
A Peculiar New Genus of Bibionomorpha (Diptera) with Brachycera-Like Modification of Antennae from Mid-Cretaceous Amber of Myanmar


{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:Simple Summary
Abstract
1. Introduction
2. Materials and Methods
3. Results
3.1. Systematic Paleontology
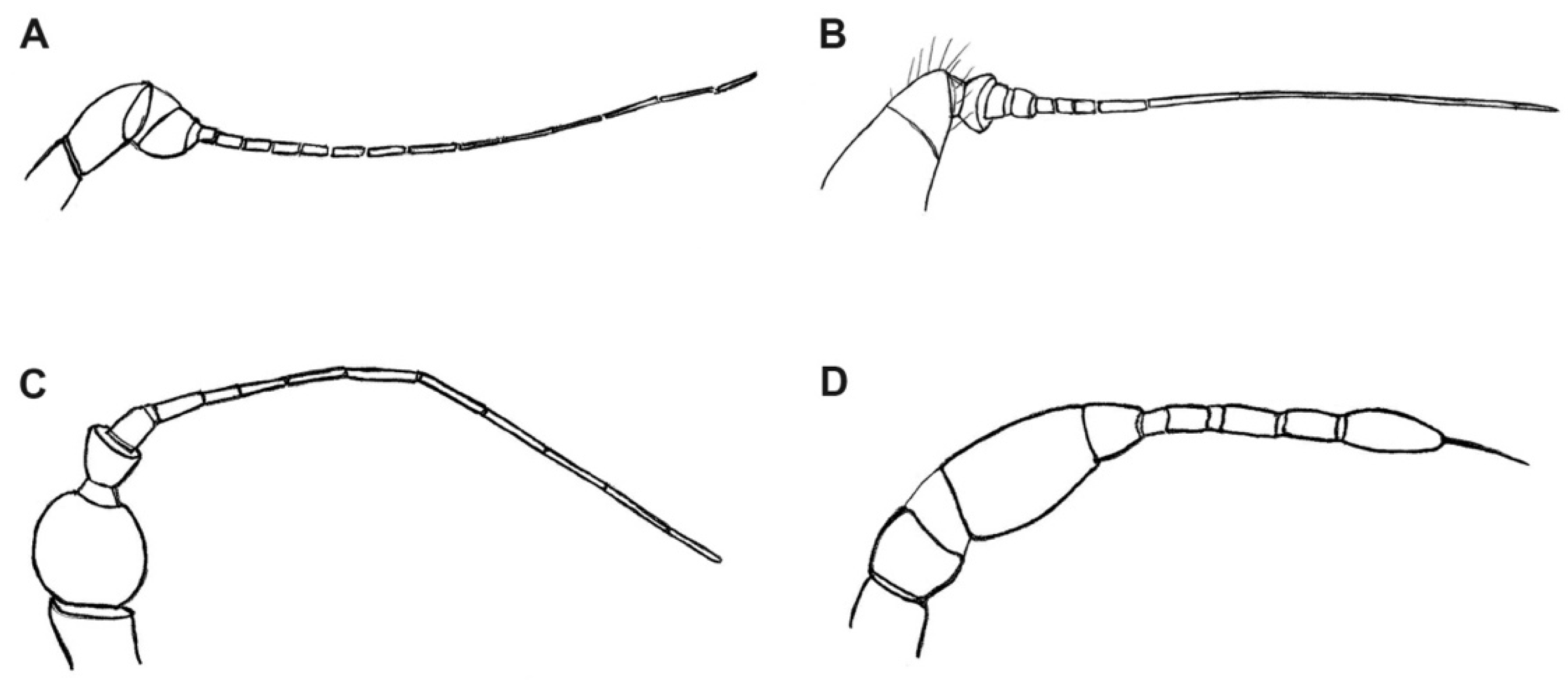
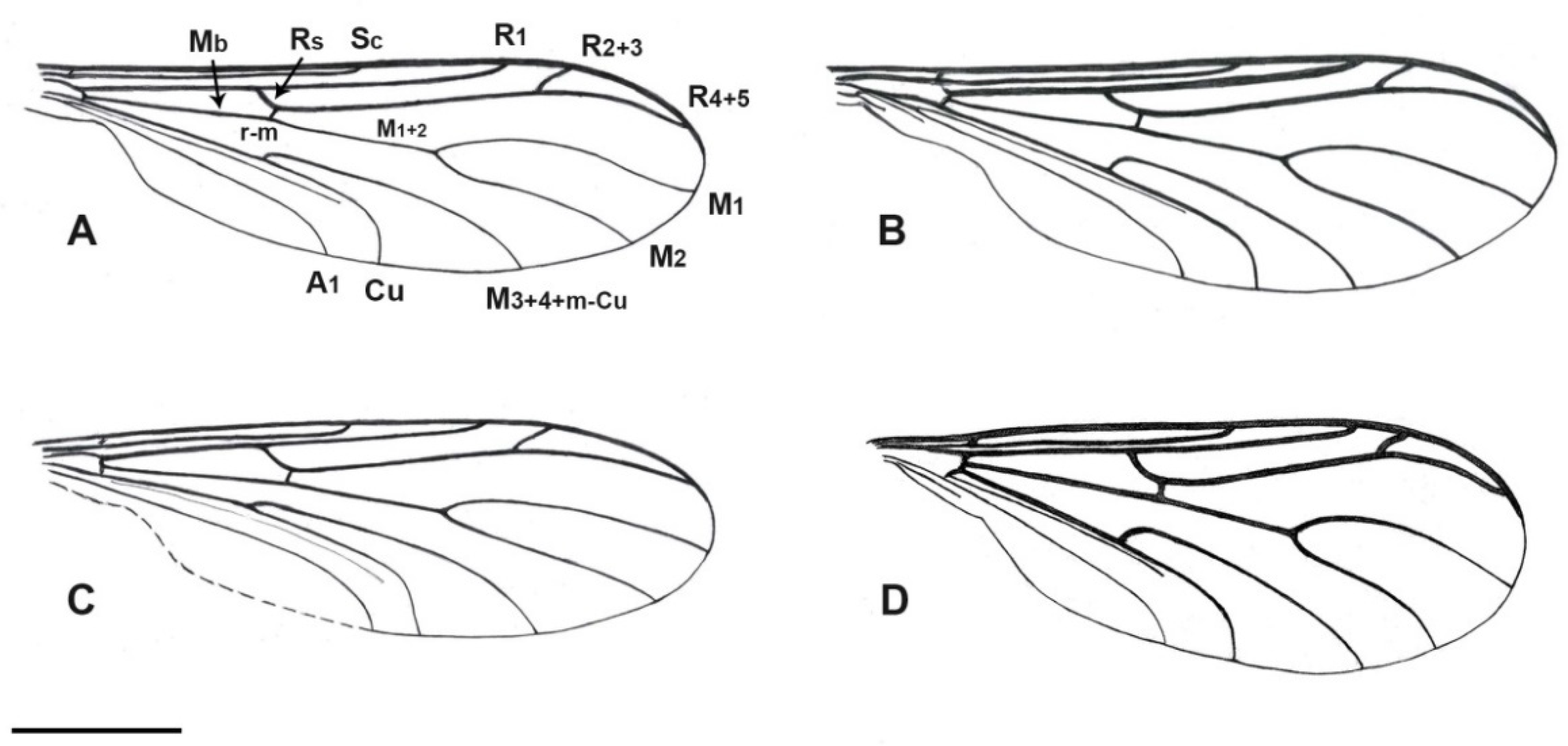
3.2. Description of a New Fossil Material
3.3. Identification Key to Species of Burmahesperinus gen. nov.
4. Discussion
5. Conclusions
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Evenhuis, N. Catalog of the Fossil Flies of the World (Insecta: Diptera), Version 2.0. Available online: http://hbs.bishopmuseum.org/fossilcat/ (accessed on 10 October 2020).
