Beetle Species–Area Relationships and Extinction Rates in Protected Areas


Department of Life, Health and Environmental Sciences, University of L’Aquila, 67100 L’Aquila, Italy
Insects 2020, 11(9), 646; https://doi.org/10.3390/insects11090646
Submission received: 21 August 2020
/
Revised: 17 September 2020
/
Accepted: 17 September 2020
/
Published: 21 September 2020
Abstract
:Simple Summary
Larger areas tend to host more species. This general ecological pattern (known as the species–area relationship, SAR) can be used to calculate expected extinction rates following area (habitat) loss. Here, using data from Italian reserves, SAR-based extinction rates are calculated for beetle groups with different ecology: terrestrial predators, aquatic predators, dung feeders, herbivores, and detritivores. Reserve area was an important predictor of species richness in all cases. However, also other factors besides area were important correlates of species richness. For some groups, species richness tends to decline with elevation and/or northwards. Extinction rates are higher for dung beetles, due to their dependence on large grazing areas, and detritivores, due to their low dispersal capabilities, which reduce their ability to reach new places when environmental conditions became less favorable. The lower extinction rates predicted for other groups can be explained by their higher dispersal ability. Extinction rates by area loss are always relatively low. This means that, in reserves with few species, many extinctions might be unnoticed.
Abstract
The species–area relationship (SAR, i.e., the increase in species richness with area) is one of the most general ecological patterns. SARs can be used to calculate expected extinction rates following area (habitat) loss. Here, using data from Italian reserves, extinction rates were calculated for beetle groups with different feeding habits: Carabidae (terrestrial predators), Hydradephaga (aquatic predators), coprophagous Scarabaeoidea (dung feeders), phytophagous Scarabaeoidea (herbivores), and Tenebrionidae (detritivores). The importance of other factors besides area (namely latitude and elevation) was investigated. Reserve area was recovered as an important predictor of species richness in all cases. For Carabidae, Hydradephaga, and Tenebrionidae, elevation exerted a negative influence, whereas latitude had a negative influence on coprophagous Scarabaeoidea and Tenebrionidae, as a consequence of current and historical biogeographical factors. Extinction rates were higher for dung beetles, due to their dependence on large grazing areas, and Tenebrionidae, due to their low dispersal capabilities. The lower extinction rates predicted for Carabidae, phytophagous Scarabaeoidea, and Hydradephaga can be explained by their higher dispersal power. If other variables besides area are considered, extinction rates became more similar among groups. Extinction rates by area loss are always relatively low. Thus, in reserves with few species, many local extinctions might be unnoticed.
1. Introduction
The species–area relationship (SAR, i.e., the increase in species richness with area) is an almost ubiquitous ecological pattern [1]. There is increasing interest in how the SAR can be used for conservation purposes, including the selection of biodiversity hotspots [2,3,4], the identification of the best size and shape of natural areas [5,6,7,8], and the prediction of species extinction [9,10]. There is a debate about how accurate species extinction rates based on the SAR are [10]. Although some analyses suggest that SAR-based extinction rates are overestimated, empirical data indicate that the SAR probably underestimates, not overestimates species extinctions [9,10].
The basic idea in the use of the SAR to predict species loss is that a reduction in area size implies a reduction in species number. Thus, the same function which is used to model the SAR can be used, in a reverse way, to predict species loss. Empirical evidence largely supports that, at least for isolates, most systems are adequately modelled by the power function (1):
where S is the species number, A is area, and c and z are fitting parameters [11,12]. The power function is frequently applied in its linearized (log-log) form using decimal logarithms (2):
S = cAz
log(S) = log(c) + z log (A)
In this form, c is the expected number of species per area unit, and z is the slope of the function [13].
The use of the SAR to predict species extinction is based on the assumption that if the original area A0 is reduced to A1, the original number of species S0 is expected to decline to S1, according to the following Equation (3) [10]:
S1 = S0 (A1/A0)z
Using this approach, it is possible to predict extinction rates in isolated blocks of fragmented habitats, including protected areas, following area loss [14,15,16].
The Italian peninsula is located in the center of the Mediterranean Basin, one of the world’s hotspots of biodiversity [8,17,18], showing exceptionally high levels of diversity for a variety of plant and animal taxa [19,20]. At the same time, due to the profound, millenary impacts on the wilderness of this area, the Mediterranean biodiversity has been strongly influenced by the human presence, and it is currently under many threats [21,22].
According to the last official report, in Italy there are some 900 protected areas, occupying a terrestrial surface of about three million hectares (about 11% of the country) [23], plus about other 400 areas that benefit from some form of protection, for a further 430,000 hectares [24]. The number and size of protected areas is, however, under continuous change.
Biodiversity knowledge for most of the Italian protected areas is extremely poor, especially for invertebrates. However, for at least some groups, there is information about the number of species present in some reserves. In particular, thanks to the interest of professional and amateur entomologists, some beetle groups are among the few invertebrates for which it is possible to gather reliable values of species richness for sets of Italian reserves. Beetles are the most diversified group of living organisms, constituting about 40% of all described insect species and 25% of all known life-forms [25,26]. Beetles occur in most terrestrial and freshwater habitats and are ecologically extremely diversified, which makes these insects very useful in comparative analyses: for example, with reference to their trophic habits, dispersal capabilities, and ecological preferences.
In this paper, I used data on beetle groups characterized by very different ecology (terrestrial predators, aquatic predators, dung feeders, herbivores, and terrestrial detritivores) from Italian reserves to investigate how extinction rates based on the use of the SAR vary according to the beetle’s ecology.
2. Materials and Methods
I collected literature data on species richness for Carabidae, Hydradephaga, Scarabaeoidea, and Tenebrionidae from Italian reserves. Carabidae are a family of mainly terrestrial predators [27], whereas Hydradephaga are a group of families (Gyrinidae, Haliplidae, Noteridae, Dytiscidae, Hygrobiidae for the Italian fauna) of freshwater predators [28,29]. Scarabaeoidea, as intended here, include Trogidae, Bolboceratidae, Geotrupidae, Hybosoridae, Ochodaeidae, Glaphyridae, and Scarabaeidae (including Aphodiinae, Scarabaeinae, Orphninae, Melolonthinae, Rutelinae, Dynastinae, and Cetoniinae), but not Lucanidae [30]. The Scarabaeoidea include species with a wide spectrum of feeding habits, but most of the species are either coprophagous (dung beetles) or phytophagous [30]. For this reason, I conducted analyses for the whole group and for the coprophagous and phytophagous species separately (the whole group analyses included more species than the sum of coprophagous and phytophagous because of the presence of species that did not belong to these two categories, for example mycetophagous species). Finally, Tenebrionidae are essentially saprophagous insects [27]. Although the Alleculinae (formerly considered a separate family) fall within the tenebrionid cladogenesis, I did not consider them in the analysis because of their extremely different and highly derived ecological characteristics (they are the only flower visiting tenebrionids and, contrary to most tenebrionids, they are flying insects) [31]. Additionally, for most reserves, there is no information on alleculines. I also omitted the genus Lagria (another flower visiting tenebrionid) and the genus Myrmechixenus, since its distribution is very poorly known. I also omitted synanthropic species [31].
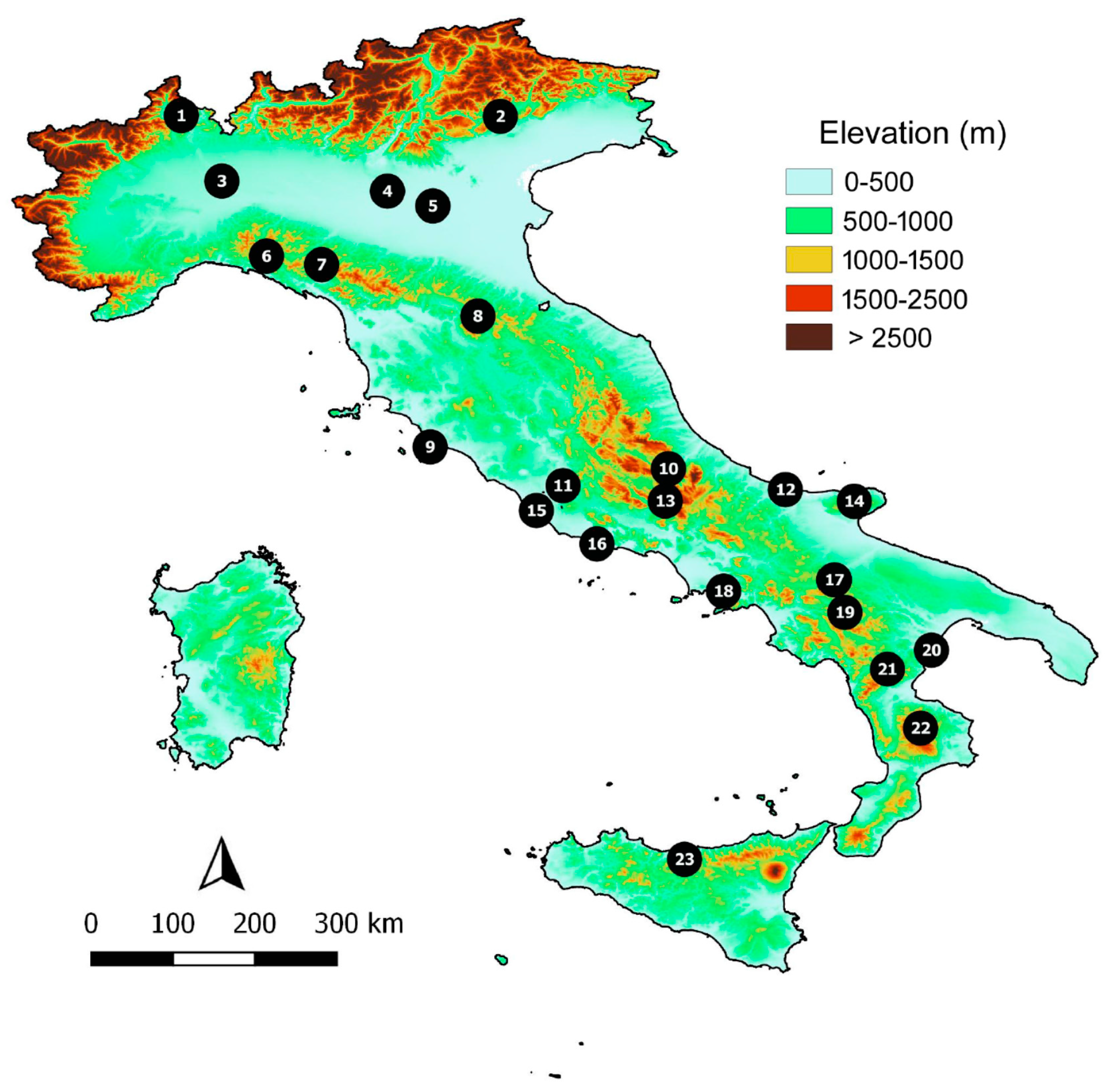
Overall, I collected data from 23 Italian reserves (18 reserves for Carabidae [32,33,34,35,36,37,38,39,40,41,42,43,44,45], 14 reserves for Hydradephaga [32,33,35,46,47,48,49,50], 18 reserves for Scarabaeoidea [32,33,35,37,49,51,52,53,54,55,56,57,58,59], and 18 reserves for Tenebrionidae [32,33,35,37,54,60,61,62,63,64,65,66,67,68,69,70,71] (Figure 1). Contiguous reserves were considered to a be a single reserve. Values of species richness for the various beetle groups in each reserve are given in Table 1.
