
The Oldest Evolutionary Lineage of Trichoneura Loew, 1850 (Diptera, Limoniidae) and the First Evidence of This Genus in Cretaceous Spanish Amber


Abstract
:Simple Summary
Abstract

1. Introduction
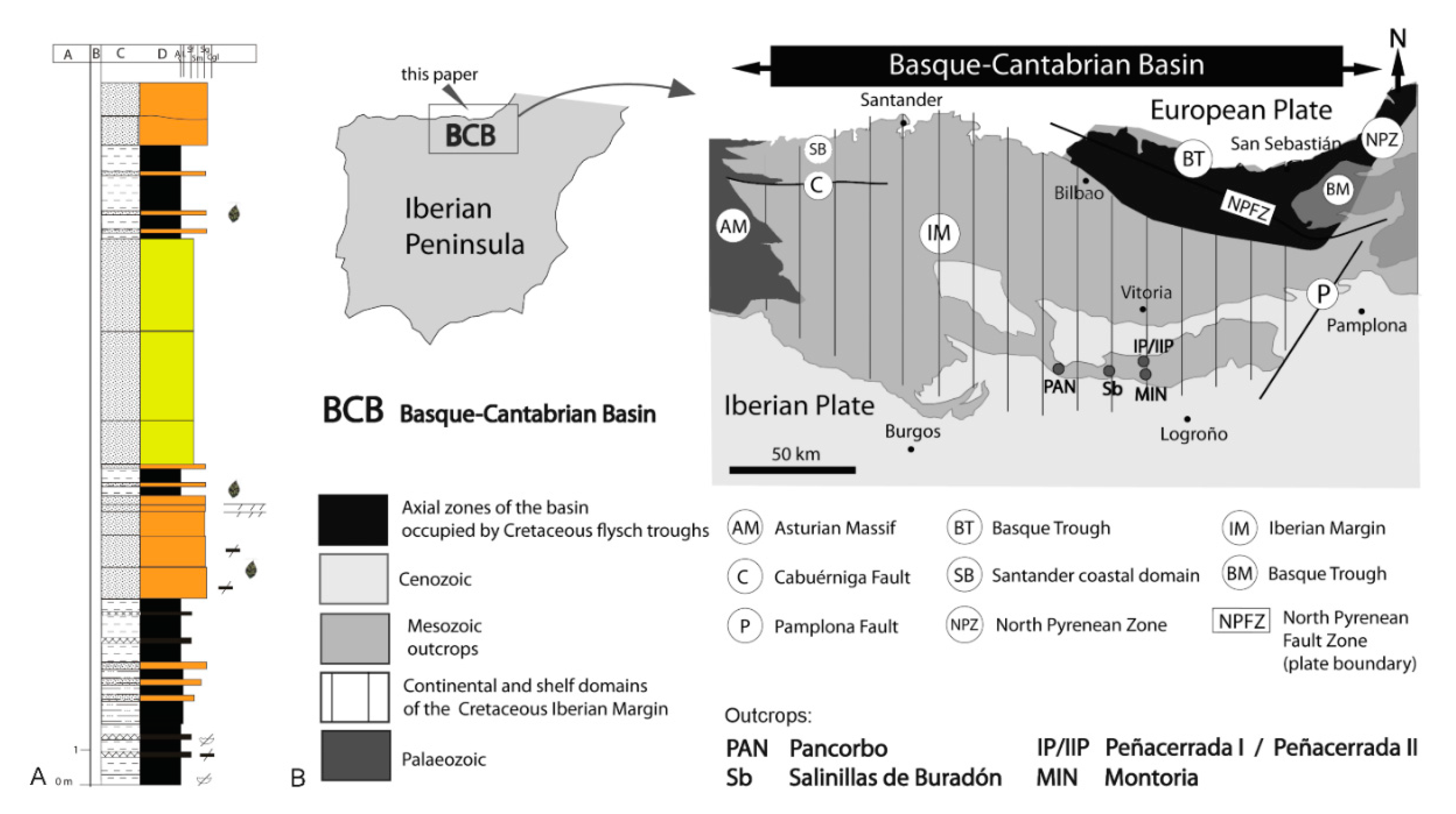
2. Material and Methods
3. Results
3.1. Systematic Paleontology
3.2. Cretalinea subgen. nov.
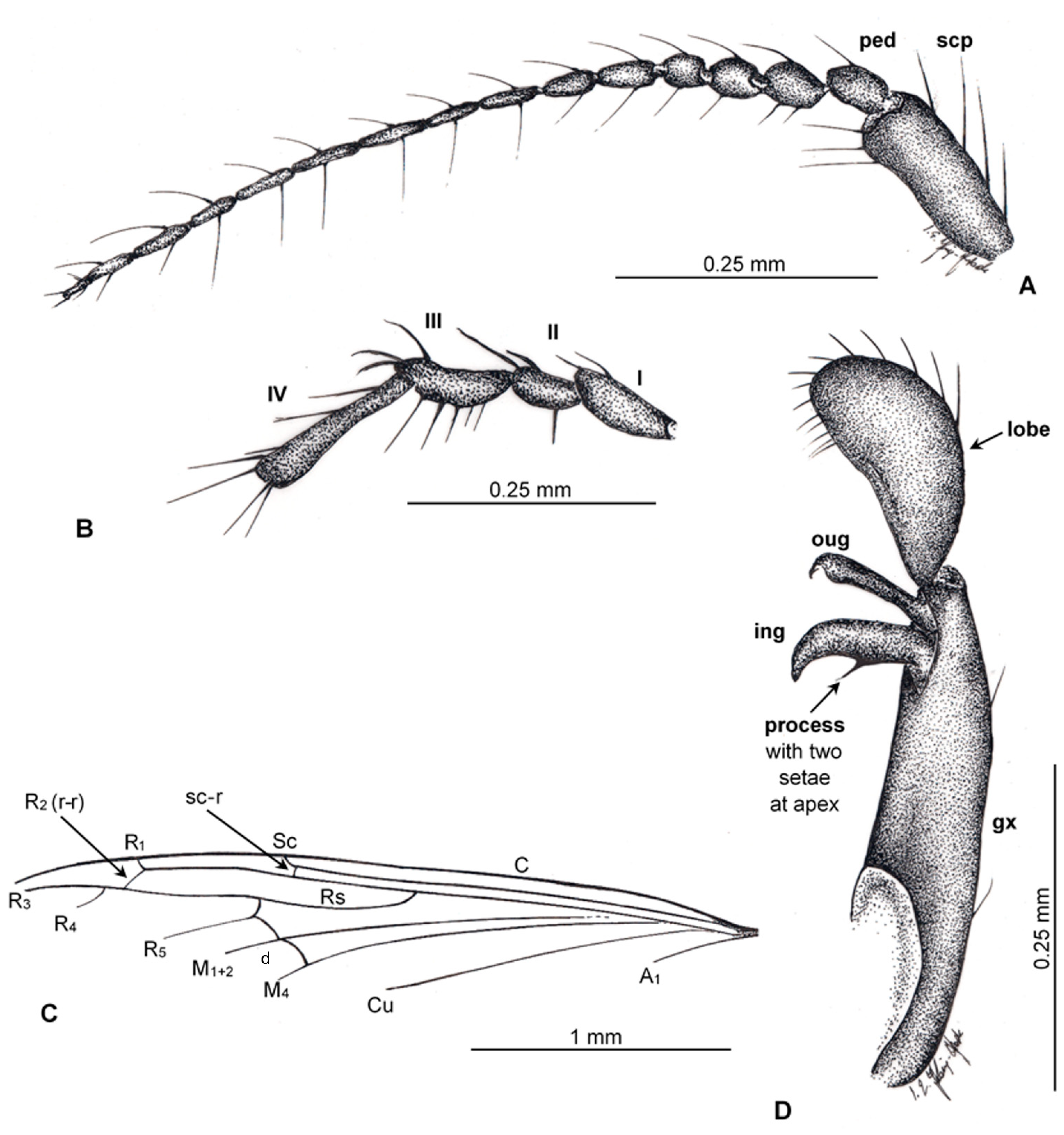
3.3. Trichoneura (Cretalinea) xavieri sp. nov.