- Van Nieukerken, E.J.; Kaila, I.J.; Kristensen, N.P.; Kristensen, N.P.; Lees, D.C.; Minet, J.; Mitter, C.; Mutanen, M.; Regier, J.C.; Simonsen, T.J.; et al. Animal biodiversity: An outline of higher-level classification and survey of taxonomic richness. Zootaxa 2011, 3148, 222–229. [Google Scholar]
- Blagoderov, V.; Grimaldi, D.A.; Fraser, N.C. How Time Flies for Flies: Diverse Diptera from the Triassic of Virginia and Early Radiation of the Order. Am. Mus. Novit. 2007, 3572, 1–39. [Google Scholar] [CrossRef]
- Shcherbakov, D.E.; Lukashevich, E.D.; Blagoderov, V.A. Triassic Diptera and initial radiation of the order. Int. J. Dipt. Res. 1995, 6, 75–115. [Google Scholar]
- Krzeminski, W.; Krzeminska, E. Triassic Diptera: Descriptions, revisions and phylogenetic relations. Acta Zool. Cracov. 2003, 46, 153–184. [Google Scholar]
- Ševčík, J.; Kaspřák, D.; Mantič, M.; Fitzgerald, S.; Ševčíková, T.; Tóthová, A.; Jaschhof, M. Molecular phylogeny of the megadiverse insect infraorder Bibionomorpha sensu lato (Diptera). PeerJ 2016, 4, e2563. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Mantič, M.; Sikora, T.; Burdíková, N.; Blagoderov, V.; Kjærandsen, J.; Kurina, O.; Ševčík, J. Hidden in Plain Sight: Comprehensive Molecular Phylogeny of Keroplatidae and Lygistorrhinidae (Diptera) Reveals Parallel Evolution and Leads to a Revised Family Classification. Insects 2020, 11, 348. [Google Scholar] [CrossRef]
- Blagoderov, V.; Grimaldi, D. Fossil Sciaroidea (Diptera) in Cretaceous Ambers, Exclusive of Cecidomyiidae, Sciaridae, and Keroplatidae. Am. Mus. Novit. 2004, 3433, 1–76. [Google Scholar] [CrossRef]
- Ševčík, J.; Krzemiński, W.; Skibińska, K. Intriguing and Beautiful: Adamacrocera adami gen. et sp. nov. from the Upper Cretaceous Amber of Myanmar Represents a New Subfamily of Keroplatidae (Diptera: Bibionomorpha). Insects 2020, 11, 552. [Google Scholar] [CrossRef]
- Kalugina, N.S.; Kovalev, V.G. Two-Winged Flies of the Jurassic of Siberia; Nauka Press: Moscow, Russia, 1990; pp. 1–199. [Google Scholar]
- Kovalev, V.G. Two-winged flies. Muscida. In Late Mesozoic Insects of Eastern Transbaikalia; Nauka Press: Moscow, Russia, 1990; pp. 123–177. [Google Scholar]
- Skartveit, J.; Ansorge, J. A new genus and subfamily of fossil Bibionidae (Diptera) from the Lower Cretaceous, with new classification of the Bibionidae. Palaeoentomology 2020, 3, 163–172. [Google Scholar] [CrossRef]
- Poinar, G. Cascoplecia insolitis (Diptera: Cascopleciidae), a new family, genus, and species of flower-visiting, unicorn fly (Bibionomorpha) in Early Cretaceous Burmese amber. Cretac. Res. 2010, 31, 71–76. [Google Scholar] [CrossRef]
- Hardy, D.E. Bibionidae. In Manual of Nearctic Diptera; McAlpine, J.F., Peterson, B.V., Shewell, G.E., Teskey, H.J., Vockeroth, J.R., Wood, D.M., Eds.; Biosystematics Research Institute: Ottawa, ON, Canada, 1981; Volume 1, pp. 217–222, 674. [Google Scholar]
- Fitzgerald, S.J. Bibionidae (March Flies, Love Bugs). In Manual of Central American Diptera; Brown, B.V., Borkent, A., Cumming, J.M., Wood, D.M., Woodley, N.E., Zumbado, M.A., Eds.; NRC Research Press: Ottawa, ON, Canada, 2009; Volume 1, pp. 245–251, 714. [Google Scholar]
- Skartveit, J. Family Bibionidae. In Contributions to a Manual of Palaearctic Diptera; Papp, L., Darvas, B., Eds.; Science Herald: Budapest, Hungary, 1997; Volume 2, pp. 41–50, 592. [Google Scholar]
- Krivosheina, N.P. Pleciidae. In Catalogue of Palaearctic Diptera. Sciaridae—Anisopodidae; Soós, Á., Papp, L., Eds.; Akadémiai Kiadó: Budapest, Hungary, 1981; Volume 4, pp. 313–316, 441. [Google Scholar]
- Fitzgerald, S.J. Evolution and Classification of Bibionidae (Diptera: Bibionomorpha). Ph.D. Thesis, State University, Corvallis, OR, USA, 2004. [Google Scholar]