For reserve areas, where possible, I referred to the official size. Some study areas did not correspond exactly to the borders of a reserve. This occurred when a study area was only a part of a larger reserve (reserve 22), when the study area was the sum of contiguous reserves (reserves 14 and 15), or when the study area included adjacent areas (reserves 10, 13, 23). In such cases, I used as the size of the study area the surface reported in the reference used to extract the faunal data; if not given, I calculated this surface from the maps provided by the authors of the faunal studies.
The selected reserves ranged from 0.17 to 1700 km2, thus spanning for four orders of magnitude. This avoided the risk of the SAR appearing to be linear because of sampling a small range of areas [72]. Reserves were distributed throughout the Italian peninsula and Sicily to fully encompass the Italian latitudinal gradient. Reserves were also representative of coastal, internal plain, and montane landscapes.
SARs were first modelled using the linearized version of the power function (Equation (2)) with log10. Values of c and z for the SAR of the various groups were compared using Analysis of Covariance (ANCOVA) [13]. z-values of SARs were used to calculate the expected number of species lost with increasing area loss using Equation (3).
Although this is the standard procedure to infer potential species loss due to area reduction, the number of species is not controlled only by area. Even when area is the most important determinant of species richness, other variables may exert a relevant role. Elevation is an important macroecological variable that influences species richness [73]. To take into account the role of elevation, for each area, I considered various measures of elevation: minimum, maximum, average, and range. Elevational range was considered as a proxy of environmental variability. Mean elevation was intended to reflect the overall orographic physiognomy of a given area, whereas minimum and maximum elevations were introduced because extreme values are clearly distinctive of coastal and montane areas.
When elevation of sampling sites was reported in the examined faunal studies, their values were used to calculate minimum elevation, maximum elevation, elevational range (maximum minus minimum) and mean elevation for the study areas (reserve 1 [43], reserves 6 and 7 [74], reserve 14 [75], reserve 18 [76], reserve 21 [77], reserve 22 [78]). For reserve 8, I used elevation of sampling points reported by Lestes [36] for Carabidae and by Rocchi and Mascagni [50] for Hydradephaga. For reserve 5, I referred to the minimum, maximum, and mean values reported by the authors of the faunal study [54]. For reserve 23, I calculated the average elevation using minimum and maximum values as given by Aliquò and Leo [60], integrated with topographic maps. In other cases, I calculated these values from topographic maps.
Due to the peculiar, north–south orientation of Italy, study areas are distributed along a latitudinal gradient. It is well known that species richness of most taxa shows a distinct latitudinal pattern, decreasing (in the northern hemisphere) from south to north [79,80,81,82,83]. This might influence the SAR by negatively affecting species richness in northern areas. For this reason, I also considered the latitude (measured in decimal degrees) of the reserve centroid as a further correlate. All variables were log10-transformed prior to analyses; a log10(x+1) transformation was used to manage zero values for coprophagous Scarabaeoidea (because of the presence of a reserve with no species) and minimum elevation (for coastal areas with a minimum elevation of 0 m).
To evaluate the relative importance of elevation (minimum, maximum, mean, and range) and latitude in determining species richness besides reserve area, I used a multimodel selection procedure in which all variables were tested individually and in all their possible combinations. Then, models were ordered by decreasing values of the small-sample corrected Akaike Information Criterion (AICc), and the model with the lowest AICc was selected as the best model; alternative models with ΔAIC values ≤ 2 were considered as equally supported [84]. When the best fit model included other variables than area, the coefficient of area in the multiple model was used as a z-value for the estimation of species loss in equation 3. In practice, this approach consists in predicting species loss by area reduction, holding stable the influence of all other variables with an exponent z that, however, takes into account their influence in species richness. This is reasonable in ecological terms, since the surface of a certain study area suitable for a certain group of species can be reduced by habitat fragmentation, loss, or alteration, whereas its geographical position and orography remain unchanged.
3. Results
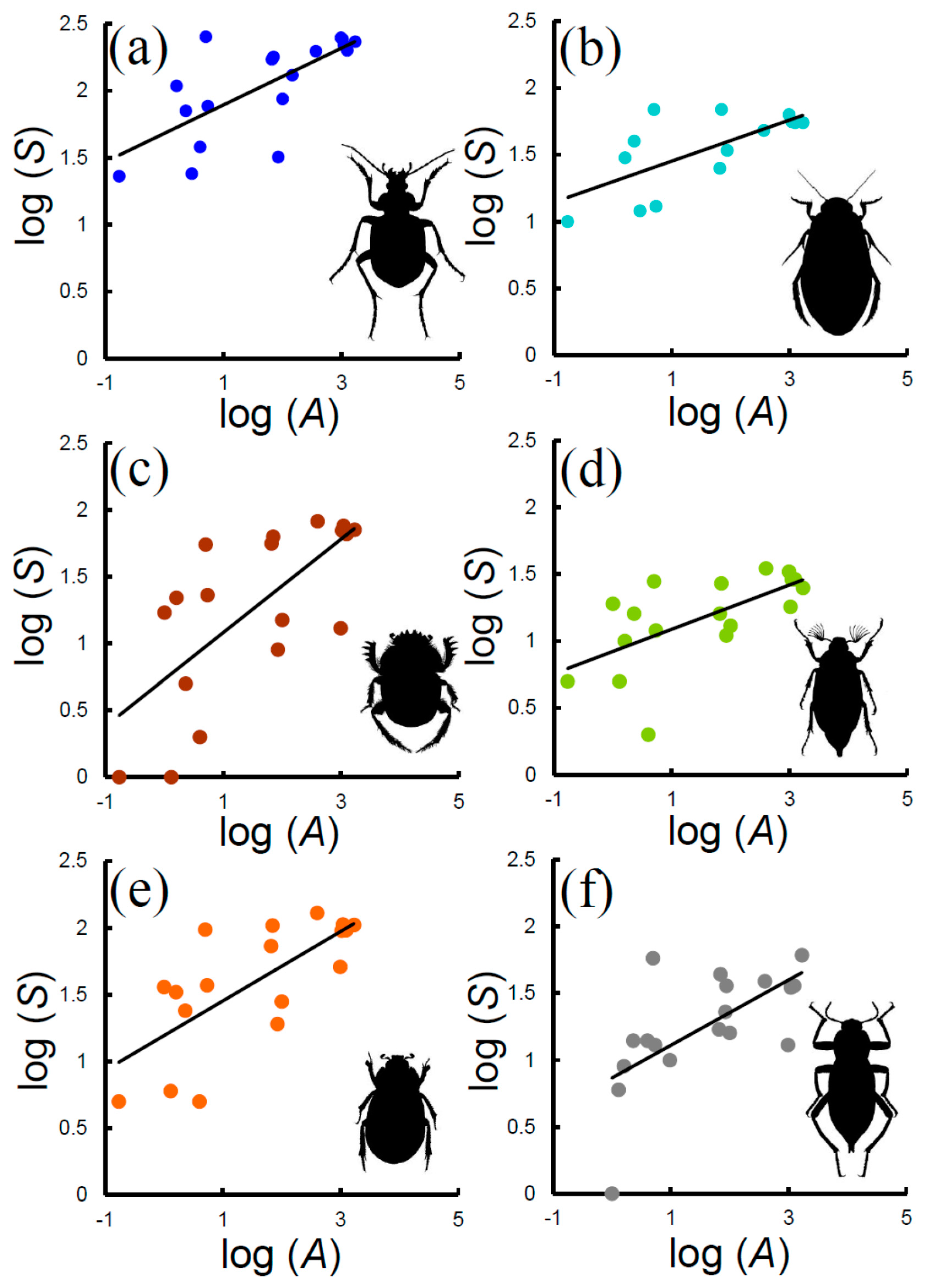
The power function modelled SARs of the various groups with very similar goodness of fit (Figure 2, Table 2). z-values, which indicate the rate at which species accumulate with area, did not vary significantly between groups (ANCOVA: F = 1.536, P = 0.200). By contrast, ANCOVA indicated an overall significance difference between c-values (F = 18.72, P << 0.001). c-values, which represent the expected number of species per area unit, decreased in the following order (~indicates that differences in c-values were not significant in pairwise ANCOVAs): Carabidae (48 species) < Hydradephaga (20) < phytophagous Scarabaeoidea (8) ~ Tenebrionidae (7) ~ coprophagous Scarabaeoidea (5).
The results of multimodel selection (Table 3) showed that area was an important predictor in all cases and, for both total and phytophagous Scarabaeoidea, it was the only variable included in the best fit models.
For Carabidae, two equally supported models (ΔAICc = 1.13) were selected as the best fit models: the first model included area (positively) and elevational range (negatively, but not significantly); the second model included only area (positively). For Hydradephaga, the best fit model included area (positively) and elevational range (negatively). For coprophagous Scarabaeoidea, the best fit model included area (positively) and latitude (negatively). For coprophagous and all Scarabaeoidea, the best fit models included only area (positively). Finally, for Tenebrionidae, the best fit model included area (positively), latitude (negatively), and minimum elevation (negatively).
