Bmapaf-1 is Involved in the Response against BmNPV Infection by the Mitochondrial Apoptosis Pathway


Abstract
:Simple Summary
Abstract
1. Introduction
2. Materials and Methods
2.1. Silkworm and BmNPV
2.2. Bioinformatics Analysis
2.3. Sample Preparation, RNA Extraction, and cDNA Synthesis
2.4. RT-qPCR
2.5. Synthesis of siRNA
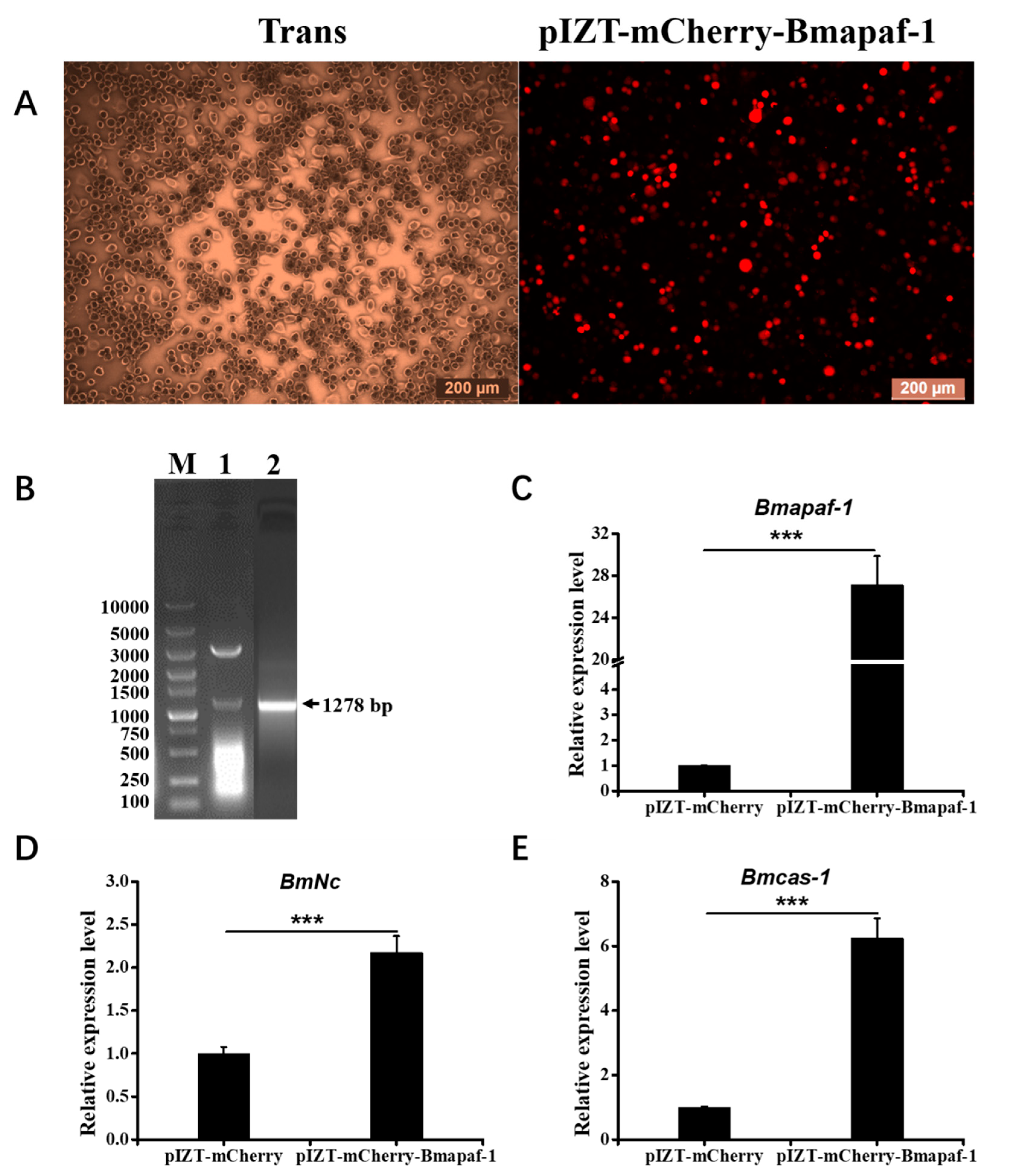
2.6. Construction of pIZT-mCherry-Bmapaf-1 Overexpression Vector
2.7. BmN Cell Culture, Transfection, and Fluorescence Signal Acquisition
2.8. Inhibition and Induction of Apoptosis
3. Results
3.1. Characterization of the BmApaf-1 Sequence
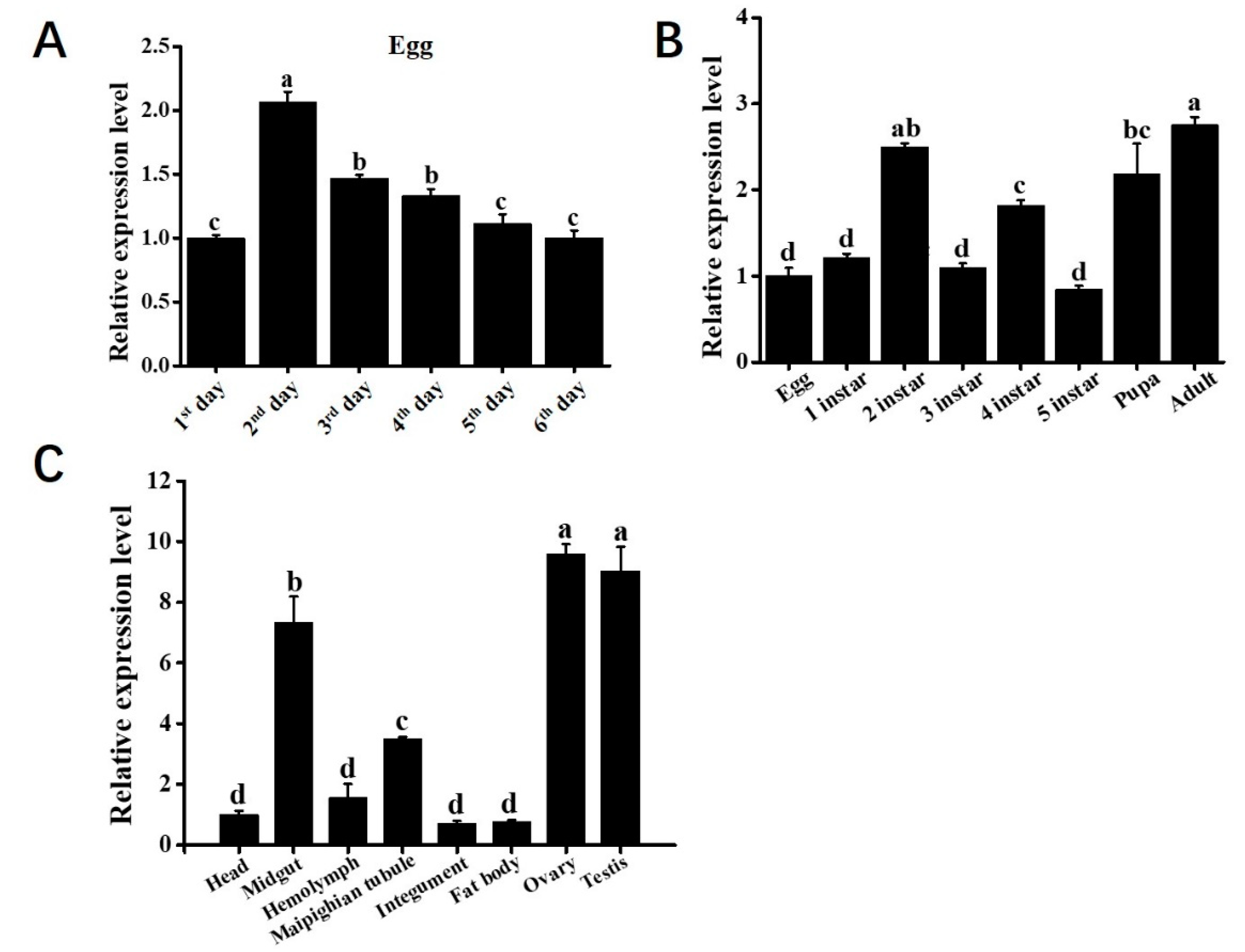
3.2. The Spatiotemporal Expression Pattern of Bmapaf-1
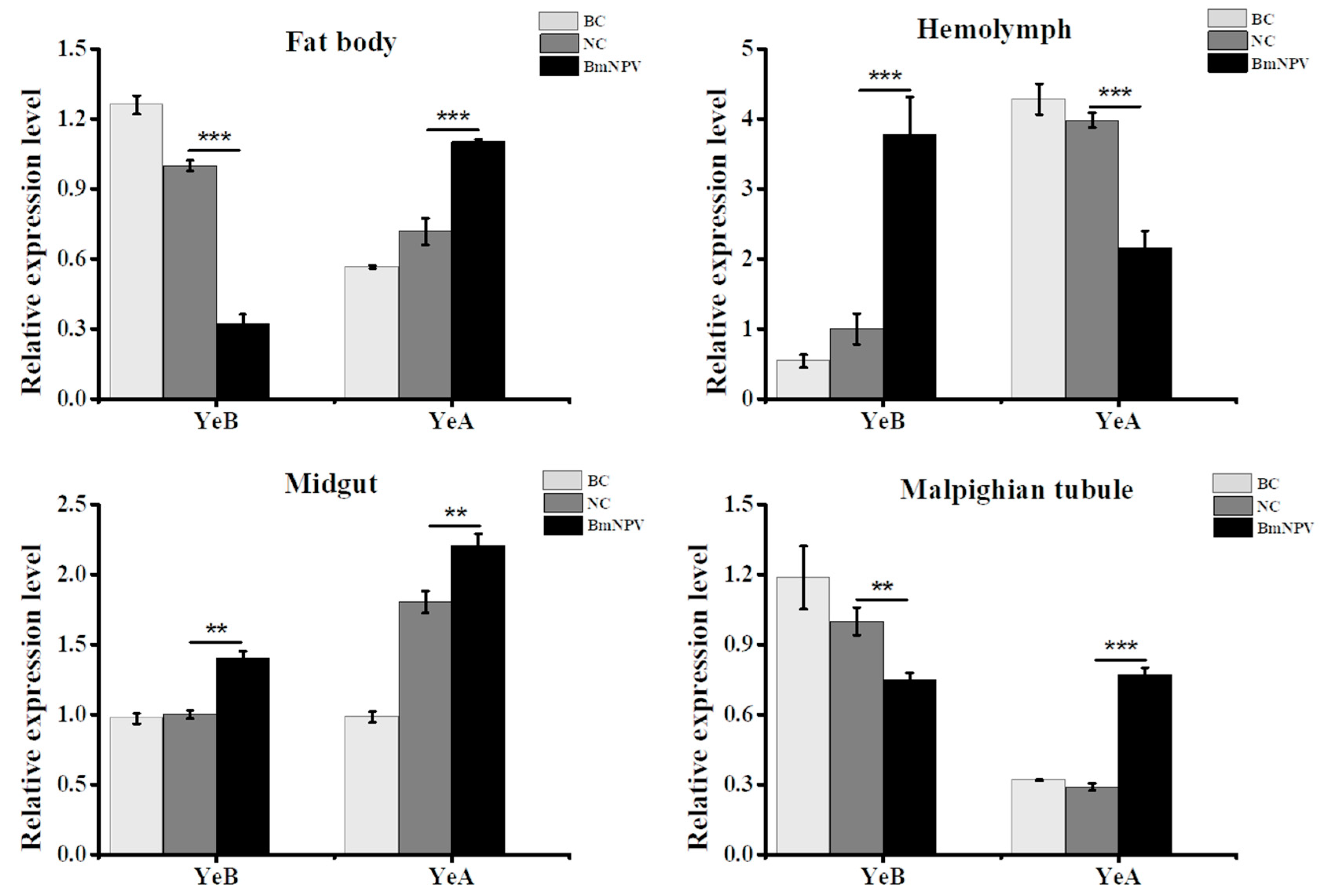
3.3. Bmapaf-1 Showed Significant Response to BmNPV Infection in Different Tissues
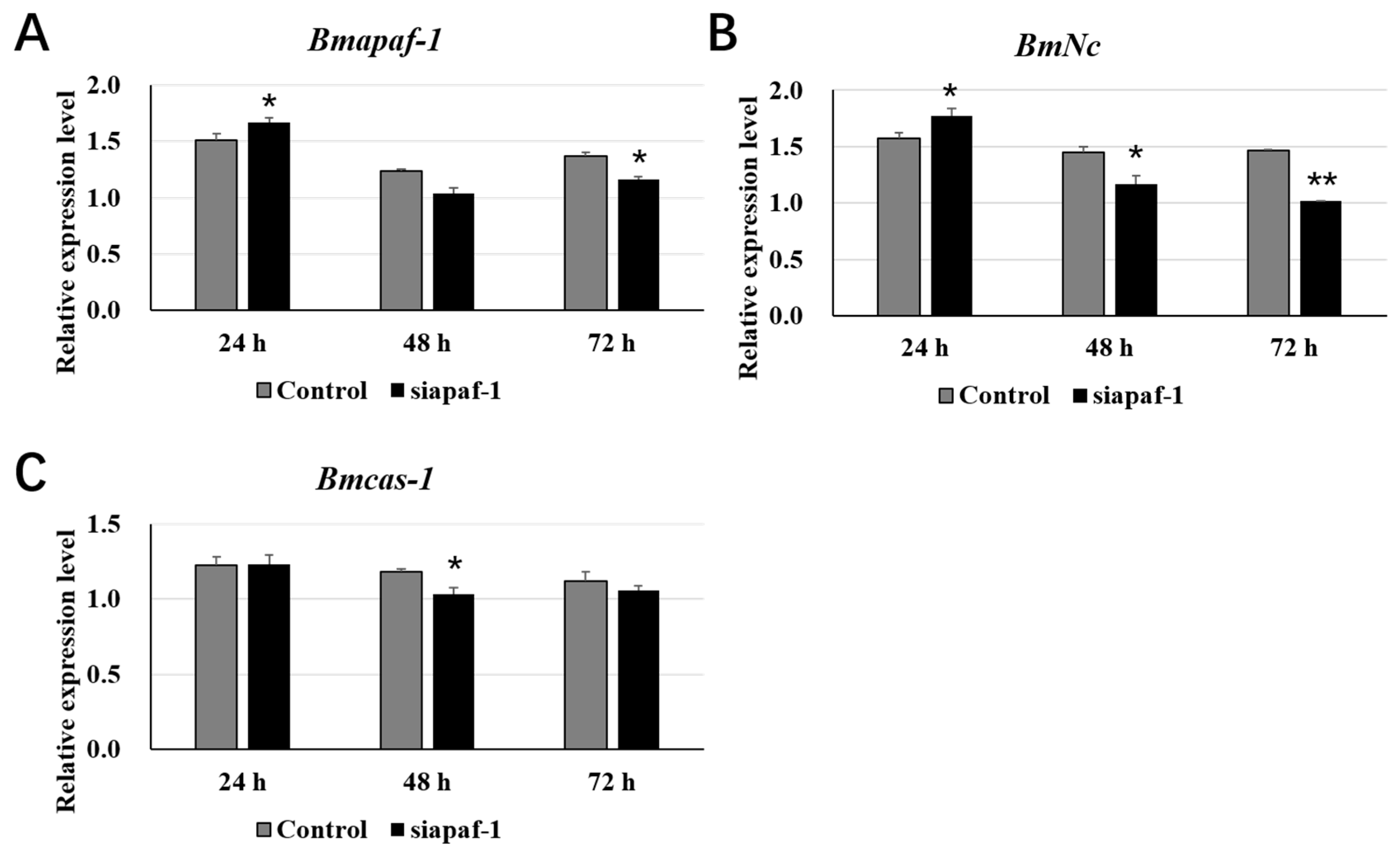
3.4. Selected Downstream Genes Were Downregulated after Knockdown of Bmapaf-1 in BmN Cells
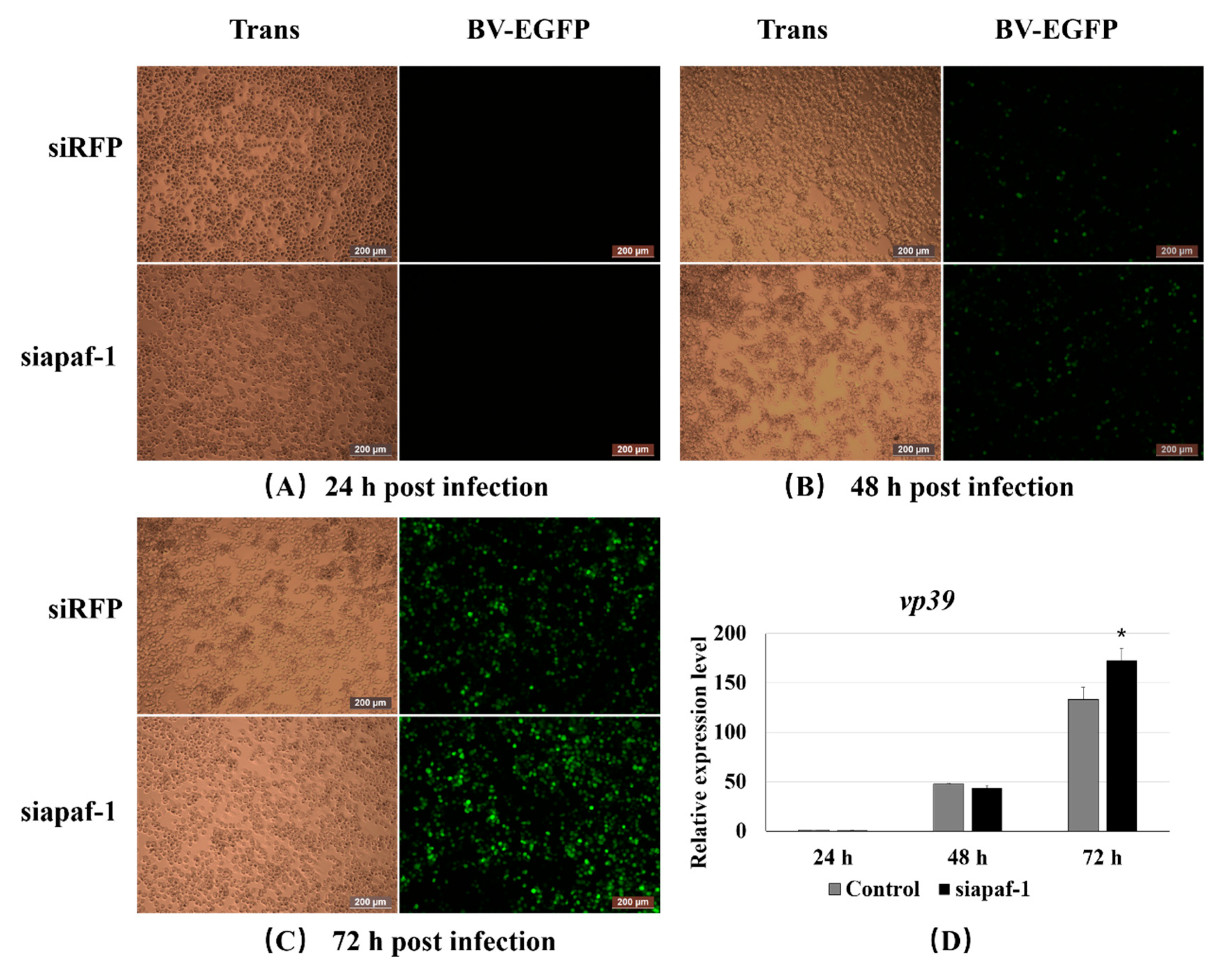
3.5. Knockdown of Bmapaf-1 Promoted BmNPV Infection in BmN Cells
3.6. Overexpression of Bmapaf-1 Upregulated the Expression of Its Downstream Genes in BmN Cells
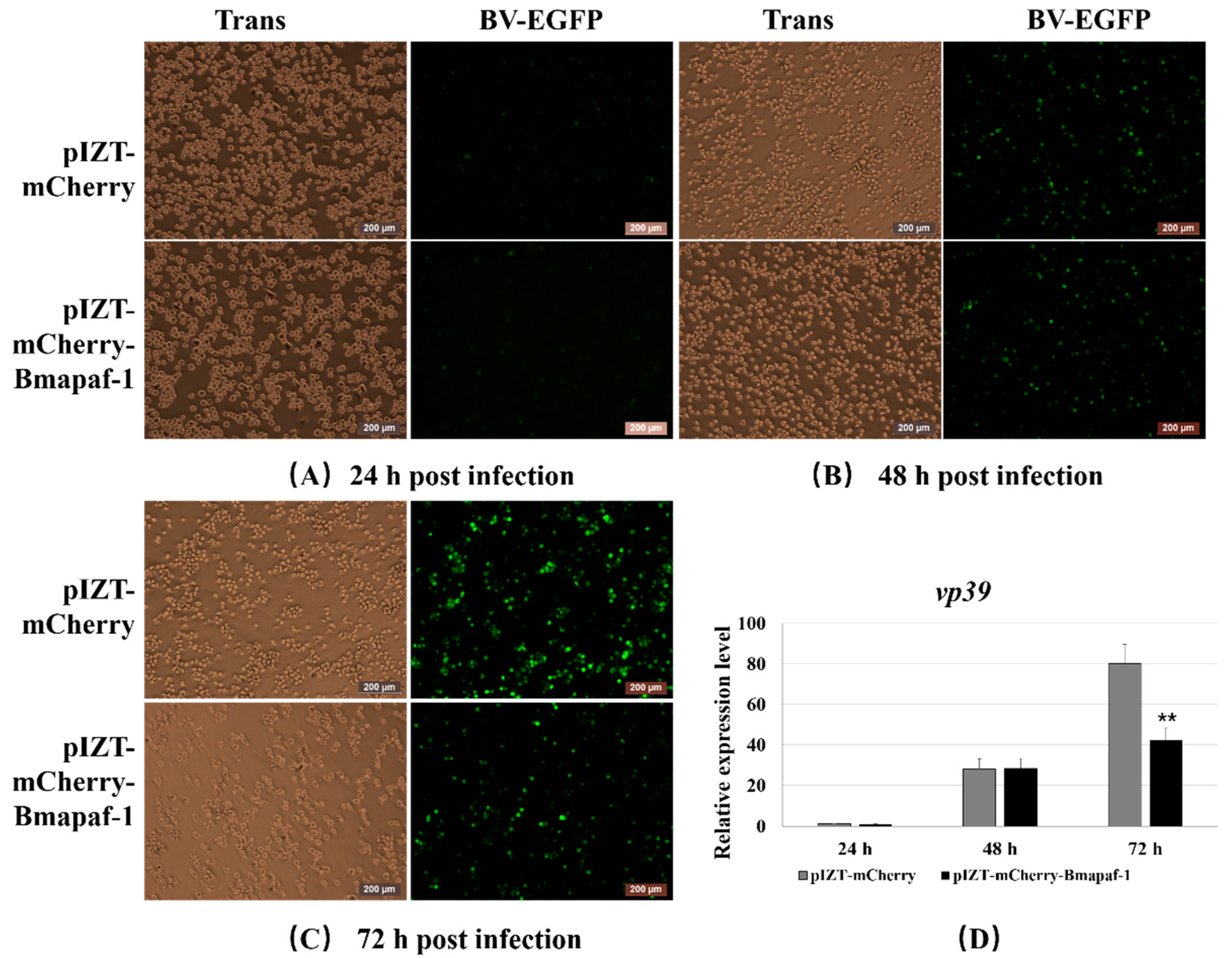
3.7. Overexpression of Bmapaf-1 Inhibited BmNPV Infection in BmN Cells
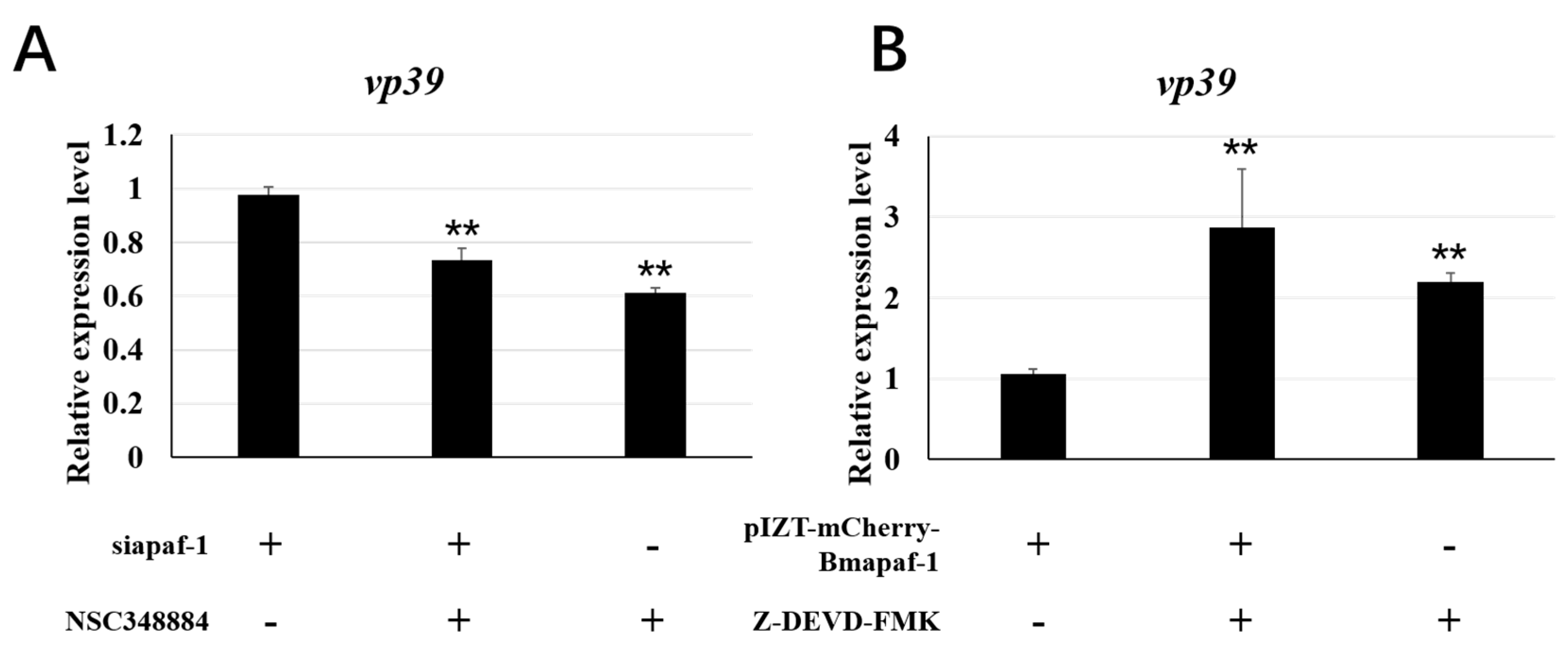
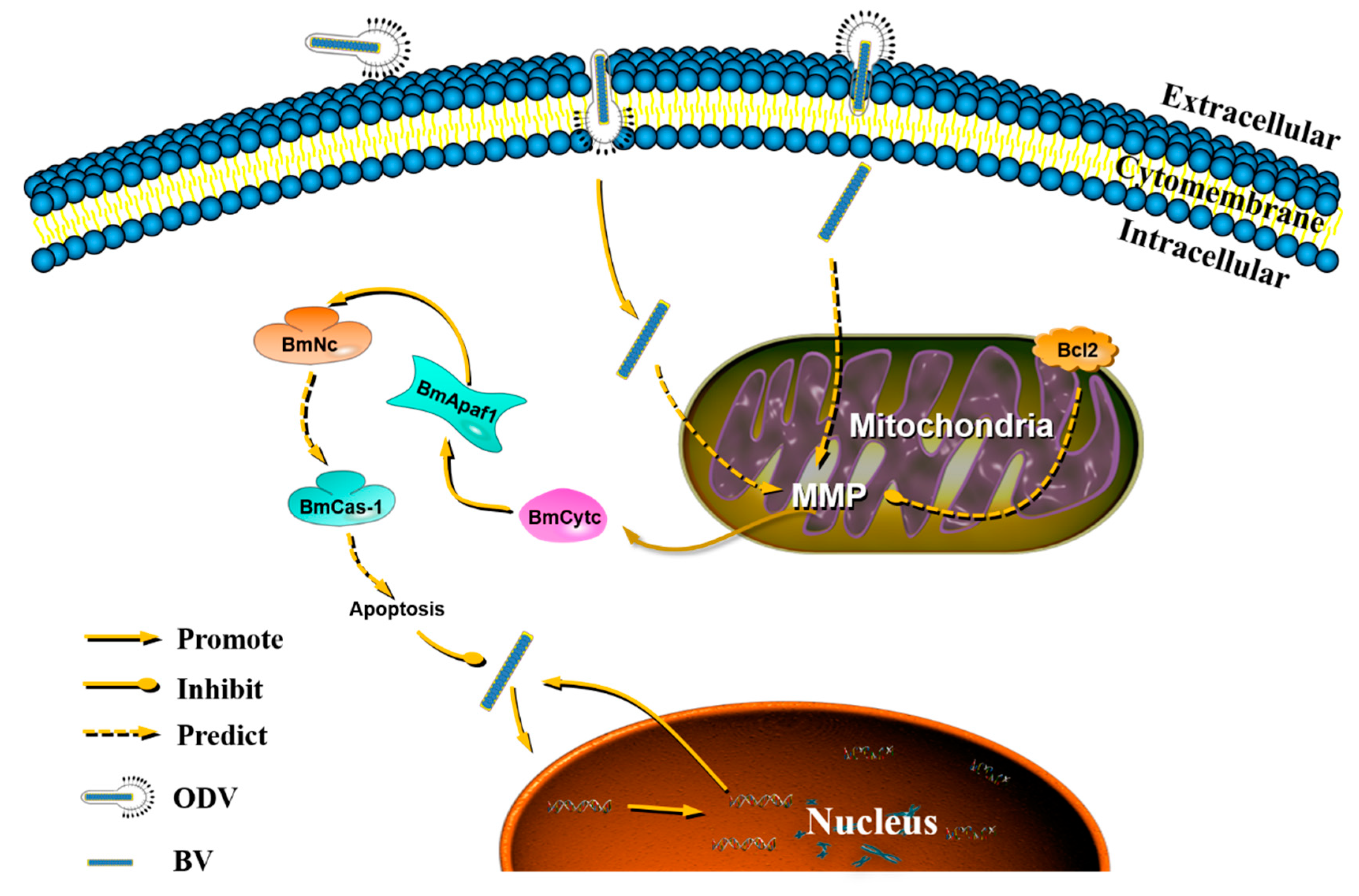
