

Tephritid Fruit Fly Species Composition, Seasonality, and Fruit Infestations in Two Central African Agro-Ecological Zones

 , , , , ,
, , , , ,
Abstract
:Simple Summary
Abstract
1. Introduction
2. Materials and Methods
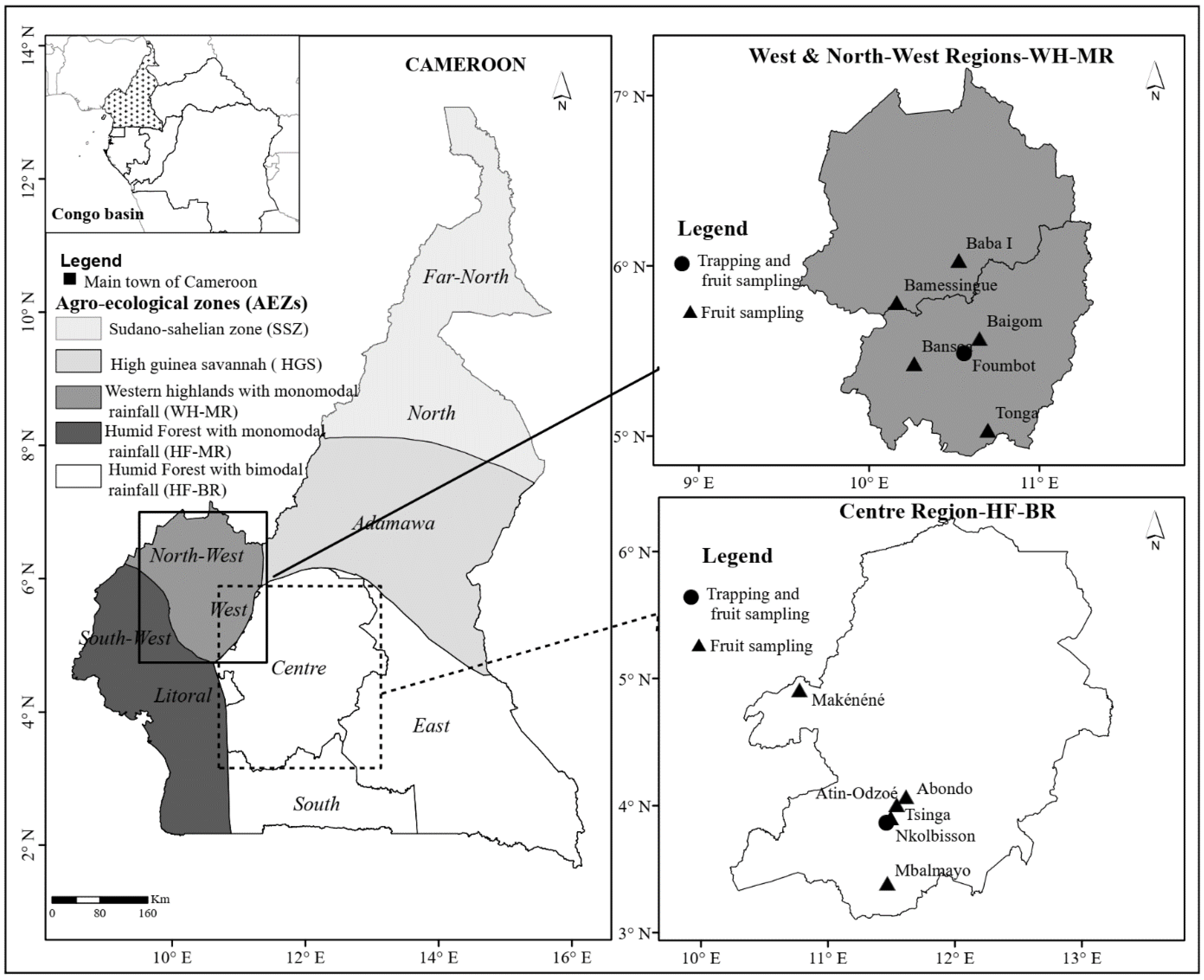
2.1. Study Sites
2.2. Male Lures and Food Baits Traps
2.3. Host Fruit Collection and Handling
2.4. Statistical Analysis
3. Results
3.1. Fruit Fly Diversity and Richness in Traps
3.2. Abundance of the Main Fruit Flies in Traps
3.3. Comparisons of Food Baits
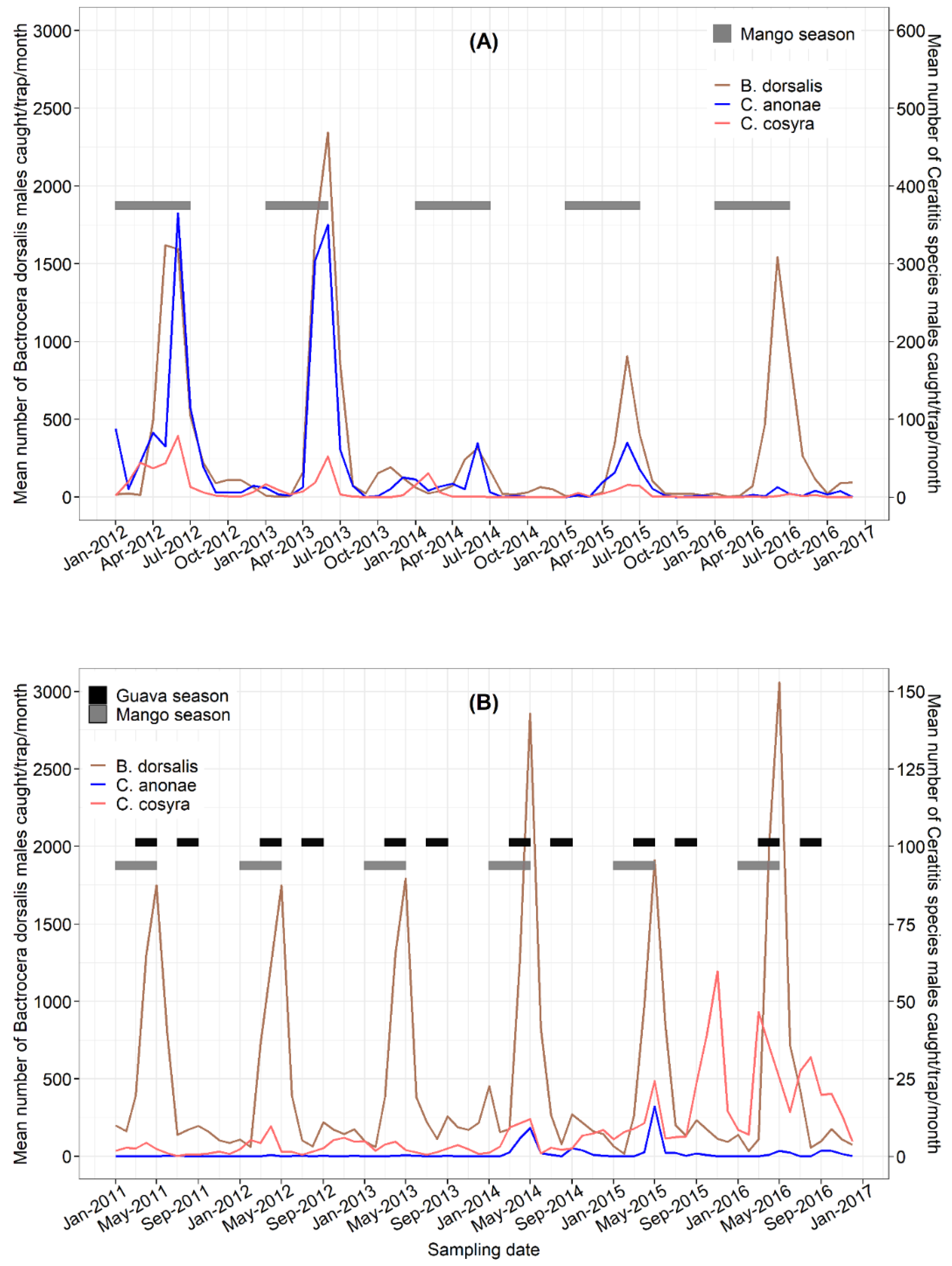
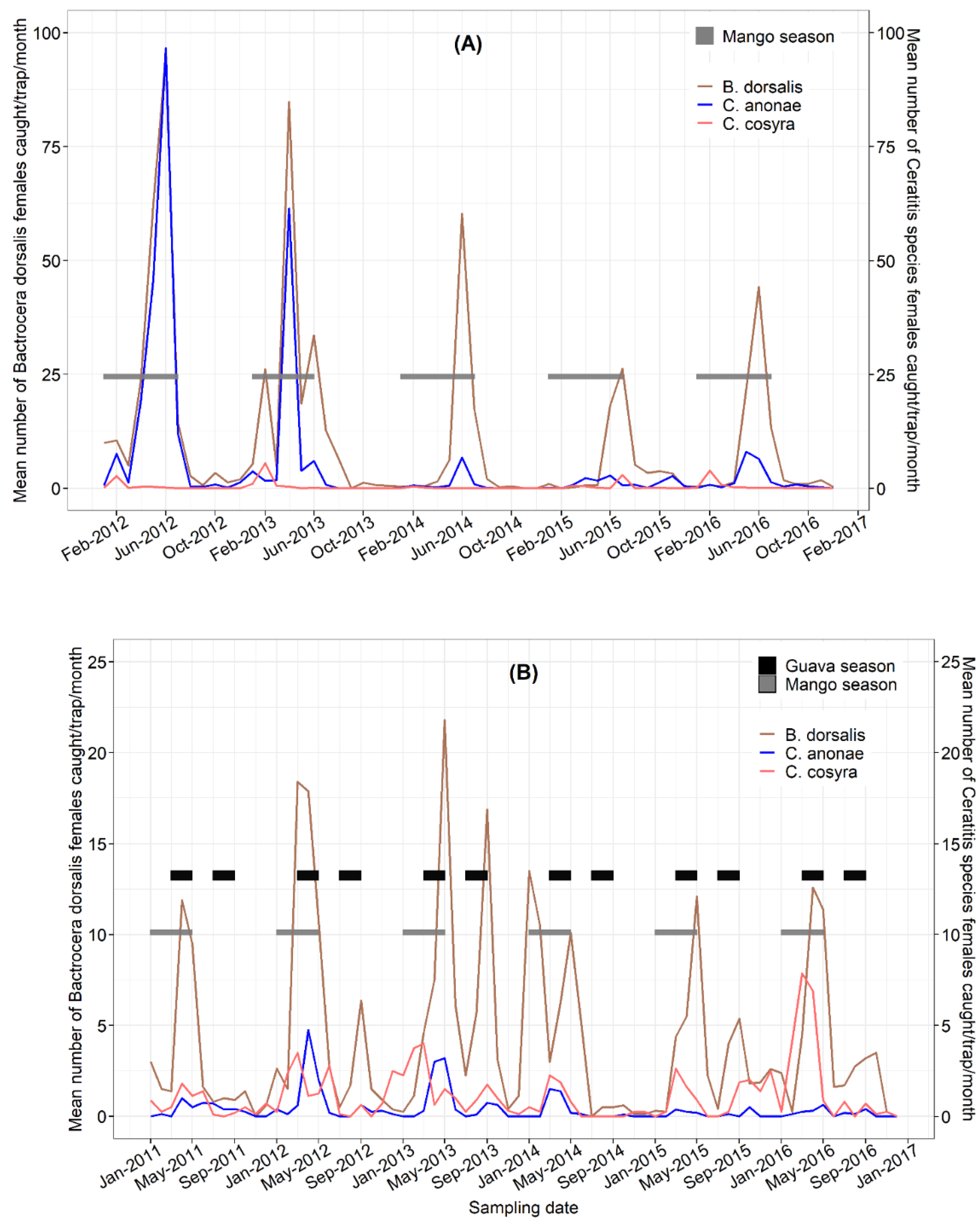
3.4. Seasonal Fluctuations
3.5. Correlation of Fruit Fly Catches and Weather Variables
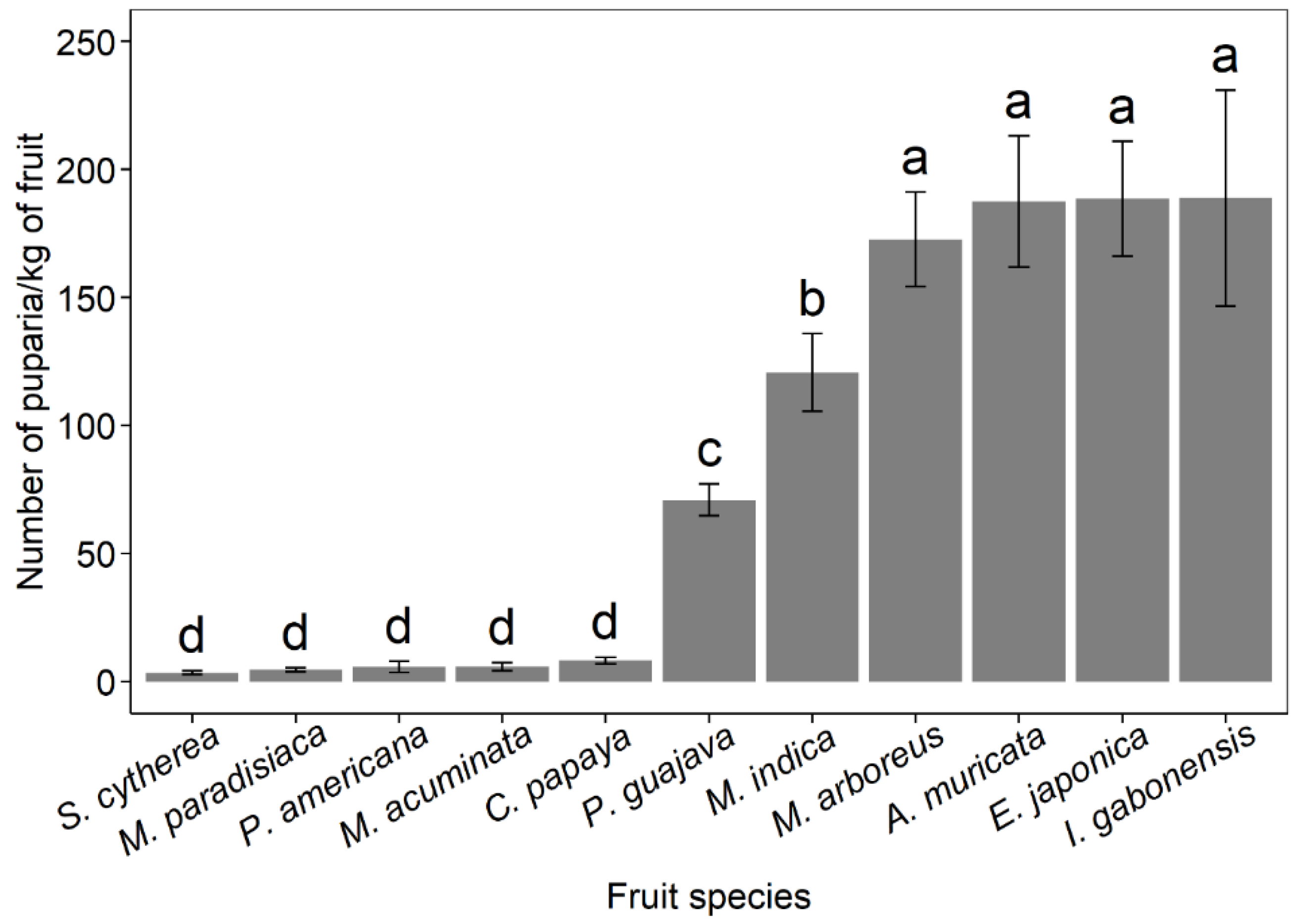
3.6. Host Range and Fruit Infestation Levels
4. Discussion
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- White, I.M.; Elson-Harris, M.M. Fruit Flies of Economic Significance: Their Identification and Bionomics; CAB International: Wallingford, UK, 1992. [Google Scholar]
- De Meyer, M.; Robertson, M.P.; Mansell, M.W.; Ekesi, S.; Tsuruta, K.; Mwaiko, W.; Vayssières, J.-F.; Peterson, A.T. Ecological niche and potential geographic distribution of the invasive fruit fly Bactrocera invadens (Diptera, Tephritidae). Bull. Entomol. Res. 2010, 100, 35–48. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- White, I.M. Taxonomy of the Dacinae (Diptera: Tephritidae) of Africa and the Middle East. Afr. Entomol. Mem. 2006, 2, 1–156. [Google Scholar]