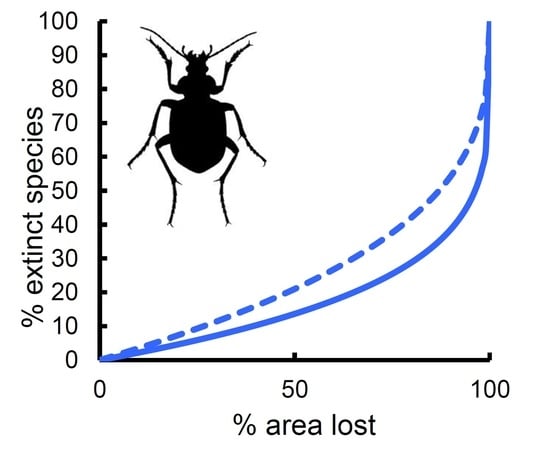
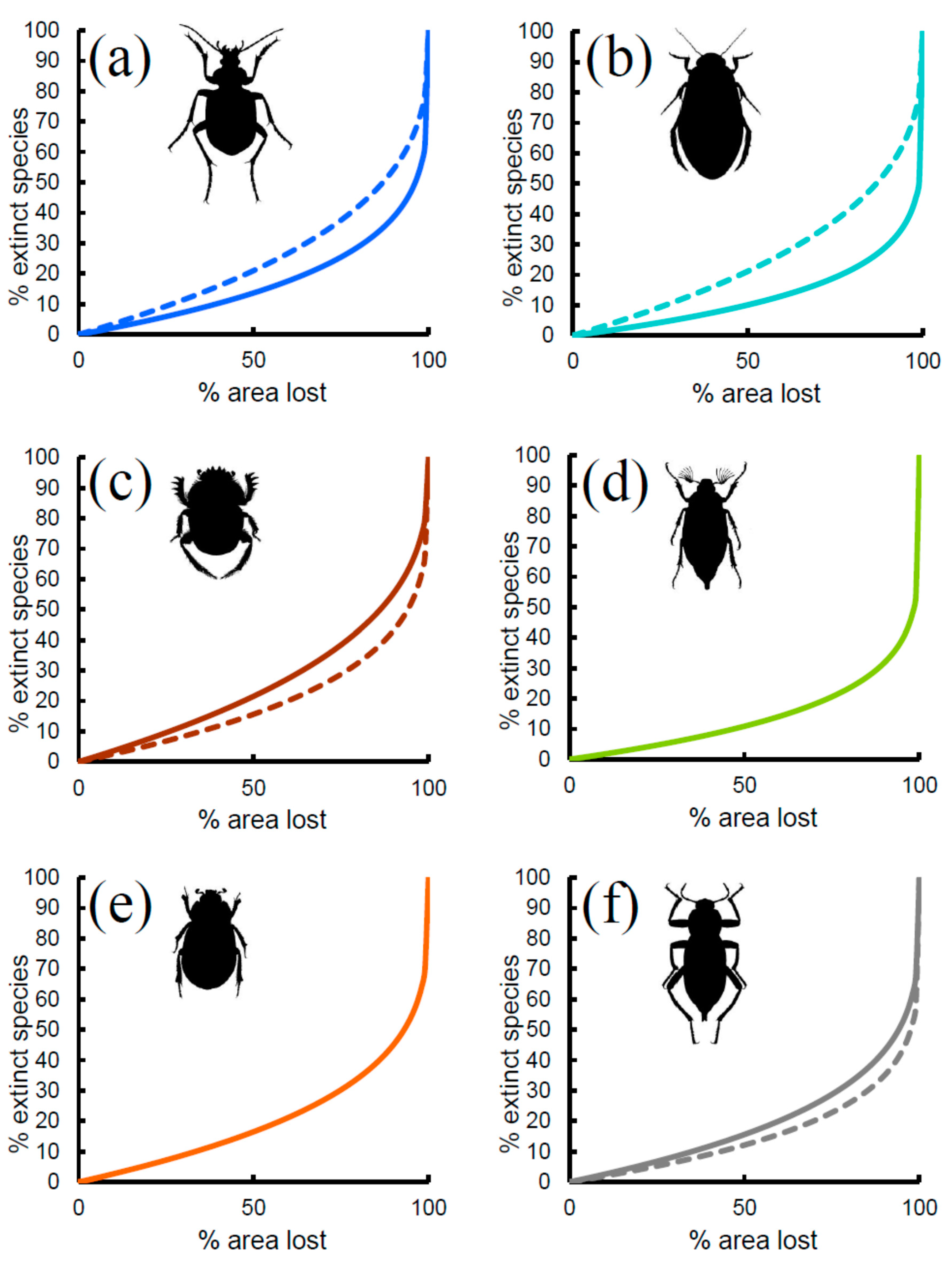
Predicted rates of species extinction based on the z-values of the power function (Figure 3, solid lines) showed similar patterns among groups, with small proportions of extinct species until the fraction of lost area was very large. The most sensitive group was the coprophagous Scarabaeoidea, for which a loss of 25% of area would lead to the local extinction of about 9% of the species (13% of species with 33% of area lost, and 21% with 50% of area lost, respectively). Tenebrionidae would lose about 7% of species if 25% of area was lost (9% of species with 33% of area lost, and 15% with 50% of area lost, respectively). Total Scarabaeoidea showed a very similar trend. Carabidae would lose about 6% of species if 25% of area was lost (8% of species with 33% of area lost, and 13% with 50% of area lost, respectively). Phytophagous Scarabaeoidea would lose about 5% of species if 25% of area was lost (6% of species with 33% of area lost, and 11% with 50% of area lost, respectively), and a virtually identical pattern was found for the Hydradephaga. Thus, sensitivity to extinction by area reduction increased in the order: coprophagous Scarabaeoidea < Tenebrionidae < Carabidae < phytophagous Scarabaeoidea < Hydradephaga.
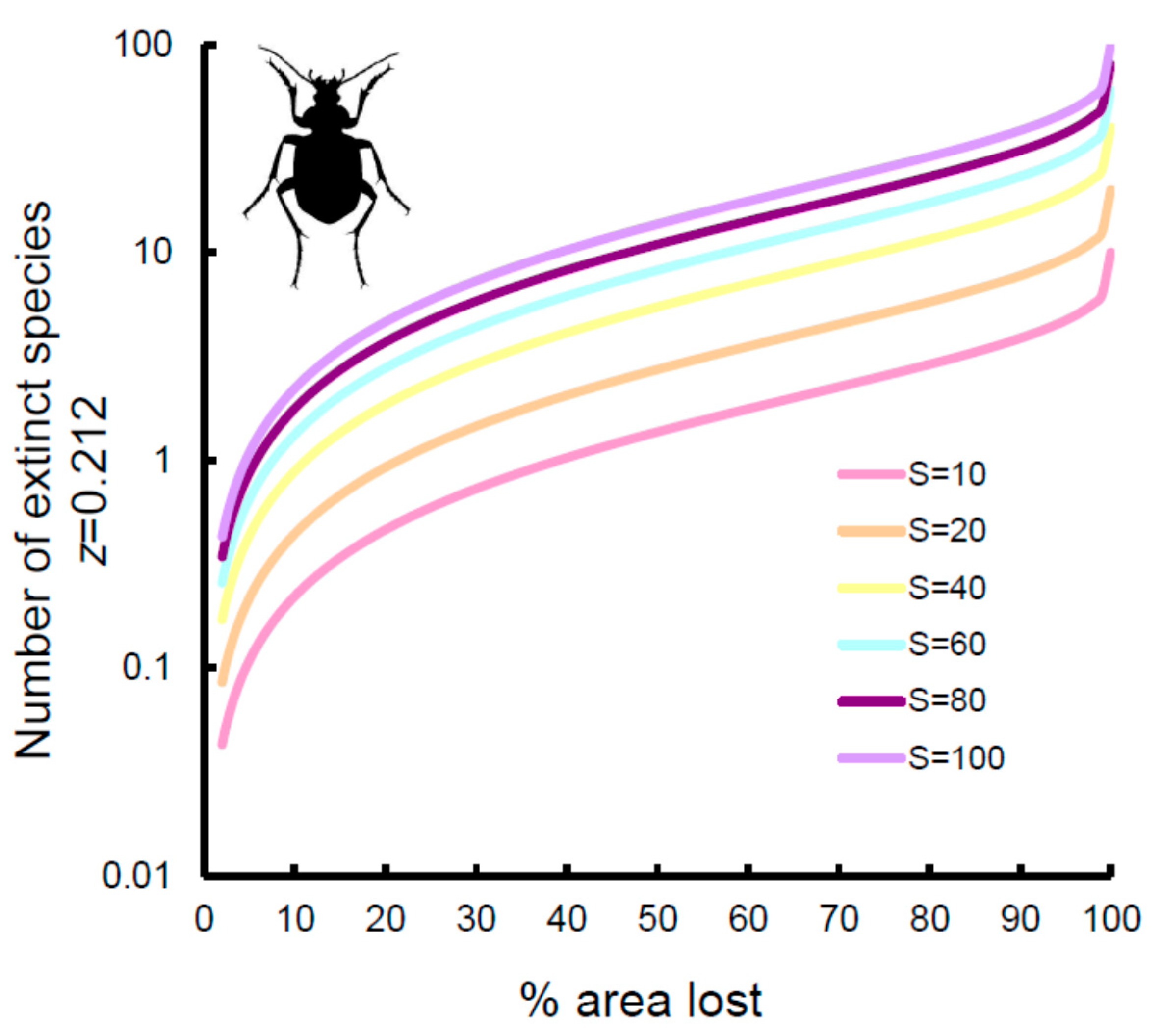
If the coefficient for area obtained from the multiple regression models was used (Figure 3, dashed lines), extinction rates increased substantially for Carabidae and Hydradephaga, with virtually identical trends (9% of species with 25% of area lost, 12% of species with 33% of area lost, and 21% with 50% of area lost, respectively). By contrast, extinction rates decreased slightly for coprophagous Scarabaeoidea (7% of species with 25% of area lost, 9% of species with 33% of area lost, and 15% with 50% of area lost, respectively) and Tenebrionidae (5% of species with 25% of area lost, 7% of species with 33% of area lost, and 12% with 50% of area lost, respectively). However, because of the relatively small number of species occurring in each reserve, extinction might be difficult to detect. As shown in Figure 4 for Carabidae (other groups had virtually identical patterns; the use of z-values from multiple model also did not change the patterns; see Figure S1), with ten species, an area loss of more than 25% is necessary to observe one extinction, whereas with 100 species, a loss of 3% is sufficient to have one extinction. A reduction of 33% of area would still translate to the loss of one species in a reserve containing 10 species, 5 species in a reserve with 40 species, and 13 species in a reserve with 100 species. This suggests that extinctions might be hardly detectable, even for a very large loss of area, if the reserve is small and contains few species.
