Molecular Assessment of Genes Linked to Immune Response Traits of Honey Bees in Conventional and Organically Managed Apiaries


Abstract
:Simple Summary
Abstract
1. Introduction
2. Materials and Methods
2.1. Honey Bee Colonies and Honey Bee Samples
2.2. Protein Determination
2.3. RNA Extraction and CDNA Synthesis
2.4. Primer Design
2.5. Real-Time Quantitative Polymerase Chain Reaction
2.6. Data Analysis
3. Results
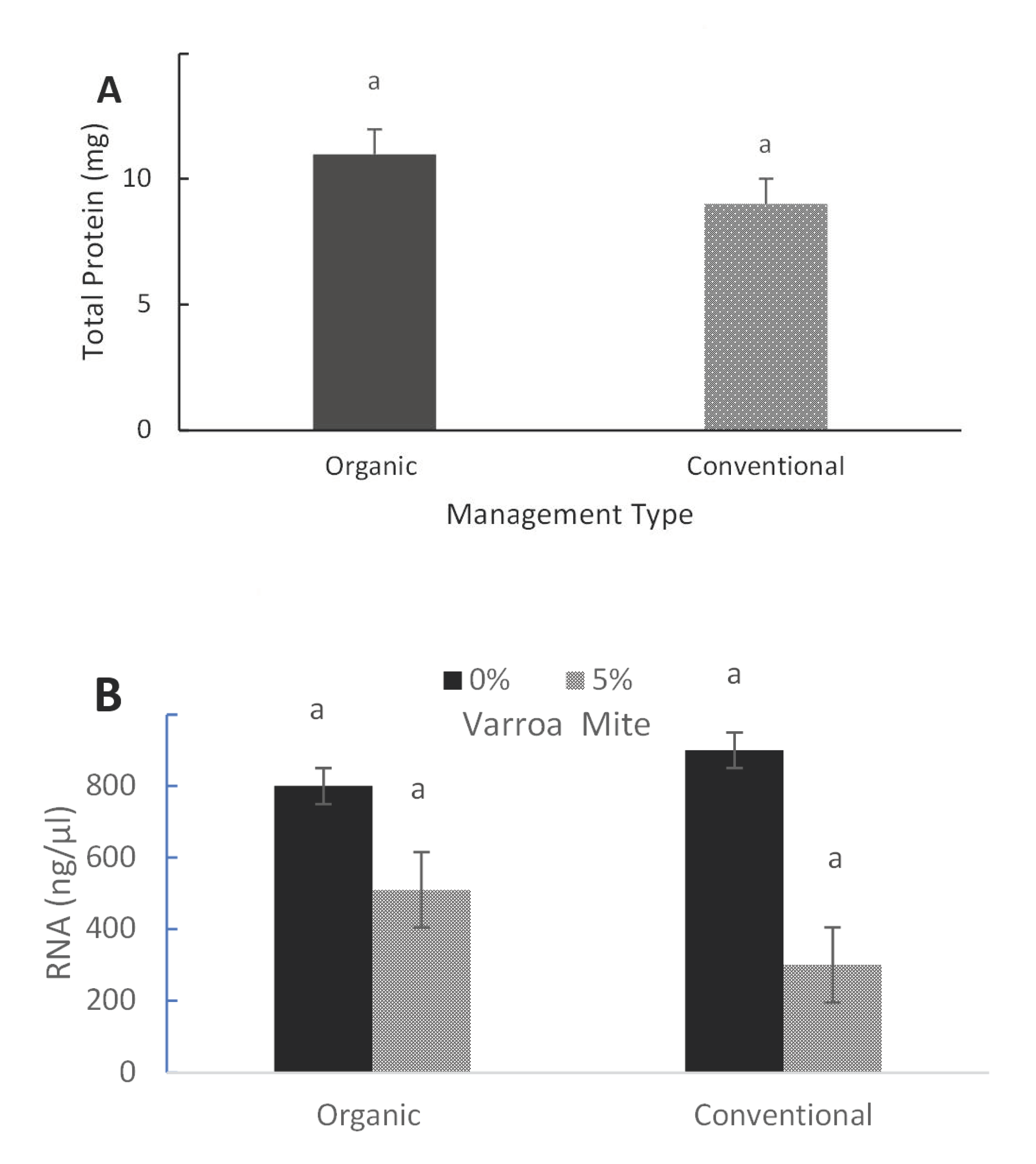
3.1. Honey Bee Protein and RNA Contents
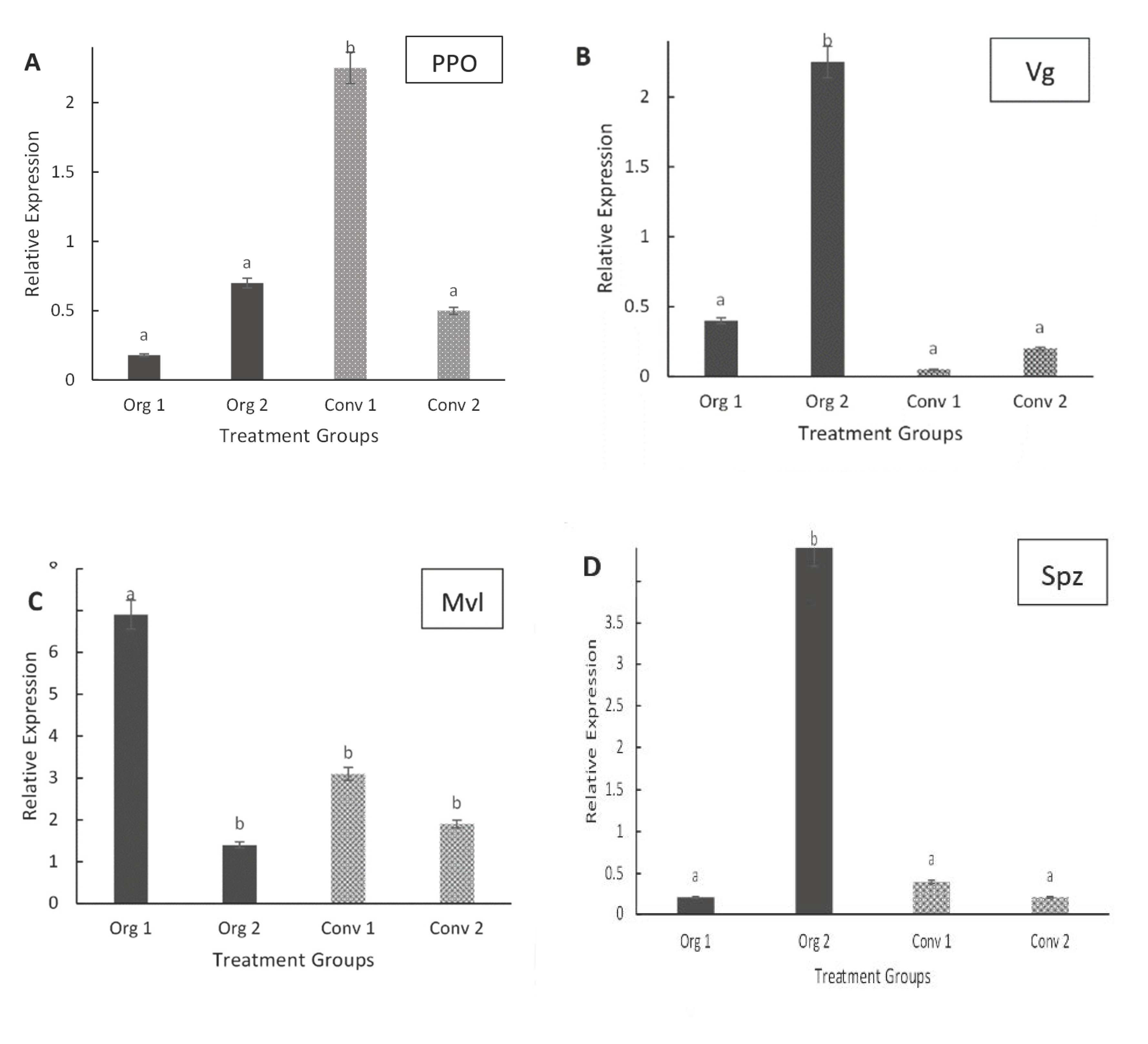
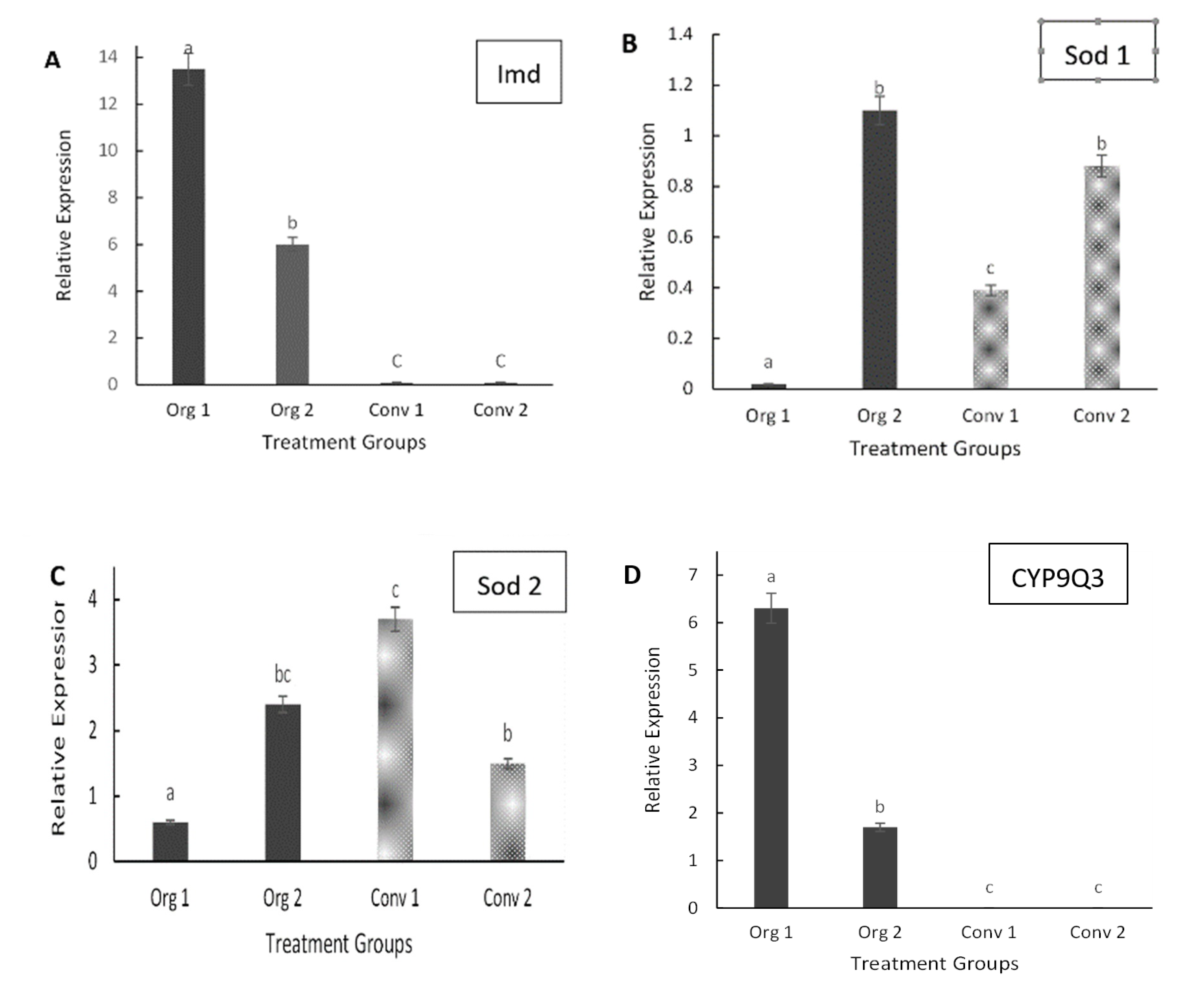
3.2. Gene Expression
4. Discussion
5. Conclusions
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Alaux, C.; Dantec, C.; Parrinello, H.; Le Conte, Y. Nutrigenomics in honey bees: Digital gene expression analysis of pollen’s nutritive effects on healthy and Varroa-parasitized bees. BMC Genom. 2011, 12, 496. Available online: https://bmcgenomics.biomedcentral.com/track/pdf/10.1186/1471-2164-12-496 (accessed on 5 March 2020).