- Gnanvossou, D.; Hanna, R.; Azandémè, G.; Georgen, G.; Tindo, T.; Agbaka, A. Inventaire et importance des dégâts des mouches de fruits sur quelques espèces de cucurbitaceae au Bénin. In Actes de l’atelier Scientifique National. Abomey Calavi; Adjanohoun, A., Igué, K., Eds.; IITA: Cotonou, Benin, 2008; Volume 6, pp. 140–145. [Google Scholar]
- De Meyer, M.; Ekesi, S. Exotic invasive fruit flies (Diptera: Tephritidae): In and out of Africa. In Fruit Fly Research and Development in Africa—Towards a Sustainable Management Strategy to Improve Horticulture; Ekesi, S., Mohamed, S.A., De Meyer, M., Eds.; Springer International Publishing: Cham, Switzerland, 2016; pp. 127–150. ISBN 978-3-319-43226-7. [Google Scholar]
- Lux, S.A.; Copeland, R.S.; White, I.M.; Manrakhan, A.; Billah, M.K. A new invasive fruit fly species from the Bactrocera dorsalis (Hendel) group detected in East Africa. Int. J. Trop. Insect Sci. 2003, 23, 355–361. [Google Scholar] [CrossRef]
- CABI. Invasive Species Compendium; CABI Publishing: Wallingford, UK, 2018. [Google Scholar]
- Hanna, R.; Gnanvossou, D.; Grout, T. Male annihilation for the control of Bactrocera invadens and Ceratitis cosyra in mango orchards. Fight. Fruit Veg. Flies Reg. West. Afr. 2008, 7, 3. [Google Scholar]
- Vayssières, J.-F.; Sinzogan, A.; Korie, S.; Ouagoussounon, I.; Thomas-Odjo, A. Effectiveness of spinosad bait sprays (GF-120) in controlling mango-infesting fruit flies (Diptera: Tephritidae) in Benin. J. Econ. Entomol. 2009, 102, 515–521. [Google Scholar] [CrossRef]