4. Discussion
The z-values of the power function SARs found in this study (z = 0.15–0.35) were perfectly in the range typically found for isolates (z = 0.2–0.4; [11,12]). The c-values indicate that Carabidae had much more species per area unit than any other group. The second group with the highest number of species per area unit is the Hydradephaga. Phytophagous and coprophagous Scarabaeoidea and Tenebrionidae have similar c-values, much lower than those for Carabidae and Hydradephaga. It is an unexpected result that predators have more species than dung feeders, herbivores, and detritivores. However, it is known that c-values reflect species dispersal abilities, being larger for groups with higher dispersal abilities [13]. Thus, it is possible that the high number of species per area unit in Carabidae and Hydradephaga reflect their higher dispersal abilities. Carabids are known to have, in general, high dispersal capabilities, especially the macropterous forms [87,88,89]. Similarly, many Hydradephaga are known to fly for long distances and move between waterbodies, which is essential for animals that must survive in lentic waterbodies subject to seasonal drought [90]. By contrast, the small number of tenebrionid species per area unit can be related to the typically low dispersal power of these mainly flightless beetles [91,92,93]. The small value of species richness per area unit of phytophagous Scarabaeoidea can be interpreted as a result that some species are associated with some plants or forms of vegetation, and hence high values of species richness can be found only in complex landscapes. Coprophagous Scarabaeoidea depend on the presence of dung, which is in turn typically associated with the presence of cattle; this means that large grazing areas are needed to host a rich fauna. The importance of area for both coprophagous and phytophagous Scarabaeoidea is further shown by the fact that this variable overwhelms the contribution of all other tested variables, being the only one retained in the multimodel selection procedure.
For Hydradephaga and possibly Carabidae, in addition to area, an important contribution in explaining species richness is given by the elevational range, which, however, exerts a negative influence. Elevational range is usually considered as a measure of environmental heterogeneity, because it is correlated with variation in temperature, precipitation, humidity, wind speed, evaporation, and insolation, and altitude is often found to be an important variable in explaining species numbers on islands, in some cases ranking only second to, or even ahead of, island area [73]. On the contrary, in the present research, elevational range had a negative influence, albeit only in the case of Hydradephaga and possibly Carabidae. This may be interpreted as a consequence of the fact that, in a mainland context, a large elevational gradient implies that most of the study area is at high altitudes. Since, in general, richness tends to decrease with elevation [94,95,96], the negative influence of elevational range may be a reflection of this general trend. Moreover, in the specific case of the Hydradephaga, the negative impact of the elevational range on species richness may be explained by the fact that, with increasing elevation, lentic waters became rare. Elevation exerted a negative influence also on Tenebrionidae, but, in this case, the variable retained in the multiple model was minimum elevation. The negative impact of minimum elevation on species richness can be explained by the fact that many tenebrionids are associated with coastal areas [97].
In accordance with the general impoverishment of species richness with increasing latitude observed for many taxa in the northern hemisphere [79,80,81,82,83], latitude had a negative influence on Tenebrionidae and coprophagous Scarabaeoidea. This can be explained with reference to both current climatic factors and biogeographical history. Tenebrionidae are mainly thermophilic animals, including many groups associated with hot and arid environments [27,97], and their species richness in Europe tends to decrease northwards due to the joint effect of current climatic factors and post glaciation incomplete recolonization [98,99,100]. In accordance with the general latitudinal pattern of biodiversity in Europe, tenebrionid richness in Italy is known to increase southwards [16], in response to variations in rainfall and temperatures, and in consequence of the refugial role played by Southern Italy during the Pleistocene [101]. There is no previous study on the latitudinal gradient of dung beetles in Europe or in Italy, but studies on dung beetle communities in Europe and North America [102,103,104] indicate that northern communities have fewer species than the southern ones. The increase in dung beetle richness with decreasing latitude in Italy can be explained by both historical reasons (i.e., the refugial role of southern areas during Pleistocene glaciations) and current climate (for example, while Aphodiini are able to colonize even cold areas, most Scarabaeinae are thermophilic animals) and land use (extensive grazing in southern Italian regions).
Extinction rates as predicted by the SAR are highest for dung beetles, which is consistent with their dependence on dung, which, in turn, is mainly associated with the presence of large grazing areas. Actually, dung beetles were the insect group which experienced the highest documented extinction rate in the city of Rome as a consequence of the loss of grazing lands due to urbanization [105]. The second most sensitive group was the Tenebrionidae, probably due to their low dispersal capabilities, which reduce their possibility of colonizing new areas when environmental conditions become less favorable. A previous study focused on these insects revealed that the geophilous tenebrionids, which are typically flightless, have higher extinction rates than the xylophilous ones [16]. Moreover, Tenebrionidae include many saproxylic beetles, which are particularly sensitive to the loss of forest vegetation [106,107]. The comparatively low extinction rates predicted for Carabidae, phytophagous Scarabaeoidea and Hydradephaga can be explained by their higher dispersal power, which reduces habitat confinement and hence makes them less sensitive to area loss, despite their dependence on other animals as food (Carabidae), plants (phytophagous Scarabaeoidea), or specific biotopes (lentic water for Hydradephaga). If other variables besides area are considered in multiple models, however, the extinction rates expected by area loss became more similar among groups, suggesting that differences in extinction rates are largely due to the influence of other factors than area. Including the effect of latitude in multiple models reduces the expected extinction rates for Tenebrionidae and coprophagous Scarabaeoidea, which highlights a prominent role of the southern areas for their conservation in Italy. In other words, for these beetles, area loss is expected to act more dramatically in southern areas. By contrast, for Carabidae and Hydradephaga, the inclusion of elevational range increased the extinction risk, which suggests that lowland areas are those that most contribute to the conservation of these beetles. Finally, the extinction rates expected by area loss are always relatively low. Thus, in small areas hosting relatively few species, a very small number of species will be lost, even if they represent a large proportion of the considered fauna. This means that, because of difficulties in obtaining complete species lists and in monitoring all species, in small reserves, with few species, the number of possible extinctions might be too small to be detected, and extinctions might remain unnoticed.
5. Conclusions
Reserve area was an important predictor of species richness in all investigated beetle groups, which is consistent with the almost ubiquitous pattern known as the species–area relationship (SAR). For Carabidae, Hydradephaga, and Tenebrionidae, elevation exerted a negative influence—a possible reflection of the general decline in species richness with altitude commonly observed in many taxa. In accordance with the general impoverishment of species richness northwards, latitude had a negative influence on coprophagous Scarabaeoidea and Tenebrionidae, as a result of current and past biogeographical factors (that is, more favorable climatic conditions in southern areas and their role as Pleistocene refuges). SAR-based extinction rates are highest for dung beetles, consistently with their dependence on large grazing areas, and for Tenebrionidae, because of their low dispersal capabilities. The relatively lower extinction rates predicted for Carabidae, phytophagous Scarabaeoidea, and Hydradephaga can be explained by their higher dispersal power. Thus, both ecological needs and dispersal abilities emerge as important determinants of SAR-based extinction rates. If other variables besides area are considered, extinction rates become more similar among groups. Because of the overall low extinction rates, extinctions might be too few to be easily detected in reserves with few species. This should be carefully considered when comparing SAR-based extinction rates with empirical data.
Supplementary Materials
The following are available online at https://www.mdpi.com/2075-4450/11/9/646/s1, Figure S1: number of extinct species expected on the basis of area loss (% of area lost) for various groups of beetles in Italian reserves for areas hosting 10, 20, 40, 60, 80, and 100 species. Curves were calculated using z-values obtained from the power function of the species–area relationship and from multiple models. Note the log-scale for the y-axis. (a) and (b) Carabidae; (c) and (d) Hydradephaga; (e) and (f) coprophagous Scarabaeoidea; (g) phytophagous Scarabaeoidea; (h) total Scarabaeoidea; (i) and (j) Tenebrionidae. Panels (a), (c), (e), (g), (h), and (i) refer to z-values obtained from the power function. Panels (b), (d), (f), and (j) refer to z-values obtained from multiple models.
Funding
This research was partially funded by the University of L’Aquila, Department of Life, Health and Environmental Sciences, FFO 2020, “Analisi multi-taxa della struttura di comunità animali, vegetali e microbiche in alta quota”.
Acknowledgments
Thanks are due to C. Mantoni for her assistance in preparing Figure 1, to L. Di Biase for her assistance with references, and to three anonymous reviewers for their comments on a previous version of this paper.
Conflicts of Interest
The author declares no conflict of interest. The funder had no role in the design of the study; in the collection, analyses, or interpretation of data; in the writing of the manuscript, or in the decision to publish the results.