3.8. Apoptosis Regulated by Bmapaf-1 Involved in Response against BmNPV ISnfection
4. Discussions
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Cheng, Y.; Wang, X.; Du, C.; Gao, J.; Xu, J. Expression Analysis of Several Antiviral Related Genes to BmNPV in Different Resistant Strains of Silkworm, Bombyx mori. J. Insect Sci. 2014, 14, 76. [Google Scholar] [CrossRef] [PubMed]
- Wang, X.Y.; Shao, Z.M.; Zhang, Y.J.; Vu, T.T.; Wu, Y.C.; Xu, J.P.; Deng, M.J. A (1)H NMR based study of hemolymph metabonomics in different resistant silkworms, Bombyx mori (Lepidotera), after BmNPV inoculation. J. Insect Physiol. 2019, 117, 103911. [Google Scholar] [CrossRef] [PubMed]
- Wang, X.Y.; Yu, H.Z.; Geng, L.; Xu, J.P.; Yu, D.; Zhang, S.Z.; Ma, Y.; Fei, D.Q. Comparative Transcriptome Analysis of Bombyx mori (Lepidoptera) Larval Midgut Response to BmNPV in Susceptible and Near-Isogenic Resistant Strains. PLoS ONE 2016, 11, e0155341. [Google Scholar] [CrossRef] [Green Version]
- Li, G.; Qian, H.; Luo, X.; Xu, P.; Yang, J.; Liu, M.; Xu, A. Transcriptomic analysis of resistant and susceptible Bombyx mori strains following BmNPV infection provides insights into the antiviral mechanisms. Int. J. Genom. 2016, 2016, 2086346. [Google Scholar]