- Badii, K.B.; Billah, M.K.; Afreh-Nuamah, K.; Obeng-Ofori, D.; Nyarko, G. Review of the pest status, economic impact and management of fruit-infesting flies (Diptera: Tephritidae) in Africa. Afr. J. Agric. Res. 2015, 10, 1488–1498. [Google Scholar] [CrossRef] [Green Version]
- Rwomushana, I.; Ekesi, S.; Gordon, I.; Ogol, C.K. Host plants and host plant preference studies for Bactrocera invadens (Diptera: Tephritidae) in Kenya, a new invasive fruit fly species in Africa. Ann. Entomol. Soc. Am. 2008, 101, 331–340. [Google Scholar] [CrossRef] [Green Version]
- Mwatawala, M.W.; De Meyer, M.W.; Makundi, R.H.; Maerere, A.P. Host range and distribution of fruit-infesting pestiferous fruit flies (Diptera, Tephritidae) in selected areas of Central Tanzania. Bull. Entomol. Res. 2009, 99, 629. [Google Scholar] [CrossRef]
- Goergen, G.; Vayssières, J.-F.; Gnanvossou, D.; Tindo, M. Bactrocera invadens (Diptera: Tephritidae), a new invasive fruit fly pest for the Afrotropical region: Host plant range and distribution in West and Central Africa. Environ. Entomol. 2011, 40, 844–854. [Google Scholar] [CrossRef]
- Leblanc, L.; Fujita, B.; Stein, S.H.; Sawamura, W.K. Trapping records of fruit fly pest species (Diptera: Tephritidae) on Oahu (Hawaiian Islands): Analysis of spatial population trends. Proc. Hawaii. Entomol. Soc. 2012, 44, 89–97. [Google Scholar]
- EUNSPH Interceptions of harmful organisms in imported plants and other objects: Data from EUNSPH. Eur.-Eur. Union Notif. Syst. Plant Health EUNSPH 2018. Available online: https://food.ec.europa.eu/plants/plant-health-and-biosecurity/europhyt/interceptions_en (accessed on 5 May 2022).