- Pinto, L.G.; Amorim, D.S. Bibionidae (Diptera: Bibionomorpha). In Morfologia e Análise Filogenética; Holos: Ribeirão Preto, Brazil, 2000. [Google Scholar]
- Grimaldi, D.; Amorim, D.S.; Blagoderov, V. The Mesozoic Family Archizelmiridae (Diptera: Insecta). J. Paleontol. 2003, 77, 368–381. [Google Scholar] [CrossRef]
- Krzemińska, E.; Krzemiński, W.; Dahl, C. Monograph of Fossil Trichoceridae (Diptera) over 180 Million Years of Evolution; Institute of Systematic and Evolution of Animals Polish Academy of Sciences: Krakow, Poland, 2009; pp. 1–171. [Google Scholar]
- Cruickshank, R.D.; Ko, K. Geology of an amber locality in the Hukawng Valley, northern Myanmar. J. Asian Earth Sci. 2003, 21, 441–445. [Google Scholar] [CrossRef]
- Grimaldi, D.A.; Engel, M.S.; Nascimbene, P.C. Fossiliferous Cretaceous amber from Myanmar (Burma): Its rediscovery, biotic diversity, and paleontological significance. Am. Mus. Novit. 2002, 3361, 1–71. [Google Scholar] [CrossRef] [Green Version]
- Shi, G.; Grimaldi, D.A.; Ge, H.; Wang, J.; Yang, M.; Lei, W.; Li, Q.; Li, X. Age constraint on Myanmar amber based on U-Pb dating of zircons. Cretac. Res. 2012, 37, 155–163. [Google Scholar] [CrossRef]
- Skartveit, J. Fossil Hesperinidae and Bibionidae (Diptera: Bibionoidea) from Baltic amber. Stud. Dipterol. 2008, 15, 3–42. [Google Scholar]
- Skartveit, J. Bibionidae. In Manual of Afrotropical Diptera. Nematocerous Diptera and Lower Brachycera; Kirk-Spriggs, A.H., Sinclair, B.J., Eds.; Suricata 5; SANBI Graphics & Editing: Pretoria, South Africa, 2017; Volume 2, pp. 425, 497–504. [Google Scholar]
- Arillo, A.; Blagoderov, V.; Peñalver, E. Early Cretaceous parasitism in amber: A new species of Burmazelmira fly (Diptera: Archizelmiridae) parasitized by a Leptus sp. mite (Acari, Erythraeidae). Cretac. Res. 2018, 86, 24–32. [Google Scholar] [CrossRef]
- Stuckenberg, B.R. Antennal evolution in the Brachycera (Diptera), with a reassessment of terminology relating to the flagellum. Stud. Dipterol. 1999, 6, 33–48. [Google Scholar]
- Amorim, D.S.; Yeates, D. Pesky gnats: Ridding dipteran classification of the “Nematocera”. Stud. Dipterol. 2006, 13, 3–9. [Google Scholar]
- Lambkin, C.L.; Sinclair, B.J.; Pape, T.; Courtney, G.W.; Skevington, J.H.; Meier, R.; Yeates, D.K.; Blagoderov, V.; Wiegmann, B.M. The phylogenetic relationships among infraorders and superfamilies of Diptera based on morphological evidence. Syst. Entomol. 2013, 38, 164–179. [Google Scholar] [CrossRef]
- Michelsen, V. Neodiptera: New insights into the adult morphology and higher level phylogeny of Diptera (Insecta). Zool. J. Linn. Soc. 1996, 117, 71–102. [Google Scholar] [CrossRef]
- Sinclair, B.J.; Cumming, J.M.; Brooks, S.E. Male terminalia of Diptera (Insecta): A review of evolutionary trends, homology and phylogenetic implications. Insect Syst. Evol. 2013, 44, 373–415. [Google Scholar] [CrossRef]
- Huber, B.A.; Sinclair, B.J.; Schmitt, M. The evolution of asymmetric genitalia in spiders and insects. Biol. Rev. 2007, 82, 647–698. [Google Scholar] [CrossRef] [PubMed]




Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Ševčík, J.; Skartveit, J.; Krzemiński, W.; Skibińska, K. A Peculiar New Genus of Bibionomorpha (Diptera) with Brachycera-Like Modification of Antennae from Mid-Cretaceous Amber of Myanmar. Insects 2021, 12, 364. https://doi.org/10.3390/insects12040364
Ševčík J, Skartveit J, Krzemiński W, Skibińska K. A Peculiar New Genus of Bibionomorpha (Diptera) with Brachycera-Like Modification of Antennae from Mid-Cretaceous Amber of Myanmar. Insects. 2021; 12(4):364. https://doi.org/10.3390/insects12040364
Chicago/Turabian StyleŠevčík, Jan, John Skartveit, Wiesław Krzemiński, and Kornelia Skibińska. 2021. "A Peculiar New Genus of Bibionomorpha (Diptera) with Brachycera-Like Modification of Antennae from Mid-Cretaceous Amber of Myanmar" Insects 12, no. 4: 364. https://doi.org/10.3390/insects12040364