- Yu, H.; Wang, X.; Xu, J.; Ma, Y.; Zhang, S.; Yu, D.; Fei, D.; Muhammad, A. iTRAQ-based quantitative proteomics analysis of molecular mechanisms associated with Bombyx mori (Lepidoptera) larval midgut response to BmNPV in susceptible and near-isogenic strains. J. Proteom. 2017, 165, 35–50. [Google Scholar] [CrossRef] [PubMed]
- Zhang, S.Z.; Wang, J.; Zhu, L.B.; Toufeeq, S.; Xu, X.; You, L.L.; Li, B.; Hu, P.; Xu, J.P. Quantitative label-free proteomic analysis reveals differentially expressed proteins in the digestive juice of resistant versus susceptible silkworm strains and their predicted impacts on BmNPV infection. J. Proteom. 2020, 210, 103527. [Google Scholar] [CrossRef]
- Smith, C.A.; Williams, G.T.; Kingston, R.; Jenkinson, E.J.; Owen, J.J.T. Apoptosis. Nature 1989, 338, 10. [Google Scholar] [CrossRef]
- Kvansakul, M. Viral Infection and Apoptosis. Viruses 2017, 9, 356. [Google Scholar] [CrossRef] [Green Version]
- Pradelli, L.A.; Bénéteau, M.; Ricci, J.-E. Mitochondrial control of caspase-dependent and -independent cell death. Cell. Mol. Life Sci. 2010, 67, 1589–1597. [Google Scholar] [CrossRef]
- Hakem, R.; Hakem, A.; Duncan, G.S.; Henderson, J.T.; Mak, T.W. Differential Requirement for Caspase 9 in Apoptotic Pathways In Vivo. Cell 1998, 94, 339–352. [Google Scholar] [CrossRef] [Green Version]
- Kuida, K.; Haydar, T.F.; Kuan, C.Y.; Gu, Y.; Flavell, R.A. Reduced Apoptosis and Cytochrome c–Mediated Caspase Activation in Mice Lacking Caspase 9. Cell 1998, 94, 325–337. [Google Scholar] [CrossRef] [Green Version]