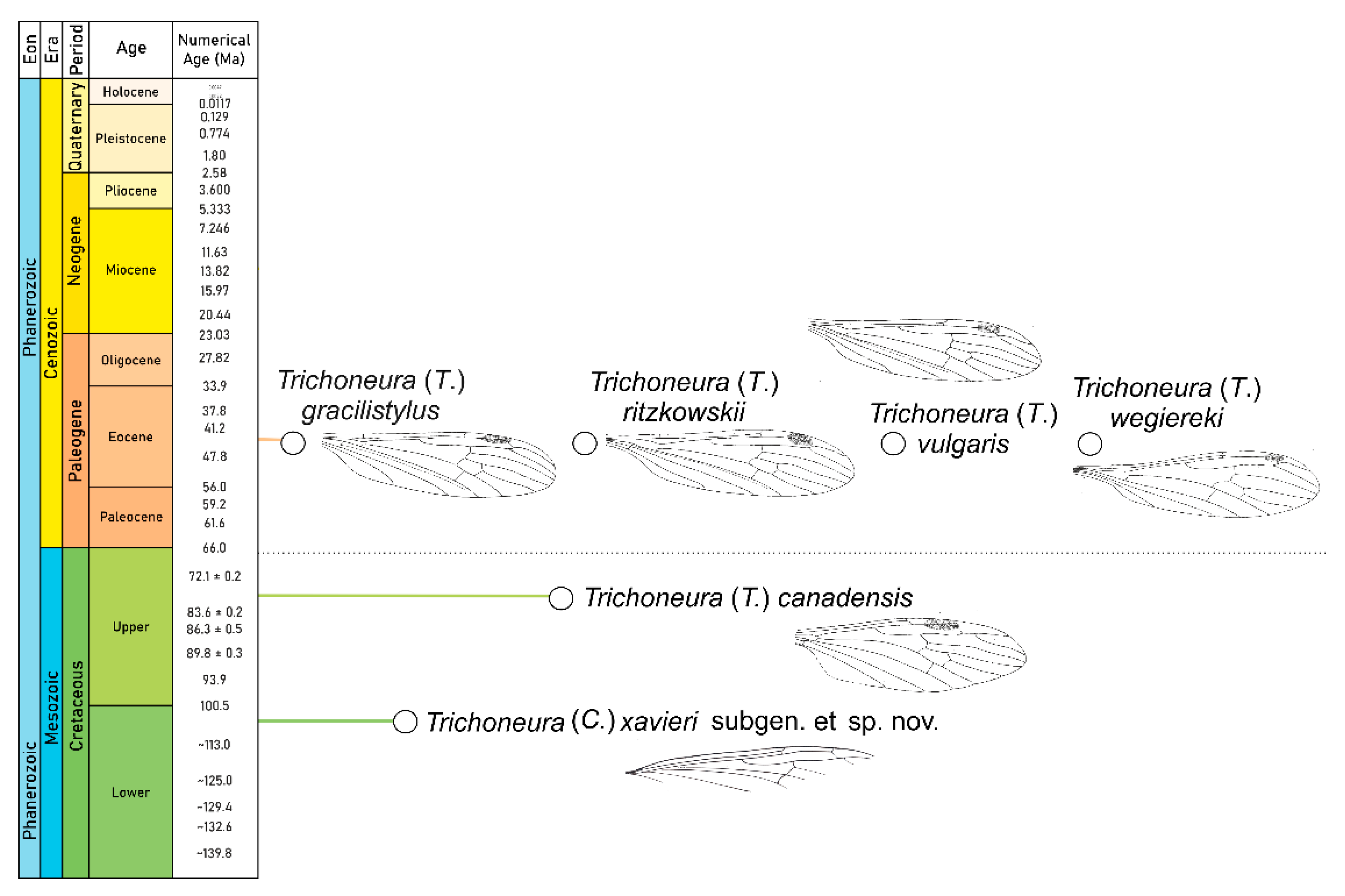
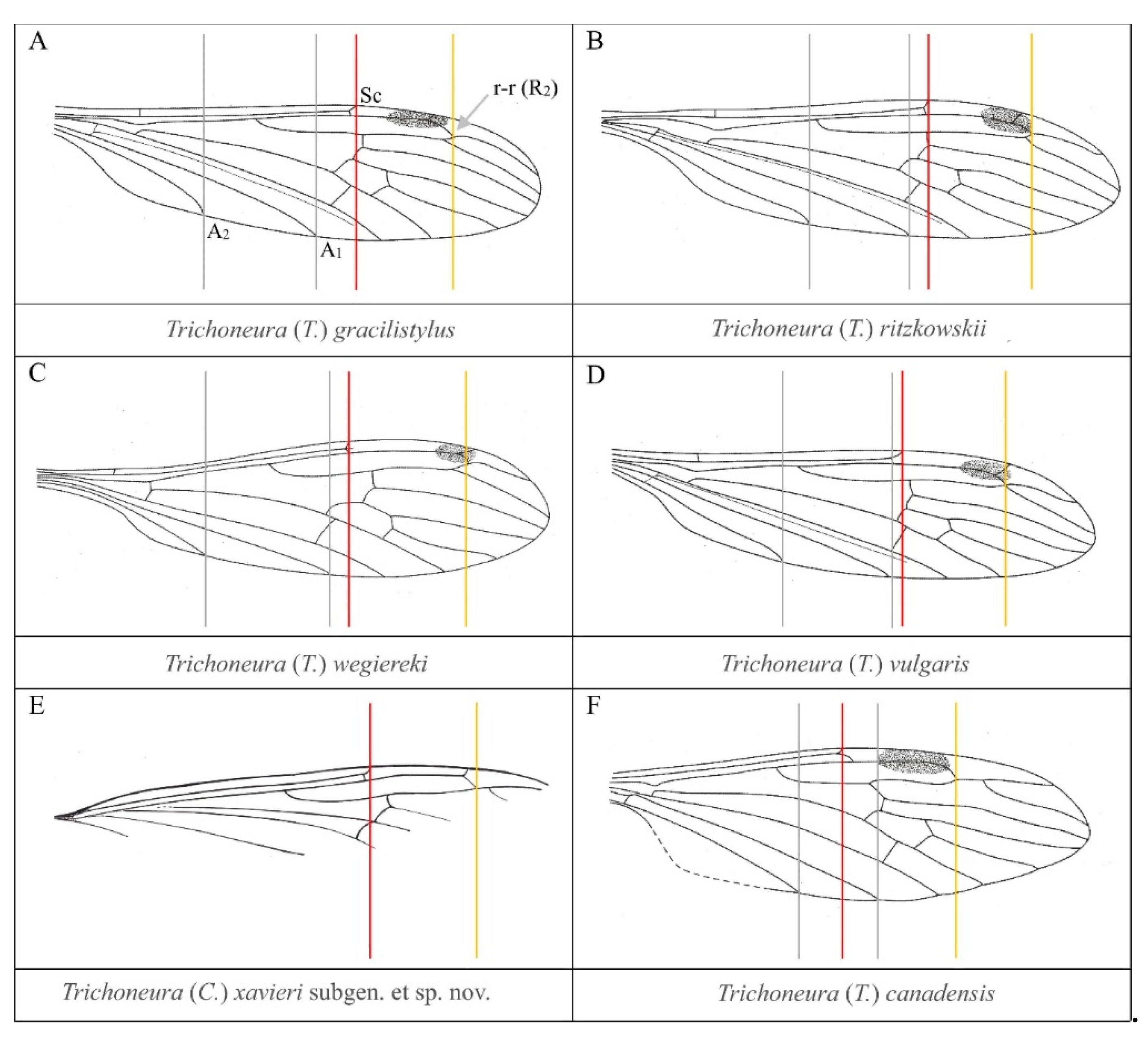
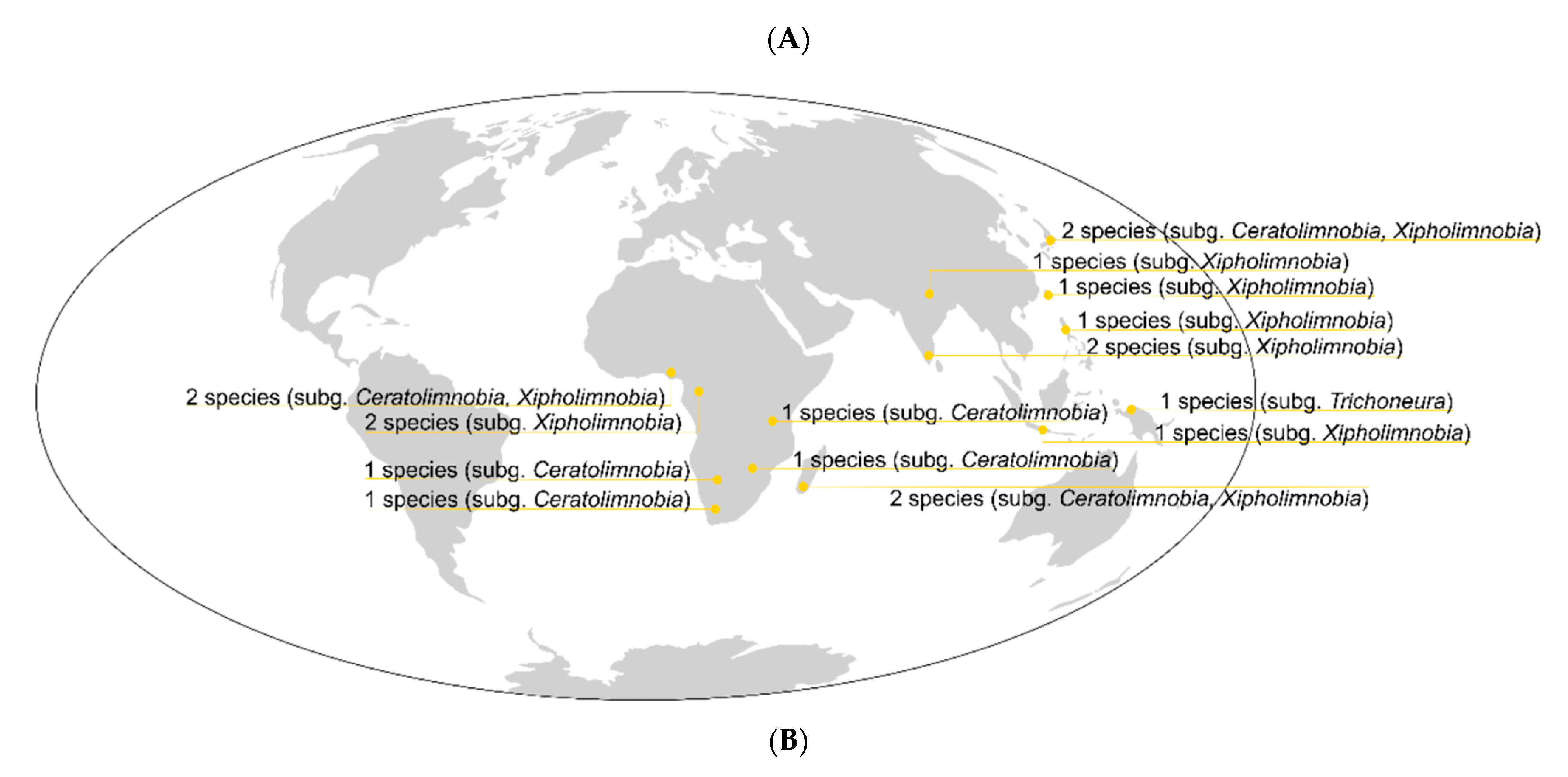
4. Discussion
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Loew, H. Über den Bernstein Und die Bernsteinfauna. Program Keiserischen Realschule Meseritz 1850, 2, 1–44. [Google Scholar]
- Oosterbroek, P. Catalogue of the Crane-Flies of the World. (Diptera, Tipuloidea: Pediciidae, Limoniidae, Cylindrotomidae, Tipulidae). 2000. Last updated 28 December 2012. Available online: http://nlbif.eti.uva.nl/ccw/index.php (accessed on 1 February 2021).