- Ekesi, S.; Billah, M.K. A Field Guide to the Management of Economically Important Tephritid Fruit Flies in Africa; ICIPE Science Press: Nairobi, Kenya, 2006. [Google Scholar]
- Ekesi, S.; Vayssières, J.-F.; Hanna, R.; Meyer, M.D.; Goergen, G.; Malachrida, A. Status of Bactrocera invadens management in Africa: Recent advances, successes and challenges. In Abstracts of the 8th International Symposium on Fruit Flies of Economic Importance, Sabater; Munoz, B., Navarro, L.V., Garcia, U., Eds.; Editorial Universitat Politècnica: Valencia, Spain, 2010; p. 336. [Google Scholar]
- Appiah, E.F.; Ekesi, S.; Afreh-Nuamah, K.; Obeng-Ofori, D.; Mohamed, S.A. African weaver ant-produced semiochemicals impact on foraging behaviour and parasitism by the opiine parasitoid, Fopius arisanus on Bactrocera invadens (Diptera: Tephritidae). Biol. Control. 2014, 79, 49–57. [Google Scholar] [CrossRef]
- Muriithi, B.W.; Affognon, H.D.; Diiro, G.M.; Kingori, S.W.; Tanga, C.M.; Nderitu, P.W.; Mohamed, S.A.; Ekesi, S. Impact assessment of Integrated Pest Management (IPM) strategy for suppression of mango-infesting fruit flies in Kenya. Crop. Prot. 2016, 81, 20–29. [Google Scholar] [CrossRef] [Green Version]
- Vayssieres, J.-F.; Adandonon, A.; Sinzogan, A.; Korie, S. Diversity of fruit fly species (Diptera: Tephritidae) associated with citrus crops (Rutaceae) in southern Benin in 2008–2009. Int. J. Biol. Chem. Sci. 2010, 4, 6. [Google Scholar] [CrossRef]
- Souza-Filho, M.F.; Raga, A.; Azevedo-Filho, J.A.; Strikis, P.C.; Guimarães, J.A.; Zucchi, R.A. Diversity and seasonality of fruit flies (Diptera: Tephritidae and Lonchaeidae) and their parasitoids (Hymenoptera: Braconidae and Figitidae) in orchards of guava, loquat and peach. Braz. J. Biol. 2009, 69, 31–40. [Google Scholar] [CrossRef] [Green Version]
- Ukey, N.S.; Chandele, A.G.; Wagh, S.S.; Bansode, G.M. Species composition of fruit flies, Bactrocera spp. (Diptera: Tephritidae) infesting guava in Maharashrtra. Pest Manag. Hortic. Ecosyst. 2013, 19, 242–244. [Google Scholar]
- Amice, R.; Sales, F. Seasonal abundance of fruit flies in New Caledonia. In Management of Fruit Flies in the Pacific; Allwood, A.J., Drew, R.A.I., Eds.; Australian Centre for International Agricultural Research: Canberra, Australia, 1997; pp. 134–139. [Google Scholar]
- Manrakhan, A. Detection and monitoring of fruit flies in Africa. In Fruit Fly Research and Development in Africa—Towards a Sustainable Management Strategy to Improve Horticulture; Ekesi, S., Mohamed, S., De Meyer, M., Eds.; Springer International Publishing: Cham, Switzerland, 2016; pp. 253–273. [Google Scholar]
- Ekesi, S.; Mohamed, S.; Tanga, C.M. Comparison of food-based attractants for Bactrocera invadens (Diptera: Tephritidae) and evaluation of Mazoferm—Spinosad bait spray for field suppression in mango. J. Econ. Entomol. 2014, 107, 299–309. [Google Scholar] [CrossRef] [Green Version]
- Gnanvossou, D.; Hanna, R.; Goergen, G.; Salifu, D.; Tanga, C.M.; Mohamed, S.A.; Ekesi, S. Diversity and seasonal abundance of tephritid fruit flies in three agro-ecosystems in Benin, West Africa. J. Appl. Entomol. 2017, 141, 798–809. [Google Scholar] [CrossRef]
- Manrakhan, A.; Daneel, J.-H.; Beck, R.; Virgilio, M.; Meganck, K.; De Meyer, M. Efficacy of trapping systems for monitoring of Afrotropical fruit flies. J. Appl. Entomol. 2017, 141, 825–840. [Google Scholar] [CrossRef]
- Hanna, R.; Gnanvossou, D.; Goergen, G.; Bokonon-Ganta, A.H.; Mohamed, S.A.; Ekesi, S.; Fiaboe, K.K.M.; Agnontchémè, A.I. Efficiency of food-based attractants for monitoring tephritid fruit flies diversity and abundance in mango systems across three West African agro-ecological zones. J. Econ. Entomol. 2020, 113, 860–871. [Google Scholar] [CrossRef]
- Rwomushana, I.; Ekesi, S.; Ogol, C.K.P.O.; Gordon, I. Effect of temperature on development and survival of immature stages of Bactrocera invadens (Diptera: Tephritidae). J. Appl. Entomol. 2008, 132, 832–839. [Google Scholar] [CrossRef]
- IAEA. Trapping Guideline for Areawide Fruit Fly Programmes; Non-Serial Publications: Vienna, Austria, 2003; 46p. [Google Scholar]
- Manrakhan, A. Fruit fly monitoring-purpose, tools and methodology. In A Field Guide to the Management of Economically Important Tephritid Fruit Flies in Africa; Ekesi, S., Billah, M.K., Eds.; ICIPE Science Press: Nairobi, Kenya, 2006; p. 14. [Google Scholar]
- Leblanc, L.; Vargas, R.I.; Rubinoff, D. Captures of pest fruit Flies (Diptera: Tephritidae) and non-target insects in BioLure and Torula yeast traps in Hawaii. Environ. Entomol. 2010, 39, 1626–1630. [Google Scholar] [CrossRef] [PubMed]
- FAO. Global Agro-Ecological Zones (GAEZv3.0) Model Documentation; FAO: Rome, Italy, 2012. [Google Scholar]
- Tindo, M.; Hanna, R.; Goergen, G. Preliminary survey of Tephritidae fruit flies (Diptera: Tephritidae) of economic importance in Southern Cameroun: Species diversity and potential for integrated management. In The First International Symposium on Crops Integrated Pest Management in the CEMAC Zone; Pefoura, A.M., Tindo, M., Ngoko, Z., Dibog, L., Eds.; 2005; pp. 39–43. [Google Scholar]
- Ngamo Tinkeu, L.; Ladang, D.; Vayssieres, J.-F.; Lyannaz, J. Diversité des espèces de mouches des fruits (Diptera: Tephritidae) dans un verger mixte dans la localité de Malang (Ngaoundéré, Cameroun). Int. J. Biol. Chem. Sci. 2010, 4, 1425–1434. [Google Scholar] [CrossRef]
- Ndzana Abanda, F.-X.; Quilici, S.; Vayssières, J.-F.; Kouodiekong, L.; Woin, N. Inventaire des espèces de mouches des fruits sur goyave dans la région de Yaoundé au Cameroun. Fruits 2008, 63, 19–26. [Google Scholar] [CrossRef] [Green Version]
- Mokam, D.G.; Aln, D.C.; Lumaret, J.-P.; Delvare, G.; Djieto-Lordon, C. First record of Dacus (Lophodacus) hamatus (Diptera: Tephritidae) in Cameroon, with emphasis on a new host plant Lagenaria siceraria (Cucurbitaceae). Afr. J. Agric. Res. 2014, 9, 636–642. [Google Scholar] [CrossRef]
- Ngom, E.; Ndjogui, E.; Nkongho, R.N.; Iyabano, A.; Levang, P.; Miaro, L.; Feintrenie, L. Diagnostic du secteur élæicole au Cameroun; CIRAD: Yaoundé, Cameroun, 2014. [Google Scholar] [CrossRef]
- Heumou, C.R.; Djiéto-Lordon, C.; Aléné, C.D.; Elono Azang, S.P. Diversity and agronomic status of tomato and pepper fruit pests in two agro-ecological zones of Southern Cameroon: Western highland and the Southern Plateau of Cameroon. Afr. J. Agric. Res. 2015, 10, 1224–1232. [Google Scholar]