- Li, K.; Li, Y.; Shelton, J.M.; Richardson, J.A.; Spencer, E.; Chen, Z.J.; Wang, X.; Williams, R.S. Cytochrome c Deficiency Causes Embryonic Lethality and Attenuates Stress-Induced Apoptosis. Cell 2000, 101, 389–399. [Google Scholar] [CrossRef] [Green Version]
- Clavier, A.; Rincheval-Arnold, A.; Colin, J.; Mignotte, B.; Guenal, I. Apoptosis in Drosophila: Which role for mitochondria? Apoptosis 2016, 21, 239–251. [Google Scholar] [CrossRef]
- Mohamad, N.; Gutierrez, A.; Nunez, M.; Cocca, C.; Martin, G.; Cricco, G.; Medina, V.; Rivera, E.; Bergoc, R. Mitochondrial apoptotic pathways. Biocell 2005, 29, 149–161. [Google Scholar] [CrossRef]
- Saleh, A. Negative regulation of the Apaf-1 apoptosome by Hsp70. Nat. Cell Biol. 2000, 2, 476–483. [Google Scholar] [CrossRef]
- Acehan, D.; Jiang, X.; Morgan, D.G.; Heuser, J.E.; Wang, X.; Akey, C.W. Three-Dimensional Structure of the Apoptosome: Implications for Assembly, Procaspase-9 Binding, and Activation. Mol. Cell 2002, 9, 423–432. [Google Scholar] [CrossRef]
- Adams, J.M.; Cory, S. Apoptosomes: Engines for caspase activation. Curr. Opin. Cell Biol. 2002, 14, 715. [Google Scholar] [CrossRef]
- Salvesen, G.S.; Abrams, J.M. Caspase activation-Stepping on the gas or releasing the brakes? Lessons from humans and flies. Oncogene 2004, 23, 2774–2784. [Google Scholar] [CrossRef] [Green Version]