- Gould, J.; Gould, C. The Honey Bee; Scientific American Library: New York, NY, USA, 1988. [Google Scholar]
- Brodschneider, R.; Crailsheim, K. Nutrition and health in honey bees. Apidologie 2010, 41, 278–294. [Google Scholar]
- Wahl, O.; Ulm, K. Influence of pollen feeding and physiological condition on pesticide sensitivity of the honey bee Apis mellifera carnica. Oecologia 1983, 59, 106–128. [Google Scholar] [PubMed]
- Frazier, M.; Mullin, C.; Frazier, C.J.; Ashcraft, S. What have pesticides got to do with it? Am. Bee J. 2008, 148, 521–523. [Google Scholar]
- Cox-Foster, D.; Conlan, S.; Holmes, E.C.; Palacios, G.; Evans, J.D.; Moran, A.; Quan, P.L.; Briese, T.; Hornig, M.; Geiser, M.D.; et al. A metagenomic survey of microbes in honey bee colony collapse disorder. PMID Sci. 2007, 318, 283–287. [Google Scholar] [CrossRef] [Green Version]
- Kraus, B.; Page, R.E. Population growth of Varroa jacobsoni Oud in Mediterranean climates of California. Apidologie 1995, 26, 149–157. [Google Scholar] [CrossRef]
- Finley, J.; Camazine, S.; Frazier, M. The epidemic of honey bee colony losses during the 1995–1996 season. Am. Bee J. 1996, 136, 805–808. [Google Scholar]
- Yang, X.; Cox-Foster, D.L. Impact of an ectoparasite on the immunity and pathology of an invertebrate: Evidence for host immunosuppression and viral amplification. Proc. Natl. Acad. Sci. USA 2005, 102, 7470–7475. [Google Scholar]
- Harbo, R.J.; Harris, J.W. Resistance to Varroa destructor (Mesostigmata: Varroidae) when mite-resistant queen honey bees (Hymenoptera: Apidae) were free-mated with unselected drones. J. Econ. Entomol. 2001, 94, 1319–1323. [Google Scholar] [CrossRef]
- Rinderer, T.; Lilia, E.; de Guzman, I.; Delatte, G.T.; Stelzer, J.A.; Lancaster, V.A.; Kuznetsov, V.; Beaman, L.; Watts, R.; Harrisa, J.W. Resistance to the parasitic mite Varroa destructor in honey bees from far-eastern Russia. Apidologie 2001, 32, 381–394. [Google Scholar] [CrossRef] [Green Version]
- Heintz, C.; Ribotto, M.; Ellis, M.; Delaplane, K.S. Best management practices for beekeepers pollinating California’s agricultural crops. Bee Culture 2011, 14, 17–19. [Google Scholar]
- USDA. Standards for the production of organic honey. In Formal Recommendation by the National Organic Standards Board to the National Organic Program; Certified Naturally Grown: Brooklyn, NY, USA, 2000; pp. 1–9. Available online: https://certified.naturallygrown.org/documents/HandbookNatBeek3ed.pdf (accessed on 23 July 2020).
