. Plecopterorum Sinensium: A Monograph of Stoneflies of China (Order Plecoptera); Yenching University: Beijing, China, 1938; 225p. [Google Scholar]
- Teslenko, V.A.; Bazova, N.V. On the Stonefly (Plecoptera) Fauna of the Transfrontier Selenga River Basin. Entomol. Rev. 2009, 89, 1059–1068. [Google Scholar] [CrossRef]
- Teslenko, V.A.; Zhiltzova, L.A. Key to the Stoneflies (Insecta, Plecoptera) of Russia and Adjacent Countries. Imagines and Nymphs; Russian Academy of Sciences, Far Eastern Branch, Dalnauka: Vladivostok, Russia, 2009; 381p. [Google Scholar]
- Judson, S.W.; Nelson, C.R. A guide to Mongolian stoneflies (Insecta: Plecoptera). Zootaxa 2012, 3541, 1–118. [Google Scholar] [CrossRef]
- Chen, Z.T. A new species and a new record of Perlodidae (Plecoptera) from China. Zootaxa 2019, 4623, 189–200. [Google Scholar] [CrossRef]
- DeWalt, R.E.; Maehr, M.D.; Neu-Becker, U.; Stueber, G. Plecoptera Species File Online. Available online: http://plecoptera.speciesfile.org (accessed on 30 April 2022).
- Du, Y.Z. A Taxonomic Study on Plecoptera from China. Unpublished. Ph.D. Dissertation, Zhejiang University, Hangzhou, China, 1999; 324p. [Google Scholar]
- Von Salzmann, E. Im Sattel Durch Zentralasien: 6000 Kilometer in 176 Tagen; Walter De Gruyter Incorporated: Berlin, Germany, 1903; 322p. [Google Scholar]
- The Commercial Press. 大清帝国全图 [Map of Tai-Ching-Ti-Kuo]; The Commercial Press: Shanghai, China, 1905; 33p. [Google Scholar]
- Teslenko, V.A.; Zwick, P.; Bazova, N.V. Redescription of Filchneria mongolica (Klapálek, 1901) (Plecoptera, Perlodidae) based on type eggs and fresh material from the Selenga and Amur River Basins of Russia and Mongolia. Zootaxa 2010, 2693, 49–59. [Google Scholar] [CrossRef]
- Chen, Z.T. Sinoperlodes, a new genus of subfamily Perlodinae (Plecoptera, Perlodidae) from coastal southeastern China. Zootaxa 2020, 4779, 584–594. [Google Scholar] [CrossRef]
- Chen, Z.T.; Du, Y.Z. The first two mitogenome from Taeniopterygidae (Insecta: Plecoptera): Structural features and phylogenetic implications. Int. J. Biol. Macromol. 2018, 111, 70–76. [Google Scholar] [CrossRef]
- Zhao, M.Y.; Huo, Q.B.; Du, Y.Z. Molecular phylogeny inferred from the mitogenome of Plecoptera with Oyamia nigribasis (Plecoptera: Perlidae). Sci. Rep. 2020, 10, 20955. [Google Scholar] [CrossRef]
- Xiang, Y.N.; Zhao, M.Y.; Huo, Q.B.; Du, Y.Z. Mitogenome of the Genus Claassenia (Plecoptera: Perlidae) and Phylogenetic Assignment to Subfamily Perlinae. Genes 2021, 12, 1986. [Google Scholar] [CrossRef]
- Ronquist, F.; Huelsenbeck, J.P. MrBayes 3: Bayesian phylogenetic inference under mixed models. Bioinformatics 2003, 19, 1572–1574. [Google Scholar] [CrossRef] [Green Version]
- Nguyen, L.T.; Schmidt, H.A.; Haeseler, A.V.; Minh, B.Q. IQ-TREE: A fast and effective stochastic algorithm for estimating maximum likelihood phylogenies. Mol. Biol. Evol. 2015, 32, 268–274. [Google Scholar] [CrossRef]
- Raušer, J. Plecoptera. Ergebnisse der zoologischen Forschungen von Dr. Z. Kaszab in der Mongolei. Entomol. Abh. Staatl. Mus. Tierkd. Dresd. 1968, 34, 329–398. [Google Scholar]