- Wang, X. The expanding role of mitochondria in apoptosis. Genes Dev. 2001, 15, 2922–2933. [Google Scholar]
- Meyer, K.; Basu, A.; Saito, K.; Ray, R.B.; Ray, R. Inhibition of hepatitis C virus core protein expression in immortalized human hepatocytes induces cytochrome c-independent increase in Apaf-1 and caspase-9 activation for cell death. Virology 2005, 336, 198–207. [Google Scholar] [CrossRef] [Green Version]
- Wang, X.Y.; Shao, Z.M.; Chen, Q.Y.; Xu, J.P.; Sun, X.; Xu, Z.P.; Li, M.W.; Wu, Y.C. Knockdown of BmTCP-1beta Delays BmNPV Infection in vitro. Front. Microbiol. 2019, 10, 578. [Google Scholar] [CrossRef]
- Guo, H.; Jiang, L.; Xia, Q. Selection of reference genes for analysis of stress-responsive genes after challenge with viruses and temperature changes in the silkworm Bombyx mori. Mol. Genet. Genom. 2015, 291, 999–1004. [Google Scholar] [CrossRef]
- Yin, J.; Zhang, J.; Li, T.; Sun, X.; Li, M.-W. BmSd gene regulates the silkworm wing size by affecting the Hippo pathway. Insect Sci. 2019, 27, 655–664. [Google Scholar] [CrossRef]
- Ye, Y.; Liu, X.I.; Xiao, M.; Zhang, Z.I.; Li, M.A. A palmitoyltransferase Approximated gene Bm-app regulates wing development in Bombyx mori. Insect Sci. 2018, 27, 2–13. [Google Scholar]