- Kanga, L.H.B.; Jones, W.A.; James, R.R. Field trials using the fungal pathogen, Metarhizium anisopliae (Deuteromycetes: Hyphomycetes) to control the ectoparasitic mite, Varroa destructor (Acari: Varroidae) in honey bee, Apis mellifera (Hymenoptera: Apidae) colonies. J. Econ. Entomol. 2003, 96, 1091–1099. [Google Scholar] [CrossRef] [PubMed]
- Delaplane, K.S.; Hood, W.M. Effects of delayed acaricide treatment in honey bee colonies parasitized by Varroa jacobsoni and a late-season treatment threshold of the south-eastern. USA. J. Apic. Res. 1997, 36, 101–108. [Google Scholar] [CrossRef]
- Strange, J.P.; Sheppard, W.S. Optimum timing of miticide applications for control of Varroa destructor (Acari: Varroidae) in Apis mellifera (Hymenoptera: Apidae) in Washington State, USA. J. Econ. Entomol. 2001, 94, 1324–1331. [Google Scholar] [CrossRef]
- Kanga, L.H.B.; Siebert, S.C.; Sheikh, M.; Legaspi, J.C. Pesticide residues in conventionally and organically managed apiaries in South and North Florida. Curr. Investig. Agric. Curr. Res. 2019, 7, 937–940. [Google Scholar] [CrossRef] [Green Version]
- Van Engelsdorp, D.; Hayes, J.; Underwood, R.M.; Caron, D.; Pettis, J.A. Survey of managed honey bee colony losses in the USA, fall 2009 to winter 2010. J. Apicul. Res. 2011, 50, 1–10. [Google Scholar] [CrossRef] [Green Version]
- Smart, M.D.; Otto, C.R.V.; Lundgren, J.G. Nutritional status of honey bee (Apis mellifera L.) workers across an agricultural land-use gradient dataset. US Geological Survey Data Release. Sci. Rep. 2019. [Google Scholar] [CrossRef]
- Bradford, M.M. A rapid and sensitive method for the quantification of microgram quantities of protein utilizing the principle of protein-dye binding. Anal. Biochem. 1976, 72, 248–254. [Google Scholar] [CrossRef]
- Dubovskiy, I.M.; Martemyanov, V.V.; Vorontsova, Y.L.; Rantala, M.J.; Gryzanova, E.V.; Glupov, V.V. Effect of bacterial infection on antioxidant activity and lipid peroxidation in the midgut of Galleria mellonella L. larvae (Lepidoptera, Pyralidae). Comp. Biochem. Physiol. 2008, 148, 1–5. [Google Scholar] [CrossRef]
- Boncristiani, H.; Underwood, R.; Schwarz, R.; Evans, J.; Pettis, J.; van Engelsdorp, D. Direct effect of acaricides on pathogen loads and gene expression levels in honey bees Apis mellifera. J. Insect Physiol. 2012, 58, 613–620. [Google Scholar] [CrossRef]
- Frost, E.H.; Shutler, D.; Hillier, N.K. Effects of fluvalinate on honey bee learning, memory, responsiveness to sucrose, and survival. J. Exp. Biol. 2013, 216, 2931–2938. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Mao, W.; Schuler, M.A.; Berenbaum, M.R. Honey constituents up-regulate detoxification and immunity genes in the western honey bee Apis mellifera. Proc. Natl. Acad. Sci. USA 2013, 110, 8842–8846. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Scharlaken, B.; de Graaf, D.C.; Gossens, K.; Brunain, M.; Peelman, L.J.; Jacobs, F.J. Reference gene selection for insect expression studies using quantitative real-time PCR: The head of the honeybee, Apis mellifera, after a bacterial challenge. J. Insect Sci. 2009, 8, 33. [Google Scholar] [CrossRef] [Green Version]
- Livak, K.J.; Schmittgen, T.D. Analysis of relative gene expression data using real-time quantitative PCR and the 2(T) (-Delta Delta C) method. Methods 2001, 25, 402–408. [Google Scholar] [CrossRef]
- SAS Institute. SAS User’s Guide; Statisitical Analysis Systems: Cary, NC, USA, 2013. [Google Scholar]