References
- Matthews, T.; Triantis, K.; Whittaker, R. (Eds.) The Species-Area Relationship: Theory and Application (Ecology, Biodiversity and Conservation); Cambridge University Press: Cambridge, UK, 2020; pp. 1–481. [Google Scholar]
- Fattorini, S. Detecting biodiversity hotspots by species-area relationships: A case study of Mediterranean beetles. Conserv. Biol. 2006, 20, 1169–1180. [Google Scholar] [CrossRef] [PubMed]
- Mazel, F.; Guilhaumon, F.; Mouquet, N.; Dvictor, V.; Gravel, D.; Renaud, J.; Cianciaruso, M.V.; Loyola, R.; Diniz-Filho, J.A.F.; Mouillot, D.; et al. Global hotspots of multifaceted mammal diversity. Glob. Ecol. Biogeogr. 2014, 23, 836–847. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Fattorini, S. The Identification of Biodiversity Hotspots Using the Species–Area Relationship. In The Species-Area Relationship: Theory and Application (Ecology, Biodiversity and Conservation); Matthews, T., Triantis, K., Whittaker, R., Eds.; Cambridge University Press: Cambridge, UK, 2020; pp. 321–344. [Google Scholar]
- Desmet, P.; Cowling, R. Using the species–area relationship to set baseline targets for conservation. Ecol. Soc. 2004, 9, 11. [Google Scholar] [CrossRef]
- Fattorini, S. The use of cumulative area curves in biological conservation: A cautionary note. Acta Oecol. 2010, 36, 255–258. [Google Scholar] [CrossRef]
- Fattorini, S. Urban biodiversity hotspots are not related to the structure of green spaces: A case study of tenebrionid beetles from Rome, Italy. Urban Ecosyst. 2014, 17, 1033–1045. [Google Scholar] [CrossRef]
- Cox, R.L.; Underwood, E.C. The importance of conserving biodiversity outside of protected areas in Mediterranean ecosystems. PLoS ONE 2011, 6, e14508. [Google Scholar] [CrossRef] [PubMed]
- Fattorini, S.; Borges, P.V.A. Species–area relationships underestimate extinction rates. Acta Oecol. 2012, 40, 27–30. [Google Scholar] [CrossRef] [Green Version]
- Fattorini, S.; Matthews, T.; Ulrich, W. Using the Species–Area Relationship to Predict Extinctions Resulting from Habitat Loss. In The Species-Area Relationship: Theory and Application (Ecology, Biodiversity and Conservation); Matthews, T., Triantis, K., Whittaker, R., Eds.; Cambridge University Press: Cambridge, UK, 2020; pp. 345–367. [Google Scholar]
- Triantis, K.A.; Guilhaumon, F.; Whittaker, R.J. The island species–area relationship: Biology and statistics. J. Biogeogr. 2012, 39, 215–231. [Google Scholar] [CrossRef]
- Matthews, T.J.; Guilhaumon, F.; Triantis, K.A.; Borregaard, M.K.; Whittaker, R.J. On the form of species–area relationships in habitat islands and true islands. Glob. Ecol. Biogeogr. 2016, 25, 847–858. [Google Scholar] [CrossRef]
- Fattorini, S.; Borges, P.V.A.; Dapporto, L.; Strona, G. What can the parameters of the species–area relationship (SAR) tell us? Insights from Mediterranean islands. J. Biogeogr. 2017, 44, 1018–1028. [Google Scholar] [CrossRef]
- Báldi, A.; Vörös, J. Extinction debt of Hungarian reserves: A historical perspective. Basic Appl. Ecol. 2006, 7, 289–295. [Google Scholar] [CrossRef]
- Benedick, S.; Hill, J.K.; Mustaffa, N.; Chey, V.K.; Maryati, M.; Searle, J.B.; Schilthuizen, M.; Hamer, K.C. Impacts of rain forest fragmentation on butterflies in northern Borneo: Species richness, turnover and the value of small fragments. J. Appl. Ecol. 2006, 43, 967–977. [Google Scholar] [CrossRef]
- Fattorini, S. Conservation Biogeography of Tenebrionid Beetles: Insights from Italian Reserves. Diversity 2020, 12, 348. [Google Scholar] [CrossRef]
- Médail, F.; Quézel, P. Biodiversity Hotspots in the Mediterranean Basin: Setting Global Conservation Priorities. Conserv. Biol. 1999, 13, 1510–1513. [Google Scholar] [CrossRef]
- Myers, N.; Mittermeier, R.A.; Mittermeier, C.G.; da Fonseca, G.A.B.; Kent, J. Biodiversity hotspots for conservation priorities. Nature 2000, 403, 853–858. [Google Scholar] [CrossRef]
- Ruffo, S.; Stoch, F. (Eds.) Checklist and Distribution of the Italian Fauna. 10,000 Terrestrial and Inland Water Species. 2 Serie, Sez. Scienze Della Vita; Memorie del Museo Civico di Storia Naturale di Verona: Verona, Italy, 2006; Volume 17, pp. 1–303. [Google Scholar]
- Blasi, C.; Biondi, E. La flora in Italia; Ministero dell’Ambiente e della Tutela del Territorio e del Mare, Sapienza Università Editrice: Rome, Italy, 2017; pp. 1–704. [Google Scholar]
- Blondel, J.; Aronson, J. Biology and Wildlife of the Mediterranean Region; Oxford University Press: Oxford, UK, 1999; pp. 1–328. [Google Scholar]
- Underwood, E.C.; Viers, J.H.; Klausmeyer, K.R.; Cox, R.L.; Shaw, M.R. Threats and biodiversity in the mediterranean biome. Divers. Distrib. 2009, 15, 188–197. [Google Scholar] [CrossRef]
- Ministero Dell’ambiente e Della Tutela del Territorio e del Mare. VI Elenco Ufficiale delle Aree Protette (EUAP). Supplemento Ordinario alla “Gazzetta Ufficiale” No. 125 31/05/2010–Serie Generale; Ministero dell’Ambiente e della Tutela del Territorio e del Mare: Roma, Italy, 2010. [Google Scholar]
- Blasi, C.; Boitani, L.; La Posta, S.; Manes, F.; Marchetti, M. Stato della Biodiversità in Italia; Palombi Editori: Roma, Italy, 2005; pp. 1–468. [Google Scholar]
- Hunt, T.; Bergsten, J.; Levkanicova, Z.; Papadopoulou, A.; St. John, O.; Wild, R.; Hammond, P.M.; Ahrens, D.; Balke, M.; Caterino, M.S.; et al. A comprehensive phylogeny of beetles reveals the evolutionary origins of a superradiation. Science 2007, 318, 1913–1916. [Google Scholar] [CrossRef]
- Powell, J.; Coleoptera, A. Encyclopedia of Insects, 2nd ed.; Resh, H.V., Cardé, T.R., Eds.; Academic Press: New York, NY, USA, 2009; p. 199. [Google Scholar]
- Dajoz, R. Les Coléoptères Carabides et Ténébrionidés; Lavoisier: Paris, France, 2002; pp. 1–522. [Google Scholar]
- Rocchi, S. Insecta Coleoptera Hydradephaga. In Checklist and Distribution of the Italian Fauna. 10,000 Terrestrial and Inland Water Species. 2 serie, Sez. Scienze Della Vita; Ruffo, S., Stoch, F., Eds.; Memorie del Museo Civico di Storia Naturale di Verona: Verona, Italy, 2006; Volume 17, pp. 165–166. [Google Scholar]
- Baca, S.M.; Alexander, A.; Gustafson, G.T.; Short, A.E.Z. Ultraconserved elements show utility in phylogenetic inference of Adephaga (Coleoptera) and suggest paraphyly of ‘Hydradephaga’. Syst. Entomol. 2017, 42, 786–795. [Google Scholar] [CrossRef]
- Carpaneto, G.M.; Piattella, E.; Valerio, L. Insecta Coleoptera Scarabaeoidea. In Checklist and Distribution of the Italian Fauna. 10,000 Terrestrial and Inland Water Species. 2 serie, Sez. Scienze Della Vita; Ruffo, S., Stoch, F., Eds.; Memorie del Museo Civico di Storia Naturale di Verona: Verona, Italy, 2006; Volume 17, pp. 193–196. [Google Scholar]
- Fattorini, S.; Mantoni, C.; Audisio, P.; Biondi, M. Taxonomic variation in levels of endemism: A case study of Italian tenebrionid beetles. Insect Conserv. Divers. 2019, 12, 351–361. [Google Scholar] [CrossRef]
- Angelini, F. Contribution to the knowledge of beetles (Insecta Coleoptera) of some protected areas of Apulia, Basilicata and Calabria (Italy). Biodivers. J. 2020, 11, 85–254. [Google Scholar] [CrossRef]
- Crucitti, P.; Brocchieri, D.; Bubbico, F.; Castelluccio, P.; Cervoni, F.; Di Russo, E.; Emiliani, F.; Giardini, M.; Pulvirenti, E. Checklist di alcuni gruppi selezionati dell’entomofauna del Parco Naturale Archeologico dell’Inviolata (Guidonia Montecelio, Roma): XlI contributo allo studio della biodiversità della Campagna Romana a nord-est di Roma. Boll. Soc. Ent. Ital. 2019, 151, 65–92. [Google Scholar] [CrossRef]
- Fattorini, S.; Vigna Taglianti, A. Use of taxonomic and chorological diversity to highlight the conservation value of insect communities in a Mediterranean coastal area: The carabid beetles (Coleoptera, Carabidae) of Castelporziano (Central Italy). Rend. Lincei Sci. Fis. 2015, 26, 625–641. [Google Scholar] [CrossRef]
- Furlanetto, D. Atlante della Biodiversità nel Parco Ticino—Edizione 2002. Elenchi Sistematici (Monografie); Consorzio Parco Lombardo della Valle del Ticino: Milan, Italy, 2002; p. 406. [Google Scholar]
- Lestes–Associazione di Ricerca e Studio nelle Scienze Naturali (2004) Relazione Finale della Ricerca Sui Coleotteri Carabidi del Parco Nazionale delle Foreste Casentinesi, Monte Falterona e Campigna. Available online: https://www.parcoforestecasentinesi.it/sites/default/files/images/cartella_ricerca/carabidi.pdf (accessed on 20 June 2020).
- Lucarelli, E.; Chelazzi, L.; Colombini, I.; Fallaci, M.; Mascagni, A. La coleotterofauna del tombolo antistante la laguna di Burano (GR): Lista e zonazioni delle specie raccolte durante un intero anno di campionamenti. Boll. Assoc. Romana Entomol. 1993, 47, 7–34. [Google Scholar]
- Vigna Taglianti, A. Coleotteri Carabidi del Parco Nazionale d’Abruzzo (Coleoptera Adephaga, Carabidae). In Progetto Biodiversità, Liste Preliminari Degli Organismi viventi del Parco Nazionale d’Abruzzo; Tassi, F., Ed.; Ente Autonomo Parco Nazionale d’Abruzzo: Roma, Italy, 1995; Volume 1, p. 32. [Google Scholar]