- Isabirye, B.E.; Akol, A.M.; Mayamba, A.; Nankinga, C.K.; Rwomushana, I. Species composition and community structure of fruit flies (Diptera: Tephritidae) across major mango-growing regions in Uganda. Int. J. Trop. Insect Sci. 2015, 35, 69–79. [Google Scholar] [CrossRef]
- De Meyer, M. Revision of the subgenus Ceratitis (Pardalaspis) Bezzi, 1918 (Diptera, Tephritidae, Ceratitini). Syst. Entomol. 1996, 21, 15–26. [Google Scholar] [CrossRef]
- Virgilio, M.; White, I.; De Meyer, M. A set of multi-entry identification keys to African frugivorous flies (Diptera, Tephritidae). ZooKeys 2014, 428, 97–108. [Google Scholar] [CrossRef]
- Magurran, A.E. Measuring Biological Diversity; Blackwell Pub: Malden, MA, USA, 2004; ISBN 978-0-632-05633-0. [Google Scholar]
- Sarada, G.; Maheswari, T.U.; Purushotham, K. Seasonal incidence and population fluctuation of fruit flies in mango and guava. Indian J. Entomol. 2001, 63, 272–276. [Google Scholar]
- Ekesi, S.; Nderitu, P.W.; Rwomushana, I. Field infestation, life history and demographic parameters of the fruit fly Bactrocera invadens (Diptera: Tephritidae) in Africa. Bull. Entomol. Res. 2006, 96, 379–386. [Google Scholar]
- Colwell, R.K. Biodiversity: Concepts, patterns, and measurement. Princet. Guide Ecol. Princet. Univ. Press Princet. 2009, 663, 257–263. [Google Scholar]
- R Development Core Team R. A Language and Environment for Statistical Computing; R Foundation for Statistical Computer: Vienna, Austria, 2019. [Google Scholar]
- Vayssières, J.-F.; Goergen, G.; Lokossou, O.; Dossa, P.; Akponon, C. A new Bactrocera species in Benin among mango fruit fly (Diptera: Tephritidae) species. Fruits 2005, 60, 371–377. [Google Scholar] [CrossRef] [Green Version]
- Mwatawala, M.W.; De Meyer, M.W.; Makundi, R.H.; Maerere, A.P. Seasonality and host utilization of the invasive fruit fly, Bactrocera invadens (Dipt., Tephritidae) in Central Tanzania. J. Appl. Entomol. 2006, 130, 530–537. [Google Scholar] [CrossRef]
- Epsky, N.D.; Kendra, P.E.; Schnell, E.Q. History and development of food-based attractants. In Trapping and the Detection, Control, and Regulation of Tephritid Fruit Flies; Shelly, T., Epsky, N.B.J.E., Reyes-Flores, J., Vargas, R., Eds.; Springer: Dordrecht, The Netherlands, 2014; p. 643. [Google Scholar]
- Vayssières, J.-F.; De Meyer, M.; Ouagoussounon, I.; Sinzogan, A.; Adandonon, A.; Korie, S.; Wargui, R.; Anato, F.; Houngbo, H.; Didier, C.; et al. Seasonal abundance of mango fruit flies (Diptera: Tephritidae) and ecological implications for their management in mango and cashew orchards in Benin (Centre & North). J. Econ. Entomol. 2015, 108, 2213–2230. [Google Scholar] [CrossRef]
- Rwomushana, I.; Ekesi, S.; Ogol, C.K.P.O.; Gordon, I. Mechanisms contributing to the competitive success of the invasive fruit fly Bactrocera invadens over the indigenous mango fruit fly, Ceratitis cosyra: The role of temperature and resource pre-emption. Entomol. Exp. Appl. 2009, 133, 27–37. [Google Scholar] [CrossRef]
- Jose, L.; Cugala, D.; Santos, L. Assessment of invasive fruit fly fruit infestation and damage in Cabo Delgado province, northern Mozambique. Afr. Crop. Sci. J. 2013, 21, 21–28. [Google Scholar]
- Ouédraogo, S.N.; Vayssières, J.-F.; Rémy Dabiré, A.; Rouland-Lefèvre, C. Biodiversité des mouches des fruits (Diptera: Tephritidae) en vergers de manguiers de l’Ouest du Burkina Faso: Structure et comparaison des communautés de différents sites. Fruits 2011, 66, 393–404. [Google Scholar] [CrossRef] [Green Version]
- N’Dépo, O.R.; Hala, N.; Allou, K.; Aboua, L.R.; Kouassi, K.P.; Vayssières, J.-F.; De Meyer, M. Abondance des mouches des fruits dans les zones de production fruitières de Côte d’Ivoire: Dynamique des populations de Bactrocera invadens (Diptera: Tephritidae). Fruits 2009, 64, 313–324. [Google Scholar] [CrossRef]
- Geurts, K.; Mwatawala, M.W.; De Meyer, M. Dominance of an invasive fruit fly species, Bactrocera invadens, along an altitudinal transect in Morogoro, Eastern Central Tanzania. Bull. Entomol. Res. 2014, 104, 288–294. [Google Scholar] [CrossRef]
- De Meyer, M.; Copeland, R.S.; Lux, S.; Mansell, M.; Whartone, R.; White, I.M.; Zenz, N. Annotated checklist of host plants for Afrotropical fruit flies (Diptera: Tephritidae) of the genus Ceratitis. Zoologische Documentatie KoninklijkMuseum voor Midden Afrika 2002, 27, 1–92. [Google Scholar]
- De Villiers, M.; Manrakhan, A.; Addison, P.; Hattingh, V. The Distribution, relative abundance, and seasonal phenology of Ceratitis capitata, Ceratitis rosa, and Ceratitis cosyra (Diptera: Tephritidae) in South Africa. Environ. Entomol. 2013, 42, 831–840. [Google Scholar] [CrossRef] [PubMed]
- Messing, R. Managing fruit flies on farms in Hawaii. Insect Pests, College of Tropical Agriculture and Human Resources, University of Hawaii at Manoa. Insect Pests 1999, 4, 1–8. [Google Scholar]
- Bateman, M.A. The ecology of fruit flies. Annu. Rev. Entomol. 1972, 17, 493–518. [Google Scholar] [CrossRef]
- Theron, C.D.; Manrakhan, A.; Weldon, C.W. Host use of the oriental fruit fly, Bactrocera dorsalis (Hendel) (Diptera: Tephritidae), in South Africa. J. Appl. Entomol. 2017, 141, 810–816. [Google Scholar] [CrossRef] [Green Version]
- Raghu, S.; Drew, R.A.I.; Clarke, A.R. Influence of host plant structure and microclimate on the abundance and behavior of a tephritid fly. J. Insect Behav. 2004, 17, 179–190. [Google Scholar] [CrossRef]
- CABI Bactrocera dorsalis. Invasive Species Compendium; CAB International: Wallingford, UK, 2022. [Google Scholar]
- Hanna, R.; Agnontcheme, D.; Gnanvossou, D.; Goergen, G.; Agbaka, A. Inventaire et importance des dégâts des mouches des fruits sur quelques variétés d agrumes au Bénin. In Actes de l’atelier Scientifique National, 6; Bibliotheque Nationale: Abomey-Calavi, Benin, 2008; pp. 128–139. [Google Scholar]
- Copeland, R.S.; Wharton, R.A.; Luke, Q.; De Meyer, M.; Lux, S.; Zenz, N.; Machera, P.; Okumu, M. Geographic distribution, host fruit, and parasitoids of African fruit fly pests Ceratitis anonae, Ceratitis cosyra, Ceratitis fasciventris, and Ceratitis rosa (Diptera: Tephritidae) in Kenya. Ann. Entomol. Soc. Am. 2006, 99, 261–278. [Google Scholar] [CrossRef]
- Cugala, D.; Ekesi, S.; Ambasse, D.; Adamu, R.S.; Mohamed, S.A. Assessment of ripening stages of Cavendish dwarf bananas as host or non-host to Bactrocera invadens. J. Appl. Entomol. 2014, 138, 449–457. [Google Scholar] [CrossRef]
- gbif.org Search. Available online: https://www.gbif.org/species/search?q= (accessed on 5 May 2022).