- Ashida, M.; Brey, P. Recent advances in research on the insect prophenoloxidase cascade. In Molecular Mechanisms of Immune Responses in Insects; Brey, P.T., Hultmark, D., Eds.; Chapman & Hall: London, UK, 1997; pp. 135–172. [Google Scholar]
- Gregorc, A.; Evan, J.; Scharf, M.; Ellis, J. Gene expression in honey bee (Apis mellifera) larvae exposed to pesticides and Varroa mites (Varroa destructor). J. Insect Physiol. 2012, 58, 1042–1049. [Google Scholar] [CrossRef]
- Smodiš Škerl, M.I.; Bolta, S.V.; Cesnik, H.B.; Gregorc, A. Residues of pesticides in honeybee (Apis mellifera carnica) bee bread and in pollen loads from treated apple orchards. Bull. Environ. Contam. Toxicol. 2009, 83, 374–377. [Google Scholar] [CrossRef]
- Ben-Shahar, Y.; Dudek, N.L.; Robinson, G.E. Phenotypic deconstruction reveals involvement of manganese transporter malvolio in honey bee division of labor. J. Exp. Biol. 2004, 207, 3281–3288. [Google Scholar] [CrossRef] [Green Version]
- Eiri, D.M.; Nieh, J.C. A nicotinic acetylcholine receptor agonist affects honey bee sucrose responsiveness and decreases waggle dancing. J. Exp. Biol. 2012, 215, 2022–2029. [Google Scholar] [CrossRef] [Green Version]
- Evans, J.D.; Aronstein, K.; Chen, Y.P.; Hetru, C.; Imler, J.L.; Jiang, H.; Kanost, M.; Thompson, G.J.; Zou, Z.; Hultmark, D. Immune pathways and defense mechanisms in honey bees Apis mellifera. Insect Mol. Biol. 2006, 15, 645–656. [Google Scholar] [CrossRef] [Green Version]




{kind=link}
{kind=link}
{kind=link}
| Gene | BeeBase * | Forward Primer (5′ to 3′) | Reverse Primer (5’ to 3’) | Ta (°C) | Function |
|---|---|---|---|---|---|
| Prophenoloxidase (PPO) | GB18313- RA | GGA CAT CAA TCG ACA AGT TG | GAC GTC GAT TCC ATT TTT CT | 52 | Catalyzes cellular defense [24] |
| Vitellogenin (Vg) | GB13999 | CAC ATC CCT CGC CCT TCC AA | CAC GTA TCC CTC GTT GTC CAT | 51 | Immune function and longevity [24] |
| Malvolio (Mvl) | CCT TGG TAT AAA GAT TAT GAC AGG AAT ATG | CAA GAG CAC TGT GAA GAT ACA AGT TAT G | 57 | Involved in sucrose responsiveness [1,22] | |
| Spaetzle (Spz) | GB 15688 | TGC ACA AAT TGT TTT TCC TGA | GTC GTC CAT GAA ATC GAT CC | 49 | Member of the Toll-immune signaling pathway against fungi and bacteria [1,23] |
| Superoxide dismutase 1 (Sod) | GB 10133 | GAC TAA AGC AGT GTG CGT TC | TTA TCG CCA AAT TCA TGA AC | 52 | Codes for antioxidants; increase lifespan [1] |
| Superoxide mutase 2 (Sod 2) | GB14346 | CAA TGT TTG CAG CAA GAC GT | GTT GCA TGG TGC TTT GAA TG | 53 | Codes for antioxidants; increases lifespan [1] |
| Immune deficiency (Imd) | GB 18606 | TGT TAA CGA CCG ATG CAA AA | CAT CGC TCT TTT CGG ATG TT | 50 | Antimicrobial defense [1,22] |
| CYP9Q3 | GB19967 | GTTCCGGGAAAATGACTAC | GGTCAAAATGGTGGTGAC | 53 | Pesticide detoxification [21] |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Siebert, S.C.; Kanga, L.H.B.; Basha, S.M.; Legaspi, J.C. Molecular Assessment of Genes Linked to Immune Response Traits of Honey Bees in Conventional and Organically Managed Apiaries. Insects 2020, 11, 637. https://doi.org/10.3390/insects11090637
Siebert SC, Kanga LHB, Basha SM, Legaspi JC. Molecular Assessment of Genes Linked to Immune Response Traits of Honey Bees in Conventional and Organically Managed Apiaries. Insects. 2020; 11(9):637. https://doi.org/10.3390/insects11090637
Chicago/Turabian StyleSiebert, Shalom C., Lambert H. B. Kanga, Sheikh M. Basha, and Jesusa C. Legaspi. 2020. "Molecular Assessment of Genes Linked to Immune Response Traits of Honey Bees in Conventional and Organically Managed Apiaries" Insects 11, no. 9: 637. https://doi.org/10.3390/insects11090637