- Vigna Taglianti, A. Coleoptera Carabidae (Insecta). In Ricerche sulla Valle Peligna (Italia Centrale, Abruzzo). Quaderni di Provincia oggi, 23/II; Osella, B.G., Biondi, M., Di Marco, C., Riti, M., Eds.; Amministrazione Provinciale de L’Aquila: L’Aquila, Italy, 1997; Volume 2, pp. 301–334. [Google Scholar]
- Vigna Taglianti, A. Carabidae (Coleoptera). In Invertebrati di una foresta della Pianura Padana, Bosco della Fontana, Secondo Contributo, Conservazione Habitat Invertebrati; Cerretti, P., Hardersen, S., Mason, F., Nardi, G., Tisato, M., Zapparoli, M., Eds.; Cierre Grafica Editore: Verona, Italy, 2004; Volume 3, pp. 51–63. [Google Scholar]
- Vigna Taglianti, A. I Coleotteri Carabidi (Coleoptera: Carabidae). In Artropodi del Parco Nazionale del Vesuvio: Ricerche preliminari. Conservazione Habitat Invertebrati; Nardi, G., Vomero, V., Eds.; Cierre edizioni: Verona, Italy, 2007; Volume 4, pp. 99–112. [Google Scholar]
- Vigna Taglianti, A. I Carabidi delle Riserve Naturali “Agoraie di Sopra e Moggetto” (Liguria, Genova) e “Guadine Pradaccio” (Emilia–Romagna, Parma) (Coleoptera, Carabidae). Boll. Assoc. Romana Entomol. 2009, 64, 99–127. [Google Scholar]
- Vigna Taglianti, A.; Audisio, P.; De Felici, S. I Coleotteri Carabidi del Parco Nazionale della Val Grande (Verbania, Piemonte). Rivista Piemontese di Storia Naturale 1999, 19, 193–245. [Google Scholar]
- Vigna Taglianti, A.; Bonavita, P.; Di Giulio, A.; Todini, A.; Maltzeff, P. I Carabidi della Tenuta Presidenziale di Castelporziano (Coleoptera, Carabidae). Boll. Assoc. Romana Entomol. 2001, 56, 115–173. [Google Scholar]
- Vigna Taglianti, A.; Bonavita, P.; Maltzeff, P. I Carabidi della Tenuta Presidenziale di Castelporziano. Addenda (Coleoptera, Carabidae). Boll. Assoc. Romana Entomol. 2007, 62, 101–103. [Google Scholar]
- Nardi, G. Adephagous water beetles: Faunistics, ecology and conservation (Coleoptera, Gyrinidae, Haliplidae, Noteridae, Dytiscidae). In Invertebrati di una Foresta della Pianura Padana, Bosco della Fontana, Secondo Contributo, Conservazione Habitat Invertebrati; Cerretti, P., Hardersen, S., Mason, F., Nardi, G., Tisato, M., Zapparoli, M., Eds.; Cierre Grafica Editore: Verona, Italy, 2004; Volume 3, pp. 65–115. [Google Scholar]
- Nardi, G. Dati preliminari sui coleotteri idroadefagi del Parco Nazionale del Circeo. In Habitat, Flora e Fauna del Parco Nazionale del Circeo; Zerunian, S., Ed.; Uff. Gestione Beni ex ASFD di Sabaudia–Parco Nazionale del Circeo: Latina, Italy, 2005; pp. 151–178. [Google Scholar]
- Nardi, G.; Maltzeff, P. Gli Idroadefagi della Tenuta Presidenziale di Castelporziano (Coleoptera, Gyrinidae, Haliplidae, Noiteridae, Hygrobiidae, Dytiscidae). Boll. Assoc. Romana Entomol. 2001, 56, 175–232. [Google Scholar]
- Nardi, G.; Cornacchia, P.; Canepari, C.; Colonnelli, E.; Liberti, G.; Mascagni, A.; Piattella, E.; Rocchi, S.; Zoia, S. Note sui Coleotteri delle Riserve Naturali “Agoraie di Sopra e Moggetto” (Liguria, Genova) e “Guadine Pradaccio” (Emilia-Romagna, Parma). (Coleoptera). Boll. Assoc. Romana Entomol. 2009, 64, 183–280. [Google Scholar]
- Rocchi, S.; Mascagni, A. Coleotterofauna acquatica e semiacquatica del Parco Nazionale delle Foreste Casentinesi, Monte Falterona e Campigna (Insecta Coleoptera Hydradephaga, Hydraenidae, Hydrophiloidea, Dryopoidea). Quaderno di Studi e Notizie di Storia Naturale della Romagna Cesena 2005, 21, 1–32. [Google Scholar]
- Carpaneto, G.M.; Migliaccio, E.; Piattella, E. Coleotteri Lamellicorni del Parco Nazionale d’Abruzzo. In Progetto Biodiversità, Liste Preliminari Degli Organismi Viventi del Parco Nazionale d’Abruzzo; Tassi, F., Ed.; Ente Autonomo Parco Nazionale d’Abruzzo: Roma, Italy, 1996; Volume 1, p. 29. [Google Scholar]
- Carpaneto, G.M.; Maltzeff, P.; Piattella, E.; Pontuale, G. I Coleotteri lamellicorni della tenuta presidenziale di Casteporziano e delle aree limitrofe (Coleoptera, Lamellicornia). Boll. Assoc. Romana Entomol. 1998, 52, 9–54. [Google Scholar]
- Carpaneto, G.M.; Maltzeff, P.; Piattella, E.; Facchinelli, L. Nuovi reperti di Coleotteri Lamellicorni della Tenuta Presidenziale di Castelporziano e delle aree limitrofe (Coleoptera, Lamellicornia). Boll. Assoc. Romana Entomol. 2001, 56, 311–329. [Google Scholar]
- Hardersen, S.; Toni, I.; Cornacchia, P.; Curletti, G.; Leo, P.; Nardi, G.; Penati, F.; Piattella, E.; Platia, G. Survey of selected beetle families in a floodplain remnant in northern Italy. Bull. Insectol. 2012, 65, 199–207. [Google Scholar]
- Lapiana, F.; Sparacio, I. I Coleotteri Lamellicorni delle Madonie (Sicilia) (Insecta Coleoptera Lucanoidea et Scarabaeoidea). Nat. Sicil. 2006, 30, 227–292. [Google Scholar]
- Marotta, O.; Andreetti, A.; Osella, G. Coleoptera Lucanoidea & Scarabaeoidea. In Ricerche sulla Valle Peligna (Italia Centrale, Abruzzo). Quaderni di Provincia Oggi, 23/II; Osella, B.G., Biondi, M., Di Marco, C., Riti, M., Eds.; Amministrazione Provinciale de L’Aquila: L’Aquila, Italy, 1997; pp. 363–377. [Google Scholar]
- Piattella, E. Coleoptera, Scarabaeoidea (Aphodiidae, Cetoniidae, Melolonthidae, Scarabaeidae). In Invertebrati di una Foresta della Pianura Padana, Bosco della Fontana, Secondo Contributo, Conservazione Habitat Invertebrati; Cerretti, P., Hardersen, S., Mason, F., Nardi, G., Tisato, M., Zapparoli, M., Eds.; Cierre Grafica Editore: Verona, Italy, 2004; Volume 3, pp. 278–280. [Google Scholar]
- Piattella, E. I Coleotteri Lamellicorni del Parco e considerazioni sul popolamento dell’area partenopea (Coleoptera: Lucanidae, Aphodiidae, Orphnidae, Scarabaeidae, Melolonthidae, Rutelidae e Cetoniidae). In Artropodi del Parco Nazionale del Vesuvio: Ricerche preliminari. Conservazione Habitat Invertebrati; Nardi, G., Vomero, V., Eds.; Cierre Edizioni: Verona, Italy, 2007; Volume 4, pp. 147–163. [Google Scholar]
- Piattella, E. Contribution to the knowledge of scarab beetles of Vincheto di Celarda Nature Reserve (Coleoptera: Scarabaeoidea). In Research on the Natural Heritage of the Reserves Vincheto di Celarda and Val Tovanella (Belluno Province, Italy). Conservation of Two Protected Areas in the Context of a Life Project. Quaderni Conservazione Habitat Invertebrati; Handersen, S., Mason, F., Viola, F., Campedel, D., Lasen, C., Cassol, M., Eds.; Arti grafiche Fiorini: Verona, Italy, 2009; Volume 5, pp. 117–124. [Google Scholar]
- Aliquò, V.; Leo, P. I Coleotteri Tenebrionidi delle Madonie (Sicilia). Nat. Sicil. 1996, 4, 281–304. [Google Scholar]
- Andreetti, A.; Di Gaetano, B.; Di Marco, C.; Osella, G.; Riti, M. Coleoptera Tenebrionidae (Insecta). In Ricerche sulla Valle Peligna (Italia Centrale, Abruzzo). Quaderni di Provincia oggi, 23/II; Osella, B.G., Biondi, M., Di Marco, C., Riti, M., Eds.; Amministrazione provinciale de L’Aquila: L’Aquila, Italy, 1997; Volume 2, pp. 425–443. [Google Scholar]
- Cavanna, C. Parte II. Catalogo dogli animali raccolti al Vulture, al Pollino ed in altri luoghi dell’Italia meridionale e centrale. Boll. Soc. Ent. Ital. 1882, 14, 31–87. [Google Scholar]
- Chelazzi, L.; DeMatthaeis, E.; Colombini, I.; Fallaci, M.; Bandini, V.; Tozzi, C. Abundance, zonation and ecological indices of a coleopteran community from a sandy beach-dune ecosystem of the southern Adriatic coast, Italy. Vie Milieu 2005, 55, 127–141. [Google Scholar]
- Fattorini, S. The Tenebrionidae (Coleoptera) of a Tyrrhenian coastal area: Diversity and zoogeographical composition. Biogeographia 2002, 23, 103–126. [Google Scholar] [CrossRef]
- Fattorini, S. I Coleotteri Tenebrionidi del Parco Nazionale del Circeo (Italia Centrale) (Coleoptera, Tenebrionidae). Boll. Assoc. Romana Entomol. 2005, 60, 47–104. [Google Scholar]
- Fattorini, S. The tenebrionid beetles of Mt Vesuvius: Species assemblages and biogeographic kinetics on an active volcano (Coleoptera Tenebrionidae). In Artropodi del Parco Nazionale del Vesuvio: Ricerche Preliminari. Conservazione Habitat Invertebrati; Nardi, G., Vomero, V., Eds.; Cierre Edizioni: Verona, Italy, 2007; Volume 4, pp. 221–243. [Google Scholar]
- Fattorini, S.; Maltzeff, P.; Salvati, L. Use of insect distribution across landscape-soil units to assess conservation priorities in a Mediterranean coastal reserve: The tenebrionid beetles of Castelporziano (Central Italy). Rend. Lincei-Sci. Fis. 2015, 26, 353–366. [Google Scholar] [CrossRef]
- Fattorini, S.; Romiti, F.; Carpaneto, G.M.; Poeta, G.; Bergamaschi, D. I Coleotteri Tenebrionidi del Sito d’Importanza Comunitaria “Foce Saccione—Bonifica Ramitelli” (Molise) (Coleoptera Tenebrionidae). Boll. Soc. Ent. Ital. 2016, 148, 57–62. [Google Scholar] [CrossRef]
- Gatti, E. Ricerche sull’entomofauna della Riserva Naturale Vincheto di Celarda–(BL) Collana Verde, 86; Ministero dell’Agricoltura e delle Foreste: Rome, Italy, 1991; p. 200.