- Gnanvossou, D.; Hanna, R.; Bokonon-Ganta, A.; Ekesi, S.; Mohamed, S.A. Release, establishment and spread of the natural enemy Fopius arisanus (Hymenoptera: Braconidae) for control of the invasive Oriental fruit fly Bactrocera dorsalis (Diptera: Tephritidae) in Benin, West Africa. In Fruit Fly Research and Development in Africa—Towards a Sustainable Management Strategy to Improve Horticulture; Ekesi, S., Mohamed, S.A., De Meyer, M., Eds.; Springer: Cham, Switzerland, 2016; pp. 575–600. ISBN 978-3-319-43224-3. [Google Scholar]
- Nanga Nanga, S.; Hanna, R.; Gnanvossou, D.; Fotso Kuate, A.; Fiaboe, K.K.M.; Djieto-Lordon, C. Fruit preference, parasitism, and offspring fitness of Fopius arisanus (Hymenoptera: Braconidae) exposed to Bactrocera dorsalis’ (Diptera: Tephritidae) infested fruit species. Environ. Entomol. 2019, 48, 1286–1296. [Google Scholar] [CrossRef]
- Ekesi, S.; Billah, M.K.; Nderitu, P.W.; Lux, S.A.; Rwomushana, I. Evidence for competitive displacement of Ceratitis cosyra by the invasive fruit fly Bactrocera invadens (Diptera: Tephritidae) on mango and mechanisms contributing to the displacement. J. Econ. Entomol. 2009, 102, 981–991. [Google Scholar] [CrossRef]
- Badii, K.B.; Billah, M.K.; Afreh-Nuamah, K.; Obeng-Ofori, D. Species composition and host range of fruit-infesting flies (Diptera: Tephritidae) in Northern Ghana. Int. J. Trop. Insect Sci. 2015, 35, 137–151. [Google Scholar] [CrossRef]
- De Meyer, M.; Mohamed, S.; White, M. Invasive Fruit Fly Pests in Africa: A Diagnostic Tool and Information Reference for the four Asian Species of Fruit Fly (Diptera, Tephritidae) that Have Become Accidentally Established as Pests in Africa, Including the Indian Ocean Islands. Available online: http://www.africamuseum.be/fruitfly/AfroAsia.htm (accessed on 14 February 2016).






{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Location | Description of the Area | Fruit Species | Management |
|---|---|---|---|
| Foumbot WH-MR IRAD research station |
|
|
|
| Nkolbisson HF-BMR IITA research station |
|
|
|
| Location | Trapping Period | Male Lure | Food Bait |
|---|---|---|---|
| Foumbot/WH_MR | January 2012 to December 2016. |
| Torula yeast. |
| Nkolbisson/HF-BMR | January 2011 to December 2016. |
| Torula yeast. |
| January 2011 to December 2013. | BioLure. | ||
| January 2012 to December 2014. | Mazoferm. |
| Fruit Fly Species | WH-MR | HF-BR | Both AEZs | |||
|---|---|---|---|---|---|---|
| Total | Relative Abundance (%) | Total | Relative Abundance (%) | Total | Relative Abundance (%) | |
| Bactrocera dorsalis | 242,324 | 94.2 | 313,227 | 97.2 | 555,551 | 95.9 |
| Ceratitis cosyra | 1543 | 0.60 | 7176 | 2.23 | 8719 | 1.50 |
| Ceratitis anonae | 10,546 | 4.10 | 1641 | 0.51 | 12,187 | 2.10 |
| Ceratitis capitata | 96 | 0.04 | 49 | 0.02 | 145 | 0.03 |
| Ceratitis ditissima | 15 | 0.006 | 0 | - | 15 | 0.003 |
| Ceratitis punctata | 2 | 0.001 | 8 | 0.002 | 10 | 0.002 |
| Ceratitis bremii | 96 | 0.037 | 6 | 0.002 | 102 | 0.02 |
| Perilampsis sp. | 6 | 0.002 | 5 | 0.002 | 11 | 0.002 |
| Dacus bivittatus | 2555 | 0.99 | 0 | 2555 | 0.44 | |
| Dacus punctatifrons | 0 | 0 | 68 | 0.02 | 68 | 0.01 |
| Total captured | 257,183 | 100 | 322,180 | 100 | 579,363 | 100 |
| Trapping Year | Male Lures | Food Bait | ||||||
|---|---|---|---|---|---|---|---|---|
| Methyl Eugenol | Terpinyl Acetate | Torula Yeast | ||||||
| B. dorsalis | C. cosyra | C. anonae | All | B. dorsalis | C. cosyra | C. anonae | All | |
| Foumbot WH-MR | ||||||||
| 2012 | 412.2 ± 50.7 a | 5.41 ± 0.79 a | 18.9 ± 3.23 a | 24.6 ± 3.81 a | 19.8 ± 4.66 a (54.8) | 0.28 ± 0.11 ab (54.5) | 16.2 ± 4.91 a (41.9) | 36.3 ± 9.34 a (49.1) |
| 2013 | 464.9 ± 77.7 a | 1.80 ± 0.39 b | 18.9 ± 4.88 a | 20.7 ± 5.16 a | 15.8 ± 2.70 ab (57.9) | 0.60 ± 0.22 a (52.2) | 6.28 ± 2.45 b (36.8) | 22.63 ± 5.73 a (52.0) |
| 2014 | 88.1 ± 9.94 c | 1.07 ± 0.23 bc | 2.06 ± 0.42 b | 3.12 ± 0.49 b | 7.04 ± 2.70 c (47.0) | 0.04 ± 0.03 b (55.6) | 0.76 ± 0.66 c (52.2) | 7.84 ± 2.92 b (47.6) |
| 2015 | 161.6 ± 24.3 bc | 1.11 ± 0.27 bc | 5.22 ± 1.24 b | 6.97 ± 1.39 b | 5.29 ± 1.26 c (65.0) | 0.33 ± 0.16 ab (40.4) | 1.16 ± 0.22 c (36.6) | 6.78 ± 1.39 b (58.7) |
| 2016 | 303.3 ± 51.4 ab | 0.17 ± 0.06 c | 0.92 ± 0.23 b | 1.09 ± 0.25 b | 7.65 ± 1.96 bc (63.7) | 0.51 ± 0.22 a (23.6) | 1.80 ± 0.46 c (59.8) | 9.97 ± 2.33 b (61.0) |
| Nkolbisson HF-BR | ||||||||
| 2011 | 452.3 ± 57.9 a | 1.59 ± 0.23 d | 0.04 ± 0.02 b | 1.63 ± 0.23 c | 5.99 ± 0.92 a (52.3) | 0.95 ± 0.16 c (33.3) | 0.63 ± 0.16 ab (43.1) | 7.57 ± 1.05 a (49.2) |