- Zhiltzova, L.A. The genus Filchneria Klap. and its position in the family Perlodidae (Plecoptera). Zool. J. 1971, 50, 1034–1040. [Google Scholar]
- Zwick, P. Rauserella, a new genus of Plecoptera (Perlodidae), with notes on related genera. In Ephemeroptera & Plecoptera. Biology-Ecology-Systematics; Landolt, P., Sartori, M., Eds.; MTL-Mauron+Tinguely & Lachat, S.A.: Fribourg, Switzerland, 1997; pp. 489–496. [Google Scholar]
- Huo, Q.B. Faunal Investigation of Plecoptera in Jiangsu, China, with the Taxonomy of Chinese Roachlike Stonefly (Plecoptera: Peltoperlidae). Unpublished Master’s Dissertation, Yangzhou University, Yangzhou, China, 2019; 77p. [Google Scholar]
- Chen, Z.T. A new brachypterous Filchneria and two new generic records for China, Arcynopteryx and Pictetiella (Plecoptera: Perlodidae). Zootaxa 2019, 4679, 511–526. [Google Scholar] [CrossRef]
- Chen, Z.T.; Ma, Y.S. Description of a new brachypterous species of Filchneria Klapálek, 1908 (Plecoptera: Perlodidae) from northwestern China. Aquat. Insects 2021, 43, 18–29. [Google Scholar] [CrossRef]
- Teslenko, V.A. A new species of Megaperlodes Yokoyama et al. 1990 (Plecoptera, Perlodidae) from the South of the Russian Far East. Zootaxa 2015, 3904, 553–562. [Google Scholar] [CrossRef]
- Kovács, T.; Murányi, D. Data to the distribution of genus Perlodes in Hungary (Plecoptera: Perlodidae). Folia Hist. Nat. Musei Matra. 2008, 32, 115–118. [Google Scholar]
- Vinçon, G.; Dia, A.; Kovács, T.; Murányi, D. A new stonefly from Lebanon, Perlodes thomasi sp. n. (Plecoptera: Perlodidae). Illiesia 2013, 9, 18–27. [Google Scholar]
- Kovács, T.; Vinçon, G.; Murányi, D.; Sivec, I. A new Perlodes species and its subspecies from the Balkan Peninsula (Plecoptera: Perlodidae). Illiesia 2012, 8, 182–192. [Google Scholar]
- Ma, S.J. 中国昆虫生态地理概述 [Ecological Geography of Chinese Insects]; Science Press: Beijing, China, 1959; 109p. [Google Scholar]
- Shen, X.C. Insect Geography of China; Henan Science and Technology Press: Zhengzhou, China, 2015; 980p. [Google Scholar]
- Du, Y.Z.; He, J.H.; Ma, Y. A preliminary study on the distribution of the family Perlidae (Plecoptera: Perloidea). Zool. Res. 1999, 20, 189–195. [Google Scholar]


















Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Huo, Q.-B.; Zhu, B.-Q.; Rehman, A.; Murányi, D.; Du, Y.-Z.; Wu, J. New Synonym and New Species Record of Filchneria (Plecoptera: Perlodidae) from China with a Morphological, Phylogenetic and Biogeographic Study on This Genus. Insects 2022, 13, 1044. https://doi.org/10.3390/insects13111044
Huo Q-B, Zhu B-Q, Rehman A, Murányi D, Du Y-Z, Wu J. New Synonym and New Species Record of Filchneria (Plecoptera: Perlodidae) from China with a Morphological, Phylogenetic and Biogeographic Study on This Genus. Insects. 2022; 13(11):1044. https://doi.org/10.3390/insects13111044
Chicago/Turabian StyleHuo, Qing-Bo, Bin-Qing Zhu, Abdur Rehman, Dávid Murányi, Yu-Zhou Du, and Jun Wu. 2022. "New Synonym and New Species Record of Filchneria (Plecoptera: Perlodidae) from China with a Morphological, Phylogenetic and Biogeographic Study on This Genus" Insects 13, no. 11: 1044. https://doi.org/10.3390/insects13111044