- Wang, X.Y.; Wu, K.H.; Pang, H.L.; Xu, P.Z.; Li, M.W.; Zhang, G.Z. Study on the Role of Cytc in Response to BmNPV Infection in Silkworm, Bombyx mori (Lepidoptera). Int. J. Mol. Sci. 2019, 20, 4325. [Google Scholar] [CrossRef] [Green Version]
- Kang, L.; Shi, H.; Liu, X.; Zhang, C.; Yao, Q.; Wang, Y.; Chang, C.; Shi, J.; Cao, J.; Kong, J. Arginine kinase is highly expressed in a resistant strain of silkworm (Bombyx mori, Lepidoptera): Implication of its role in resistance to Bombyx mori nucleopolyhedrovirus. Comp. Biochem. Physiol. Part B Biochem. Mol. Biol. 2011, 158, 230–234. [Google Scholar] [CrossRef]
- Duprez, L.; Wirawan, E.; Berghe, T.V.; Vandenabeele, P. Major cell death pathways at a glance. Microbes Infect. 2009, 11, 1050–1062. [Google Scholar] [CrossRef]
- Wang, Q.; Ju, X.; Chen, L.; Chen, K. Caspase-1 from the silkworm, Bombyx mori, is involved in Bombyx mori nucleopolyhedrovirus infection. Z. Nat. C J. Biosci. 2016, 72, 147–153. [Google Scholar] [CrossRef]
- Nagamine, T.; Sako, Y. A Role for the Anti-Viral Host Defense Mechanism in the Phylogenetic Divergence in Baculovirus Evolution. PLoS ONE 2016, 11, e0156394. [Google Scholar] [CrossRef] [Green Version]
- Long, G.; Pan, X.Y.; Kormelink, R.; Vlak, J.M. Functional entry of baculovirus into insect and mammalian cells is dependent on clathrin-mediated endocytosis. J. Virol. 2006, 80, 8830–8833. [Google Scholar] [CrossRef] [Green Version]