| 2012 | 420.7 ± 54.2 a | 3.72 ± 0.50 bc | 0.07 ± 0.03 b | 3.79 ± 0.50 c | 9.35 ± 2.15 a (41.5) | 2.26 ± 0.48 ab (42.1) | 0.97 ± 0.28 a (23.8) | 12.7 ± 2.62 a (40.2) |
| 2013 | 446.0 ± 65.8 a | 2.44 ± 0.36 cd | 0.06 ± 0.02 b | 2.50 ± 0.36 c | 10.0 ± 1.43 a (39.3) | 1.98 ± 0.32 ab (25.7) | 0.89 ± 0.18 ab (20.4) | 12.9 ± 1.62 a (39.9) |
| 2014 | 598.1 ± 86.6 a | 5.58 ± 0.92 b | 1.96 ± 0.49 a | 7.57 ± 1.31 b | 7.07 ± 1.43 a (36.6) | 0.76 ± 0.20 c (34.2) | 0.42 ± 0.16 bcd (38.6) | 8.30 ± 1.54 a (38.2) |
| 2015 | 425.4 ± 60.0 a | 17.75 ± 2.53 a | 1.92 ± 0.61 a | 19.8 ± 2.74 a | 5.88 ± 0.92 a (42.1) | 1.39 ± 0.27 bc (19.7) | 0.19 ± 0.06 d (35.0) | 7.62 ± 1.04 a (37.7) |
| 2016 | 588.4 ± 109.8 a | 21.27 ± 2.36 a | 0.64 ± 0.16 b | 22.0 ± 2.43 a | 6.39 ± 1.24 a (41.7) | 3.13 ± 0.76 a (41.1) | 0.24 ± 0.08 cd (28.0) | 9.85 ± 1.67 a (41.1) |
| Attractant | Bactrocera dorsalis | Ceratitis cosyra | Ceratitis anonae | All Fruit Flies |
|---|---|---|---|---|
| BioLure | 5.33 ± 0.52 | 1.25 ± 0.16 | 1.94 ± 0.34 | 8.66 ± 0.81 |
| Torula yeast | 8.45 ± 0.92 | 1.73 ± 0.20 | 0.83 ± 0.12 | 11.1 ± 1.09 |
| t-ratio, df, p-value | 3.36, 155, <0.001 | 2.18, 155, 0.030 | −3.48, 155, <0.001 | 2.46, 155, 0.016 |
| Mazoferm-6% | 3.96 ± 0.56 | 0.69 ± 0.10 | 0.63 ± 0.11 | 5.30 ± 0.68 |
| Torula yeast | 8.81 ± 0.98 | 1.67 ± 0.21 | 0.76 ± 0.12 | 11.3 ± 1.15 |
| t-ratio, df, p-value | 4.40, 155, <0.001 | 4.60, 155, <0.001 | 1.30, 155, 0.194 | 4.75, 155, <0.001 |
| Weather Variables | Bactrocera dorsalis | Ceratitis cosyra | Ceratitis anonae | |||
|---|---|---|---|---|---|---|
| Male | Female | Male | Female | Male | Female | |
| Foumbot WH-MR | ||||||
| TempMin | 0.422 (0.001) | 0.450 (0.001) | 0.186 (0.154) | 0.025 (0.848) | 0.336 (0.009) | 0.372 (0.003) |
| TempMean | −0.331 (0.010) | 0.033 (0.804) | −0.062 (0.636) | 0.442 (0.001) | −0.256 (0.049) | 0.117 (0.375) |
| TempMax | −0.496 (0.001) | −0.270 (0.037) | −0.300 (0.020) | 0.294 (0.023) | −0.510 (0.001) | −0.203 (0.119) |
| RHMin | 0.122 (0.352) | 0.119 (0.365) | 0.070 (0.596) | −0.025 (0.852) | 0.050 (0.703) | 0.007 (0.955) |
| RHMean | 0.129 (0.324) | 0.069 (0.600) | 0.028 (0.830) | −0.118 (0.371) | 0.032 (0.810) | −0.059 (0.652) |
| RHMax | 0.088 (0.505) | 0.019 (0.887) | 0.076 (0.565) | −0.120 (0.362) | 0.134 (0.309) | 0.001 (0.998) |
| Rainfall | 0.298 (0.021) | 0.132 (0.313) | −0.187 (0.154) | −0.199 (0.127) | −0.042 (0.750) | −0.042 (0.750) |
| Nkolbisson HF-BR | ||||||
| TempMin | 0.145 (0.223) | 0.079 (0.507) | −0.179 (0.131) | −0.079 (0.509) | −0.079 (0.509) | −0.079 (0.509) |
| TempMean | −0.003 (0.980) | 0.190 (0.109) | 0.171 (0.152) | −0.037 (0.756) | −0.037 (0.756) | −0.037 (0.756) |
| TempMax | −0.095 (0.429) | 0.063 (0.598) | 0.397 (0.001) | 0.146 (0.221) | 0.146 (0.221) | 0.146 (0.221) |
| RHMin | 0.268 (0.023) | 0.054 (0.651) | −0.247 (0.036) | 0.111 (0.354) | 0.111 (0.354) | 0.111 (0.354) |
| RHMean | 0.267 (0.024) | 0.062 (0.607) | −0.156 (0.191) | 0.178 (0.134) | 0.178 (0.134) | 0.178 (0.134) |
| RHMax | 0.145 (0.225) | 0.077 (0.523) | −0.085 (0.478) | 0.137 (0.252) | 0.137 (0.252) | 0.137 (0.252) |
| Rainfall | 0.535 (0.001) | 0.374 (0.001) | 0.209 (0.078) | 0.429 (0.001) | 0.429 (0.001) | 0.429 (0.001) |
| Plant Family/Species | Origin | Sampling Site | Weight (kg) | No. of Emerged Adult Fruit Flies * | |||||
|---|---|---|---|---|---|---|---|---|---|
| B. dorsalis | C. cosyra | C. anonae | C. capitata | C. ditissima | D. bivittatus | ||||
| Western Highlands with Monomodal Rainfall | |||||||||
| Anacardiaceae | |||||||||
| Mangifera indica L. | Exotic | Baba I, Bamessingue, Bansoa, Foumbot | 964 | 14,462 (98.1) | 7 (0.05) | 269 (1.83) | - | - | - |
| Lauraceae | - | - | - | - | - | ||||
| Persea americana Miller | Exotic | Foumbot | 71.4 | 14 (100) | - | - | - | - | - |
| Myrtaceae | |||||||||
| Psidium guajava L. | Exotic | Baba I, Bamessingue, Bansoa, Foumbot | 1186 | 22,058 (91.0) | 30 (0.12) | 2124 (8.76) | 9 (0.04) | 17 (0.07) | - |
| Rosaceae | |||||||||
| Eriobotrya japonica (Thunb.) Lindley | Exotic | Santa | 3.0 | 415 (99.5) | - | - | - | 2 (0.48) | - |
| Rubiaceae | |||||||||
| Sarcocephalus latifolius (Smith) Bruce | Native | Tonga | 6.03 | - | 314 (100) | - | - | - | - |
| Rutaceae | |||||||||
| Citrus limon (L.) Baufman f. | Exotic | Foumbot | 3.10 | - | - | - | - | - | - |
| Citrus reticulata Blanco | Exotic | Foumbot | 3.20 | 17 (100) | - | - | - | - | - |