- Kato, D.; Tachi, T. The first records of the tribe Lechriini (Diptera: Limoniidae) in Japan, with descriptions of three new species. Euroasian Entomol. J. 2020, 19, 273–280. [Google Scholar]
- Alexander, C.P. New or little-known Tipulidae (Diptera). III. Ethiopian species. Ann. Mag. Nat. Hist. 1920, 5, 465–472. [Google Scholar] [CrossRef]
- Alexander, C.P. New or little-known Tipulidae (Diptera). IV. Ethiopian species. Ann. Mag. Nat. Hist. 1921, 7, 305–322. [Google Scholar] [CrossRef]
- Krzemiński, W.; Teskey, H.J. New taxa of Limoniidae (Diptera: Nematocera) from Canadian amber. Can. Entomol. 1987, 119, 887–892. [Google Scholar] [CrossRef]
- Alexander, C.P. Crane-flies of the Baltic amber (Diptera). Bernstein Forsch. 1931, 2, 1–135. [Google Scholar]
- Krzemiński, W. Fossil Tipulomorpha (Diptera, Nematocera) from Baltic amber (Upper Eocene): Introductory part: Subfamily Lechriinae (Limoniidae). Polskie Pismo Entomol. 1990, 60, 177–194. [Google Scholar]
- Kania, I. Subfamily Limoniinae Speiser, 1909 (Diptera, Limoniidae) from Baltic amber (Eocene): The genus Trichoneura Loew, 1850. Acta Zool. Crac. 2015, 58, 1–19. [Google Scholar] [CrossRef]
- Corral, J.C.; López del Valle, R.; Alonso, J. El ámbar cretácico de Álava (Cuenca Vasco-Cantábrica, norte de España). Su colecta y preparación. Estud. Mus. Cienc. Nat. Álava 1999, 14, 7–21. [Google Scholar]
- Nascimbene, P.; Silverstein, H. The preparation of fragile Cretaceous ambers for conservation and study of organismal inclusions. Studies on Fossils in Amber, with Particular Reference to the Cretaceous of New Jersey; Grimaldi, D., Ed.; Backhuys Publishers: Leiden, The Netherlands, 2000; pp. 93–102. [Google Scholar]
- McAlpine, J.F.; Peterson, B.V.; Shewell, G.E.; Teskey, H.J.; Vockeroth, J.R.; Wood, D.M. Manual of Nearctic Diptera. In Research Branch Agriculture; Canada Communication Group Pub: Ottawa, ON, Canada, 1981; Volume 1, pp. 27–674. [Google Scholar]
- Krzemiński, W. Three new species of the genus Helius Lepeletier & Serville (Diptera, Limoniidae) from the Middle Miocene of Stavropol (northern Caucasus, Russia). Acta Zool. Crac. 2002, 45, 317–320. [Google Scholar]
- Ribeiro, G.C. Phylogeny of the Limnophilinae (Limoniidae) and early evolution of the Tipulomorpha (Diptera). Invertebrate Syst. 2008, 22, 627–694. [Google Scholar] [CrossRef]
- Barrón, E.; Peyrot, D.; Rodríguez-López, J.P.; Meléndez, N.; López del Valle, R.; Najarro, M.; Rosales, I.; Comas-Rengifo, M.J. Palynology of Aptian and upper Albian (Lower Cretaceous) amber-bearing outcrops of the southern margin of the Basque-Cantabrian Basin (northern Spain). Cretac. Res. 2015, 52, 292–312. [Google Scholar] [CrossRef]