- Sedlic, F.; Wakatsuki, T.; Pravdic, D.; Bosnjak, Z. Mitochondrial membrane potential regulates production of reactive oxygen species and opening of mitochondrial permeability transition pore. FASEB J. 2009, 23, 576. [Google Scholar]
- Pan, M.H.; Chen, M.; Huang, S.J.; Yu, Z.S.; Lu, C. Cloning and protein release of cytochrome c in the apoptosis cells of silkworm. Sci. Agric. Sin. 2009, 42, 2546–2551. [Google Scholar]









{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Genes Name | Forward Primers (5′-3′) | Revers Primers (5′-3′) |
|---|---|---|
| Bmapaf-1 | TCACAACCCTCTAAAATCACACCAG | CGACAGCCAGTAATGGGTGTATGAG |
| BmNc | GAGGACGATGTGAGCAGGGAT | TTCAGCAGGAACGAAATGTAGC |
| Bmcas-1 | AACGGCAATGAAGACGAAGG | GGTGCCCGTGCGAGATTTTA |
| BmGAPDH | CCGCGTCCCTGTTGCTAAT | CTGCCTCCTTGACCTTTTGC |
| VP39 | CAACTTTTTGCGAAACGACTT | GGCTACACCTCCACTTGCTT |
| Bmapaf-1 KE | GGGGTACCAGGAAGCTGCTGCAGCA | CGGAATTCTATGTTTTCGACTTCGTTGAC |
| Primer Names | Sequences (5′-3′) |
|---|---|
| Bmapaf-1-1 Olig-1 | GATCACTAATACGACTCACTATAGGGGCTAATCTGGTCATAGTTATT |
| Bmapaf-1-1 Olig-2 | AATAACTATGACCAGATTAGCCCCTATAGTGAGTCGTATTAGTGATC |
| Bmapaf-1-1 Olig-3 | AAGCTAATCTGGTCATAGTTACCCTATAGTGAGTCGTATTAGTGATC |
| Bmapaf-1-1 Olig-4 | GATCACTAATACGACTCACTATAGGGTAACTATGACCAGATTAGCTT |
| Bmapaf-1-2 Olig-1 | GATCACTAATACGACTCACTATAGGGGCTAATTATCACCCGCAAATT |