| Citrus sinensis (L.) Osbeck | Exotic | Foumbot | 5.35 | - | - | - | - | - | - |
| Plant Family/Species | Origin | Sampling Site | Weight (kg) | No. of Emerged Adult Fruit Flies * | |||||
| B. dorsalis | C. cosyra | C. anonae | C. capitata | C. ditissima | D. bivittatus | ||||
| Humid Forest with Bimodal Rainfall | |||||||||
| Anacardiaceae | |||||||||
| Mangifera indica L. | Exotic | Nkolbisson | 800 | 43,075 (99.6) | 26 (0.06) | 140 (0.32) | 4 (0.01) | - | - |
| Spondias cytherea Sonner | Exotic | Nkolbisson | 108 | 126 (99.2) | 1 (0.79) | - | - | - | - |
| Trichoscypha abutEngl. | Native | Atin-Odzoé | 0.39 | 1 (14.3) | - | 6 (85.7) | - | - | - |
| Annonaceae | |||||||||
| Annona montana L. | Exotic | Tsinga | 6.44 | 3 (11.5) | 23 (88.5) | - | - | - | - |
| Annona muricata L. | Exotic | Nkolbisson | 96 | 1159 (9.22) | 11,265 (89.7) | 142 (1.13) | - | - | - |
| Annona squamosa L. | Exotic | Nkolbisson | 164 | 2406 (55.6) | 961 (22.2) | 960 (22.2) | - | - | - |
| Apocynaceae | |||||||||
| Voacanga africanaStapf. | Native | Tsinga, Nkolbisson | 1.1 | 1 (14.3) | - | - | - | 6 (85.7) | - |
| Burseraceae | |||||||||
| Dacryodes edulis(G. Don) Lam | Native | Nkolbisson | 27 | 17 (100) | - | - | - | - | - |
| Caricaceae | |||||||||
| Carica papaya L. | Exotic | Nkolbisson | 628 | 2436 (77.8) | - | 8 (0.26) | - | - | 687 (21.9) |
| Cercopriaceae | |||||||||
| Myrianthus arboreus-P.-Beauv | Native | Nkolbisson | 142 | 99 (1) | 1 (0.01) | 9803 (98.8) | - | 15 (0.15) | - |
| Clusiaceae | |||||||||
| Allanblackia floribunda Oliv. | Exotic | Mbalmayo | 5.3 | ||||||
| Garcinia xanthochymus Hook. f. | Exotic | Mbalmayo | 0.06 | - | - | - | 10 (100) | - | - |
| Irvingiaceae | |||||||||
| Irvingia gabonensis (Aubry Lecomte) Baill | Native | Mbalmayo, Nkolbisson | 138 | 20,775 (100) | - | - | - | - | - |
| Irvingia womboluVermoesen | Native | Abondo | 79 | 6391 (100) | - | - | - | - | - |
| Plant Family/Species | Origin | Sampling Site | Weight (kg) | No. of Emerged Adult Fruit Flies * | |||||
| B. dorsalis | C. cosyra | C. anonae | C. capitata | C. ditissima | D. bivittatus | ||||
| Humid Forest with Bimodal Rainfall | |||||||||
| Lauraceae | |||||||||
| Persea americana Miller | Exotic | Nkolbisson | 287 | 227 (38) | 353 (59.0) | 18 (3.01) | - | - | - |
| Moraceae | |||||||||
| Ficus mucuso Filcalho | Native | Nkolbisson | 20.5 | - | - | - | - | - | - |
| Musaceae | |||||||||
| Musa acuminate L. | Exotic | Nkolbisson | 479.5 | 1421 (99.7) | - | 4 (0.28) | - | - | - |
| Musa paradisiaca L. | Exotic | Nkolbisson | 564.3 | 1778 (99.2) | - | 15 (0.84) | - | - | - |
| Myrtaceae | |||||||||
| Psidium guajava L. | Exotic | Nkolbisson | 849.6 | 34,619 (87.2) | 2392 (6.03) | 2569 (6.47) | 98 (0.25) | 7 (0.02) | - |
| Oxalidaceae | |||||||||
| Averrhoa carambola L. | Exotic | Tsinga | 12.7 | 3 (100) | - | - | - | - | - |
| Rosaceae | |||||||||
| Eriobotrya japonica (Thunb.) Lindley | Exotic | Nkolbisson | 32.4 | 1413 (56.3) | 5 (0.2) | 1070 (42.6) | 24 (0.96) | - | - |
| Rubiaceae | |||||||||
| Sarcocephalus latifolius (Smith) Bruce | Native | Makénéné | 9.69 | 281 (29.2) | 681 (70.8) | - | - | - | - |
| Rutaceae | |||||||||
| Citrus limon (L.) Baufman f. | Exotic | Nkolbisson | 20.9 | 7 (100) | - | - | - | - | - |
| Citrus reticulata Blanco | Exotic | Nkolbisson | 1.43 | - | - | - | - | - | - |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Nanga Nanga, S.; Hanna, R.; Fotso Kuate, A.; Fiaboe, K.K.M.; Nchoutnji, I.; Ndjab, M.; Gnanvossou, D.; Mohamed, S.A.; Ekesi, S.; Djieto-Lordon, C. Tephritid Fruit Fly Species Composition, Seasonality, and Fruit Infestations in Two Central African Agro-Ecological Zones. Insects 2022, 13, 1045. https://doi.org/10.3390/insects13111045
Nanga Nanga S, Hanna R, Fotso Kuate A, Fiaboe KKM, Nchoutnji I, Ndjab M, Gnanvossou D, Mohamed SA, Ekesi S, Djieto-Lordon C. Tephritid Fruit Fly Species Composition, Seasonality, and Fruit Infestations in Two Central African Agro-Ecological Zones. Insects. 2022; 13(11):1045. https://doi.org/10.3390/insects13111045
Chicago/Turabian StyleNanga Nanga, Samuel, Rachid Hanna, Apollin Fotso Kuate, Komi K. M. Fiaboe, Ibrahim Nchoutnji, Michel Ndjab, Désiré Gnanvossou, Samira A. Mohamed, Sunday Ekesi, and Champlain Djieto-Lordon. 2022. "Tephritid Fruit Fly Species Composition, Seasonality, and Fruit Infestations in Two Central African Agro-Ecological Zones" Insects 13, no. 11: 1045. https://doi.org/10.3390/insects13111045