- Linnaeus, C. Systema Nature per Regna tria Naturae, Secundum Classes, Ordines, Genera, Species, cum Caracteribus, Differentiis, Synonymi, Locis, 10th ed.; Laurentii Salvii: Holmiae, UK, 1758; p. 824. [Google Scholar] [CrossRef] [Green Version]
- Rohdendorf, B.B. The oldest infraorders of Diptera from the Triassic of Middle Asia. Palaeontol. Zhurnal 1961, 2, 90–100. [Google Scholar]
- Speiser, P. 4 Orthoptera. Orthoptera Nematocera. Wissenschaftliche Ergebnisse der Schwedischen Zoologische Expededition nach Kilimandjaro. Meru 10 Diptera 1909, 31–65. [Google Scholar]
- Alexander, C.P. Fauna Sumatrensis (Beitrag Nr. 39). Superfamily Tipuloidea (Dipt.) I. Suppl. Entomol. 1927, 15, 90–102. [Google Scholar]
- Gavryushin, D.I. Six new species of limoniid flies (Diptera: Limoniidae) from Tanzania and notes on other species new to the country’s fauna. Russ. Entomol. J. 2016, 25, 273–286. [Google Scholar] [CrossRef] [Green Version]
- Peñalver, E.; Delclòs, X. Spanish Amber. In Biodiversity of Fossils in Amber from the Major World Deposits; Penney, D., Ed.; Siri Scientific Press: Manchester, UK, 2010; pp. 236–270. [Google Scholar]
- Maksoud, S.; Azar, D. Lebanese amber: Latest updates. Palaeoentomology 2020, 3, 125–155. [Google Scholar] [CrossRef]
- Shi, G.; Grimaldi, D.A.; Harlow, G.E.; Wang, J.; Wang, J.; Yang, M.; Lei, W.; Li, Q.; Li, X. Age constraint on Burmese amber based on U-Pb dating of zircons. Cretac. Res. 2012, 37, 155–163. [Google Scholar] [CrossRef]
- Le Peletier, A.L.M.; Serville, J.G.A. Entomologie, ou histoire naturelle des crustacés, des arachnides et des insectes. Encycl. Method. Hist. Nat. 1828, 10, 345–833. [Google Scholar]
- Krzemiński, W.; Kania, I.; Azar, D. The Early Cretaceous evidence of rapid evolution of the genus Helius Lepeletier and Serville, 1828 (Limoniidae, Diptera). Cretac. Res. 2014, 48, 96–101. [Google Scholar] [CrossRef]
- Kania, I.; Krzemiński, W.; Azar, D. The oldest representative of Helius Lepeletier & Serville 1828 (Limoniidae, Diptera) from Lebanese amber (Early Cretaceous). Insect Syst. Evol. 2013, 44, 1–8. [Google Scholar]
- Krzemiński, W.; Arillo, A. Alavia neli, n. gen. and n. sp.—The first Limoniidae (Diptera) from the Lower Cretaceous amber of Álava (Spain). Alavesia 2007, 1, 11–13. [Google Scholar]
- Kania, I.; Krzemiński, W.; Arillo, A. First representative of the genus Helius Lepeletier and Serville, 1828 (Diptera, Limoniidae) from the Lower Cretaceous Álava amber (Spain). Cretac. Res. 2016, 63, 33–38. [Google Scholar] [CrossRef]