- Scupola, A. Tenebrionidae. In Invertebrati di una Foresta della Pianura Padana. Bosco della Fontana. Centro Nazionale per lo Studio e la Conservazione della Biodiversià Forestale Bosco della Fontana; Mason, F., Cerretti, P., Tagliapietra, A., Speight, M.C.D., Zapparoli, M., Eds.; Gianluigi Arcari Editore: Mantova, Italy, 2002; p. 93. [Google Scholar]
- Scupola, A. Coleoptera, Tenebrionidae (excluding Alleculinae, Lagriinae). In Invertebrati di una Foresta della Pianura Padana, Bosco della Fontana, Secondo Contributo, Conservazione Habitat Invertebrati; Cerretti, P., Hardersen, S., Mason, F., Nardi, G., Tisato, M., Zapparoli, M., Eds.; Cierre Grafica Editore: Verona, Italy, 2004; Volume 3, p. 285. [Google Scholar]
- Connor, E.F.; McCoy, E.D. The statistics and biology of the species–area relationship. Am. Nat. 1979, 113, 791–833. [Google Scholar] [CrossRef]
- Fattorini, S.; Dapporto, L.; Strona, G.; Borges, P.V.A. Calling for a new strategy to measure environmental (habitat) diversity in Island Biogeography: A case study of Mediterranean tenebrionids (Coleoptera: Tenebrionidae). Fragm. Entomol. 2015, 47, 1–14. [Google Scholar] [CrossRef]
- Spada, L.; Bardiani, M.; Nardi, G. Artropodi delle Riserve Naturali “Agoraie di Sopra e Moggetto” (Liguria, Genova) e “Guadine Pradaccio” (Emilia-Romagna, Parma). Introduzione. Boll. Assoc. Romana Entomol. 2009, 64, 3–32. [Google Scholar]
- Angelini, F. Coleotterofauna del Promontorio del Gargano (Coleoptera). Atti Mus. Civ. Stor. Nat. Grosseto 1987, 11/12, 5–84. [Google Scholar]
- Nardi, G.; Vomero, V. Introduzione. In Artropodi del Parco Nazionale del Vesuvio: Ricerche preliminari. Conservazione Habitat Invertebrati; Nardi, G., Vomero, V., Eds.; Cierre edizioni: Verona, Italy, 2007; pp. 15–28. [Google Scholar]
- Angelini, F. Coleotterofauna del Massiccio del Pollino (Basilicata-Calabria) (Coleoptera). Entomologica 1986, 21, 37–125. [Google Scholar]
- Angelini, F. Coleotterofauna dell’Altipiano della Sila (Calabria, Italia) (Coleoptera). Mem. Soc. Entomol. Ital. 1991, 70, 171–254. [Google Scholar]
- Ashton, K.G. Are ecological and evolutionary rules being dismissed prematurely? Divers. Distrib. 2001, 7, 289–295. [Google Scholar] [CrossRef] [Green Version]
- Fattorini, S. Testing the latitudinal gradient: A narrow-scale analysis of tenebrionid richness (Coleoptera, Tenebrionidae) in the Aegean archipelago (Greece). Ital. J. Zool. 2006, 73, 203–211. [Google Scholar] [CrossRef]
- Whittaker, R.J.; Nogues-Bravo, D.; Araujo, M.B. Geographical gradients of species richness: A test of the water energy conjecture of Hawkins et al. (2003) using European data for five taxa. Global Ecol. Biogeogr. 2007, 16, 76–89. [Google Scholar] [CrossRef]
- Baselga, A. Determinants of species richness, endemism and turnover in European longhorn beetles. Ecography 2008, 31, 263–271. [Google Scholar] [CrossRef]
- Schuldt, A.; Assmann, T. Environmental and historical effects on richness and endemism patterns of carabid beetles in the western Palaearctic. Ecography 2009, 32, 705–714. [Google Scholar] [CrossRef]
- Burnham, K.P.; Anderson, D.R. Model. Selection and Multimodel Inference: A Practical Information-Theoretic Approach, 2nd ed.; Springer: New York, NY, USA, 2002; p. 488. [Google Scholar]
- R Core Team. R: A Language and Environment for Statistical Computing; R Foundation for Statistical Computing. 2018. Available online: http://www.r-project.org/ (accessed on 10 September 2018).
- Bartoń, K. MuMIn: Multi-Model Inference. R Package Version 1.43.17. Available online: https://CRAN.R-project.org/package=MuMIn2020 (accessed on 10 June 2020).
- Den Boer, P.J. On the significance of dispersal power for populations of carabid-beetles (Coleoptera, Carabidae). Oecologia 1970, 4, 1–28. [Google Scholar] [CrossRef] [PubMed]
- Thiele, H.U. Dispersal and Dispersal Power of Carabid Beetles. In Carabid Beetles in Their Environments. Zoophysiology and Ecology; Thiele, H.U., Ed.; Springer: Berlin/Heidelberg, Germany, 1977; Volume 10, pp. 284–297. [Google Scholar]
- Den Boer, P.J. Density limits and survival of local populations in 64 carabid species with different powers of dispersal. J. Evol Biol. 1990, 3, 19–48. [Google Scholar] [CrossRef] [Green Version]
- Bilton, D.T. Dispersal in Dytiscidae. In Ecology, Systematics, and Natural History of Predaceous Diving Beetles (Coleoptera: Dytiscidae); Yee, D.A., Ed.; Springer: New York, NY, USA, 2014; pp. 387–407. [Google Scholar]
- Fattorini, S. Both Recent and Pleistocene geography determine animal distributional patterns in the Tuscan Archipelago. J. Zool. 2009, 277, 291–301. [Google Scholar] [CrossRef]
- Fattorini, S. Faunal patterns in tenebrionids (Coleoptera: Tenebrionidae) on the Tuscan Islands: The dominance of paleogeography over Recent geography. Eur. J. Entomol. 2009, 106, 415–423. [Google Scholar] [CrossRef] [Green Version]
- Papadopoulou, A.; Anastasiou, I.; Keskin, B.; Vogler, A.P. Comparative phylogeography of tenebrionid beetles in the Aegean archipelago: The effect of dispersal ability and habitat preference. Mol. Ecol. 2009, 18, 2503–2517. [Google Scholar] [CrossRef]
- Fattorini, S. Disentangling the effects of available area, mid-domain constraints, and species environmental tolerance on the altitudinal distribution of tenebrionid beetles in a Mediterranean area. Biodivers. Conserv. 2014, 23, 2545–2560. [Google Scholar] [CrossRef]
- Fattorini, S.; Mantoni, C.; Di Biase, L.; Strona, G.; Pace, L.; Biondi, M. Elevational Patterns of Generic Diversity in the Tenebrionid Beetles (Coleoptera Tenebrionidae) of Latium (Central Italy). Diversity 2020, 12, 47. [Google Scholar] [CrossRef] [Green Version]
- Fattorini, S.; Mantoni, C.; Di Biase, L.; Pace, L. Mountain Biodiversity and Sustainable Development. In Encyclopedia of the UN Sustainable Development Goals. Life on Land; Leal Filho, W., Azul, A., Brandli, L., Özuyar, P., Wall, T., Eds.; Springer International Publishing: New York, NY, USA, 2020; pp. 1–31. [Google Scholar]
- Fattorini, S. Ecology and conservation of tenebrionid beetles in Mediterranean coastal areas. In Insect Ecology and Conservation; Fattorini, S., Ed.; Research Signpost: Trivandrum, Kerala, India, 2008; pp. 165–297. [Google Scholar]
- Fattorini, S.; Baselga, A. Species richness and turnover patterns in European tenebrionid beetles. Insect Conserv. Diver. 2012, 5, 331–345. [Google Scholar] [CrossRef]
- Fattorini, S.; Ulrich, W. Drivers of species richness in European Tenebrionidae (Coleoptera). Acta Oecol. 2012, 43, 22–28. [Google Scholar] [CrossRef]
- Fattorini, S.; Ulrich, W. Spatial distributions of European Tenebrionidae point to multiple postglacial colonization trajectories. Biol. J. Linn. Soc. 2012, 105, 318–329. [Google Scholar] [CrossRef]
- Fattorini, S. Tenebrionid beetle distributional patterns in Italy: Multiple colonisation trajectories in a biogeographical crossroad. Insect Conserv. Divers. 2014, 7, 144–160. [Google Scholar] [CrossRef]
- Hanski, I. Species Richness. In Dung Beetle Ecology; Hanski, I., Cambefort, Y., Eds.; Princeton University Press: Princeton, NJ, USA, 1991; pp. 350–365. [Google Scholar]
- Lobo, J.M. Species diversity and composition of dung beetle (Coleoptera: Scarabaeoidea) assemblages in North America. Can. Entomol. 2000, 132, 307–321. [Google Scholar] [CrossRef]
- Radtke, M.G.; da Fonseca, C.R.V.; Williamson, G.B. Dung beetle communities: A neotropical-north temperate comparison. Neotrop. Entomol. 2010, 39, 19–27. [Google Scholar] [CrossRef] [Green Version]
- Fattorini, S. Insect extinction by urbanization: A long term study in Rome. Biol. Conserv. 2011, 144, 370–375. [Google Scholar] [CrossRef]
- Carpaneto, G.M.; Baviera, C.; Biscaccianti, A.B.; Brandmayr, P.; Mazzei, A.; Mason, F.; Battistoni, A.; Teofili, C.; Rondinini, C.; Fattorini, S.; et al. Red List of Italian Saproxylic Beetles: Taxonomic overview, ecological features and conservation issues (Coleoptera). Fragm. Entomol. 2015, 47, 53–126. [Google Scholar] [CrossRef]
- Fattorini, S.; Galassi, D.M.P. Role of urban green spaces for saproxylic beetle conservation: A case study of tenebrionids in Rome, Italy. J. Insect Conserv. 2016, 20, 737–745. [Google Scholar] [CrossRef]
Figure 1.
Location of the 23 Italian reserves considered in this study. Reserve names are given in Table 1.
Figure 1.
Location of the 23 Italian reserves considered in this study. Reserve names are given in Table 1.

Figure 2.
Species–area relationships (SARs) for various beetle groups in Italian reserves: (a) Carabidae, (b) Hydradephaga, (c) coprophagous Scarabaeoidea, (d) phytophagous Scarabaeoidea, (e) total Scarabaeoidea, (f) Tenebrionidae. Decimal logarithms are used.
Figure 2.
Species–area relationships (SARs) for various beetle groups in Italian reserves: (a) Carabidae, (b) Hydradephaga, (c) coprophagous Scarabaeoidea, (d) phytophagous Scarabaeoidea, (e) total Scarabaeoidea, (f) Tenebrionidae. Decimal logarithms are used.

Figure 3.
Extinction rates (% of extinct species) expected on the basis of area loss (% of area lost) for various beetle groups in Italian reserves: (a) Carabidae, (b) Hydradephaga, (c) coprophagous Scarabaeoidea, (d) phytophagous Scarabaeoidea, (e) total Scarabaeoidea, (f) Tenebrionidae. Solid lines: expected rates from the species–area relationship. Dashed lines: expected rates from multiple regression models.
Figure 3.
Extinction rates (% of extinct species) expected on the basis of area loss (% of area lost) for various beetle groups in Italian reserves: (a) Carabidae, (b) Hydradephaga, (c) coprophagous Scarabaeoidea, (d) phytophagous Scarabaeoidea, (e) total Scarabaeoidea, (f) Tenebrionidae. Solid lines: expected rates from the species–area relationship. Dashed lines: expected rates from multiple regression models.

Figure 4.
Number of extinct species expected on the basis of area loss (% of area lost) for the Carabidae of Italian reserves for areas hypothetically hosting 10, 20, 40, 60, 80, and 100 species. Curves were calculated using z = 0.212, as obtained from the power function of the species–area relationship. Use of z-values from multiple models did not change the pattern. Note the log-scale for the y-axis.
Figure 4.
Number of extinct species expected on the basis of area loss (% of area lost) for the Carabidae of Italian reserves for areas hypothetically hosting 10, 20, 40, 60, 80, and 100 species. Curves were calculated using z = 0.212, as obtained from the power function of the species–area relationship. Use of z-values from multiple models did not change the pattern. Note the log-scale for the y-axis.


{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Table 1.