| Bmapaf-1-2 Olig-2 | AATTTGCGGGTGATAATTAGCCCCTATAGTGAGTCGTATTAGTGATC |
| Bmapaf-1-2 Olig-3 | AAGCTAATTATCACCCGCAAACCCTATAGTGAGTCGTATTAGTGATC |
| Bmapaf-1-2 Olig-4 | GATCACTAATACGACTCACTATAGGGTTTGCGGGTGATAATTAGCTT |
| RFP-Olig-1 | GATCACTAATACGACTCACTATAGGGGCACCCAGACCATGAGAATTT |
| RFP-Olig-2 | AAATTCTCATGGTCTGGGTGCCCCTATAGTGAGTCGTATTAGTGATC |
| RFP-Olig-3 | AAGCACCCAGACCATGAGAATCCCTATAGTGAGTCGTATTAGTGATC |
| RFP-Olig-4 | GATCACTAATACGACTCACTATAGGGATTCTCATGGTCTGGGTGCTT |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Wang, X.-y.; Ding, X.-y.; Chen, Q.-y.; Zhang, K.-x.; Zhao, C.-x.; Tang, X.-d.; Wu, Y.-c.; Li, M.-w. Bmapaf-1 is Involved in the Response against BmNPV Infection by the Mitochondrial Apoptosis Pathway. Insects 2020, 11, 647. https://doi.org/10.3390/insects11090647
Wang X-y, Ding X-y, Chen Q-y, Zhang K-x, Zhao C-x, Tang X-d, Wu Y-c, Li M-w. Bmapaf-1 is Involved in the Response against BmNPV Infection by the Mitochondrial Apoptosis Pathway. Insects. 2020; 11(9):647. https://doi.org/10.3390/insects11090647
Chicago/Turabian StyleWang, Xue-yang, Xin-yi Ding, Qian-ying Chen, Kai-xiang Zhang, Chun-xiao Zhao, Xu-dong Tang, Yang-chun Wu, and Mu-wang Li. 2020. "Bmapaf-1 is Involved in the Response against BmNPV Infection by the Mitochondrial Apoptosis Pathway" Insects 11, no. 9: 647. https://doi.org/10.3390/insects11090647