- Kania, I.; Krzemiński, W.; Arillo, A. A new peculiar species of the genus Helius Lepeletier & Serville, 1828 (Diptera, Limoniidae) from Cretaceous Álava amber (Spain). Earth Environ. Sci. Trans. R. Soc. Edinb. 2017, 107, 1–7. [Google Scholar]
- Latreille, P.A. Histoire Naturelle, Generale et Particuliere, des Crustaces et des Insectes. Ouvrage Faisant Suite a l Histoire Naturelle Generale et Particuliere, Composee par Leclerc de Buffon, et Redigee par C.S. Sonnini, Membre de Plusieurs Societes Savantes. Tome troisieme. Familles Naturelles des Genres; Dufart: Paris, France, 1802; Volume i–xii, pp. 13–467. [Google Scholar]
- Azar, D.; Gèze, R.; Acra, F. Lebanese Amber. In Biodiversity of Fossils in Amber from the Major World Deposits; Penney, D., Ed.; Siri Scientific Press: Manchester, UK, 2010; pp. 271–293. [Google Scholar]
- Doria, G.; Royer, D.L.; Wolfe, A.P.; Fox, A.; Westgate, J.W.; Bereling, D.J. Declinic atmospheric CO2 durring the Late Middle Eocene climate transitin. Am. J. Sci. 2011, 311, 63–75. [Google Scholar] [CrossRef]








{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
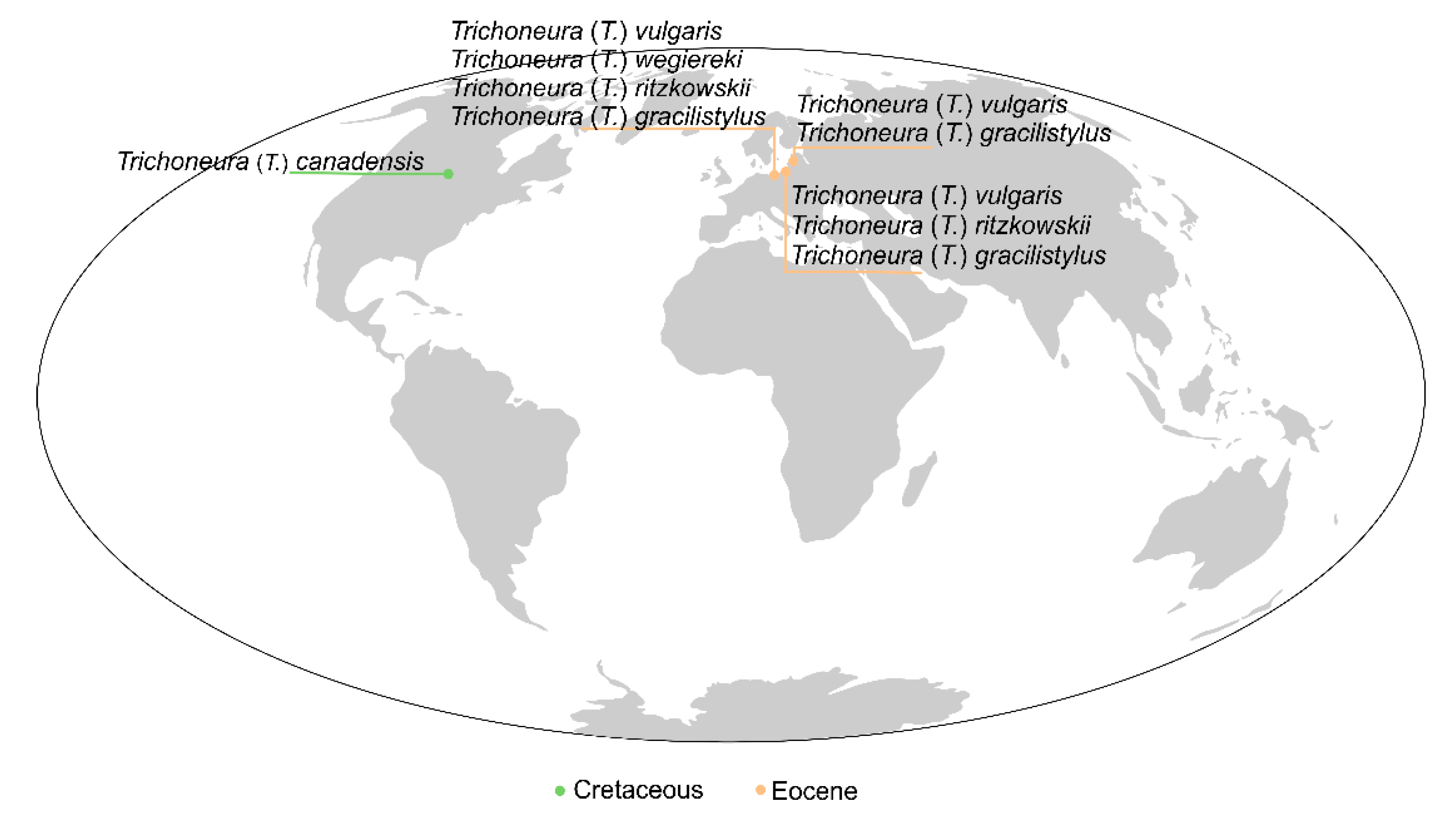
| Species | Time Scale | Type of Material | Locality |
|---|---|---|---|
| Trichoneura (Trichoneura) gracilistylus Alexander, 1931 [7] | Eocene | Baltic amber | Baltic area |
| Trichoneura (Trichoneura) ritzkowskii Krzemiński, 1990 [8] | Eocene | Baltic amber | Baltic area |
| Trichoneura (Trichoneura) wegiereki Kania, 2015 [9] | Eocene | Baltic amber | Baltic area |
| Trichoneura (Trichoneura) vulgaris Loew, 1850 [1] | Eocene | Baltic amber | Baltic area |
| Trichoneura (Trichoneura) canadensis Krzemiński et Teskey, 1987 [6] | Upper Cretaceous | Canadian amber | Canada |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Kania-Kłosok, I.; Krzemiński, W.; Kopeć, K.; Arillo, A. The Oldest Evolutionary Lineage of Trichoneura Loew, 1850 (Diptera, Limoniidae) and the First Evidence of This Genus in Cretaceous Spanish Amber. Insects 2021, 12, 411. https://doi.org/10.3390/insects12050411
Kania-Kłosok I, Krzemiński W, Kopeć K, Arillo A. The Oldest Evolutionary Lineage of Trichoneura Loew, 1850 (Diptera, Limoniidae) and the First Evidence of This Genus in Cretaceous Spanish Amber. Insects. 2021; 12(5):411. https://doi.org/10.3390/insects12050411
Chicago/Turabian StyleKania-Kłosok, Iwona, Wiesław Krzemiński, Katarzyna Kopeć, and Antonio Arillo. 2021. "The Oldest Evolutionary Lineage of Trichoneura Loew, 1850 (Diptera, Limoniidae) and the First Evidence of This Genus in Cretaceous Spanish Amber" Insects 12, no. 5: 411. https://doi.org/10.3390/insects12050411