Number of species for various beetle groups in Italian reserves. Res: Reserve number 1 (reserves are numbered as in Figure 1); Lat: Latitude (decimal degrees); Lon: Longitude (decimal degrees); Area: reserve size (km2); Min: Minimum elevation (m); Max: Maximum elevation (m); Range: Elevational range (m); Mean: Mean elevation (m). Beetle groups: Ca: Carabidae; Hy: Hydradephaga; Co: coprophagous Scarabaeoidea; Ph: phytophagous Scarabaeoidea; Sc: all Scarabaeoidea; Te: Tenebrionidae. For reserve 8, elevational data without parentheses refer to Carabidae sampling points, and those in parentheses to Hydradephaga sampling points.
Table 1.
Number of species for various beetle groups in Italian reserves. Res: Reserve number 1 (reserves are numbered as in Figure 1); Lat: Latitude (decimal degrees); Lon: Longitude (decimal degrees); Area: reserve size (km2); Min: Minimum elevation (m); Max: Maximum elevation (m); Range: Elevational range (m); Mean: Mean elevation (m). Beetle groups: Ca: Carabidae; Hy: Hydradephaga; Co: coprophagous Scarabaeoidea; Ph: phytophagous Scarabaeoidea; Sc: all Scarabaeoidea; Te: Tenebrionidae. For reserve 8, elevational data without parentheses refer to Carabidae sampling points, and those in parentheses to Hydradephaga sampling points.
| Res | Lat | Lon | Area | Min | Max | Range | Mean | Ca | Hy | Co | Ph | Sc | Te |
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| 1 | 46.03 | 8.48 | 146 | 400 | 2200 | 1820 | 1116.5 | 131 | - | - | - | - | - |
| 2 | 46.02 | 11.98 | 1 | 225 | 233 | 8 | 229 | - | - | 16 | 19 | 36 | 1 |
| 3 | 45.31 | 8.93 | 983 | 116 | 300 | 184 | 208 | 249 | 63 | 12 | 33 | 51 | 13 |
| 4 | 45.20 | 10.74 | 2.3 | 22 | 26 | 4 | 24 | 71 | 40 | 4 | 16 | 24 | 14 |
| 5 | 45.04 | 11.24 | 1.3 | 7 | 15 | 8 | 14 | - | - | 0 | 5 | 6 | 6 |
| 6 | 44.49 | 9.42 | 0.17 | 1326 | 1350 | 24 | 1330.5 | 23 | 10 | 0 | 5 | 5 | - |
| 7 | 44.40 | 10.02 | 2.9 | 1330 | 1470 | 140 | 1406.5 | 24 | 12 | - | - | - | - |
| 8 | 43.83 | 11.73 | 368.5 | 550 (350) | 1200 (1450) | 650 (1106) | 868 (753.5) | 198 | 48 | - | - | - | - |
| 9 | 42.40 | 11.21 | 4 | 0 | 3 | 3 | 1.5 | 38 | 1 | 1 | 2 | 5 | 14 |
| 10 | 42.16 | 13.82 | 100 | 300 | 400 | 100 | 350 | 87 | - | 14 | 13 | 28 | 16 |
| 11 | 41.98 | 12.67 | 5.4 | 50 | 120 | 70 | 85 | 77 | 13 | 22 | 12 | 37 | 13 |
| 12 | 41.93 | 15.11 | 9.6 | 0 | 1 | 1 | 0.5 | - | - | - | - | - | 10 |
| 13 | 41.81 | 13.79 | 1040 | 700 | 2249 | 1549 | 1474.5 | 243 | - | 69 | 18 | 95 | - |
| 14 | 41.81 | 15.85 | 1700 | 0 | 950 | 950 | 456 | 233 | 55 | 70 | 25 | 105 | 61 |
| 15 | 41.70 | 12.38 | 70 | 0 | 70 | 70 | 35 | 179 | 69 | 62 | 27 | 104 | 44 |
| 16 | 41.34 | 13.04 | 88 | 0 | 541 | 541 | 270.5 | - | 34 | - | - | - | 36 |
| 17 | 40.95 | 15.63 | 66 | 650 | 1326 | 676 | 988 | 172 | 25 | 55 | 16 | 73 | 17 |
| 18 | 40.82 | 14.43 | 84.8 | 200 | 1281 | 1081 | 740.5 | 32 | - | 8 | 11 | 19 | 23 |
| 19 | 40.59 | 15.75 | 1.6 | 764 | 770 | 6 | 767 | 109 | 30 | 21 | 10 | 33 | 9 |
| 20 | 40.18 | 16.70 | 5 | 0 | 6 | 6 | 3 | 253 | 69 | 54 | 28 | 97 | 58 |
| 21 | 39.97 | 16.21 | 1100 | 800 | 2000 | 1200 | 1482 | 220 | 56 | 75 | 29 | 106 | 35 |
| 22 | 39.32 | 16.58 | 1250 | 1000 | 1900 | 900 | 1425 | 201 | 55 | 65 | 29 | 96 | 36 |
| 23 | 37.89 | 14.00 | 399.4 | 400 | 1979 | 1579 | 1189.5 | - | - | 81 | 35 | 129 | 39 |
1 Reserve names: 1: Val Grande National Park; 2: Vincheto di Celarda Nature Reserve; 3: Ticino Regional Park; 4: Bosco Fontana Nature Reserve; 5: Isola Boscone Nature Reserve; 6: Agoraie Nature Reserve; 7: Guadine Pradaccio Nature Reserve; 8: Foreste Casentinesi National Park; 9: Burano Nature Reserve; 10: Pescara Spring Nature Reserve and adjacent areas in the Valle Peligna; 11: Inviolata Archeological and Nature Park; 12: Foce Saccione Site of Community Importance; 13: Abruzzo, Latium and Molise National Park; 14: Gargano National Park and Isola Varano Nature Reserve; 15: Castelporziano Presidential Estate and Castelfusano Urban Park; 16: Circeo National Park; 17: Vulture Natural Park; 18: Vesuvius National Park; 19: Pantano di Pignola Reserve; 20: Policoro Reserve; 21: Pollino National Park; 22: Sila National Park; 23: Madonie Regional Natural Park.
Table 2.
Species–area relationships (SARs) for beetles in Italian reserves. SARs were modelled using the linearized version of the power function with decimal logarithms: log(S) = log(c) + z log (A), where S is the species number, A is area, and c and z are fitting parameters. SE: standard error, t = Student’s t, p = probability, R2 = goodness of fit.
Table 2.
Species–area relationships (SARs) for beetles in Italian reserves. SARs were modelled using the linearized version of the power function with decimal logarithms: log(S) = log(c) + z log (A), where S is the species number, A is area, and c and z are fitting parameters. SE: standard error, t = Student’s t, p = probability, R2 = goodness of fit.
| Beetle Group, R2 Values, and Estimated Parameters | Estimate ± SE | t | p |
|---|---|---|---|
| Carabidae (R2 = 0.50) | |||
| log(c) | 1.68 ± 0.11 | 15.65 | <<0.001 |
| z | 0.21 ± 0.05 | 4.03 | <0.001 |
| Hydradephaga (R2 = 0.47) | |||
| log(c) | 1.30 ± 0.10 | 13.65 | <<0.001 |
| z | 0.15 ± 0.05 | 3.26 | 0.007 |
| Scarabaeoidea (coprophagous) (R2 = 0.49) | |||
| log(c) | 0.73 ± 0.18 | 4.15 | <<0.001 |
| z | 0.35 ± 0.09 | 3.93 | 0.001 |
| Scarabaeoidea (phytophagous) (R2 = 0.44) | |||
| log(c) | 0.92 ± 0.10 | 9.41 | <<0.001 |
| z | 0.17 ± 0.05 | 3.37 | 0.004 |
| Scarabaeoidea (total) (R2 = 0.51) | |||
| log(c) | 1.19 ± 0.13 | 9.44 | <<0.001 |
| z | 0.26 ± 0.06 | 4.08 | <0.001 |
| Tenebrionidae (R2 = 0.42) | |||
| log(c) | 0.87 ± 0.14 | 6.26 | <<0.001 |
| z | 0.25 ± 0.07 | 3.38 | 0.004 |
Table 3.
Best fit models for the influence of area, latitude, and elevation on beetle richness in Italian reserves. Min: Minimum elevation (m); Range: Elevational range (m); df: degrees of freedom; R2adj: adjusted R2; AICc: corrected Akaike Information Criterion. Errors refer to standard errors. * = p <0.05, ** = p < 0.01, *** p < 0.001.
Table 3.
Best fit models for the influence of area, latitude, and elevation on beetle richness in Italian reserves. Min: Minimum elevation (m); Range: Elevational range (m); df: degrees of freedom; R2adj: adjusted R2; AICc: corrected Akaike Information Criterion. Errors refer to standard errors. * = p <0.05, ** = p < 0.01, *** p < 0.001.
| Intercept | Area | Latitude | Min | Range | df | R2adj | AICc | |
|---|---|---|---|---|---|---|---|---|
| Carabidae | ||||||||
| 1.90 ± 0.15 (***) | 0.34 ± 0.08 (***) | −0.20 ± 0.10 | 4 | 0.56 | 7.7 | |||
| 1.68 ± 0.11 (***) | 0.21 ± 0.05 (***) | 3 | 0.47 | 8.9 | ||||
| Hydradephaga | ||||||||
| 1.71 ± 0.09 (***) | 0.34 ± 0.04 (***) | −0.34 ± 0.06 (***) | 4 | 0.84 | −11.2 | |||
| Scarabaeoidea (coprophagous) | ||||||||
| 19.74 ± 8.21 (*) | 0.25 ± 0.09 (*) | −11.61 ± 5.01 (*) | 4 | 0.58 | 27.8 | |||
| Scarabaeoidea (phytophagous) | ||||||||
| 0.92 ± 0.10 (***) | 0.17 ± 0.05 (**) | 3 | 0.38 | 8.7 | ||||
| Scarabaeoidea (total) | 1.19 ± 0.13 (***) | 0.26 ± 0.06 (***) | 3 | 0.48 | 18.0 | |||
| Tenebrionidae | ||||||||
| 17.97 ± 4.14 (**) | 0.19 ± 0.05 (**) | −10.34 ± 2.52 (**) | −0.16 ± 0.04 (**) | 5 | 0.76 | 5.2 |
© 2020 by the author. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
MDPI and ACS Style
Fattorini, S. Beetle Species–Area Relationships and Extinction Rates in Protected Areas. Insects 2020, 11, 646. https://doi.org/10.3390/insects11090646
AMA Style
Fattorini S. Beetle Species–Area Relationships and Extinction Rates in Protected Areas. Insects. 2020; 11(9):646. https://doi.org/10.3390/insects11090646
Chicago/Turabian StyleFattorini, Simone. 2020. "Beetle Species–Area Relationships and Extinction Rates in Protected Areas" Insects 11, no. 9: 646. https://doi.org/10.3390/insects11090646
Note that from the first issue of 2016, this journal uses article numbers instead of page numbers. See further details here.