The First Record of Monochamus saltuarius (Coleoptera; Cerambycidae) as Vector of Bursaphelenchus xylophilus and Its New Potential Hosts in China


Abstract
:Simple Summary
Abstract
1. Introduction
2. Materials and Methods
2.1. Source and Preservation of Dead Trees
2.2. Experimental Conifers and Food Sources
2.3. Detection of PWN Carried by M. saltuarius
2.4. Experiments on Feeding Transmission by M. saltuarius
2.5. Data Analysis
3. Results
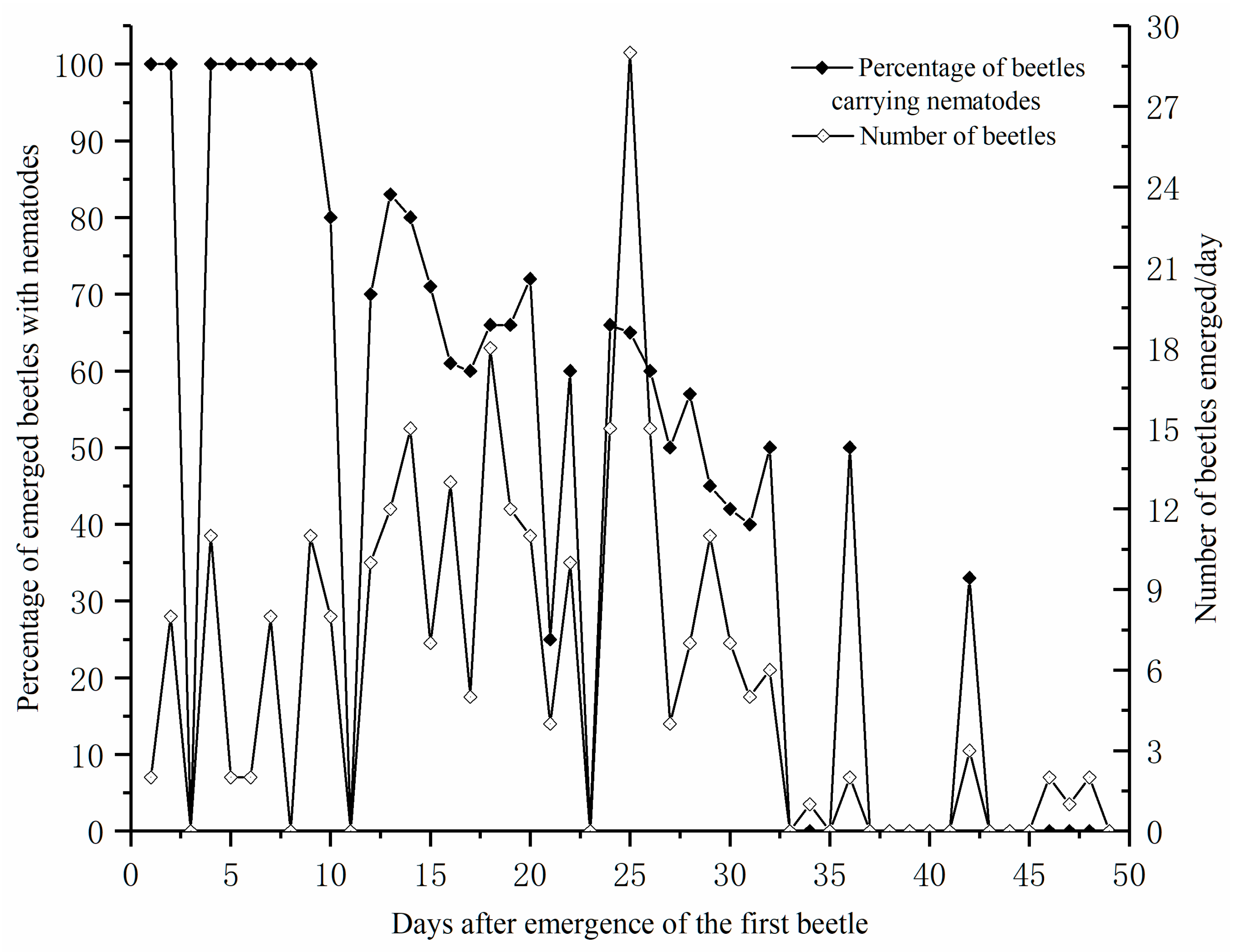
3.1. Emergence of M. saltuarius and Identification of Nematodes
3.2. Types of Transmission of PWNs into Pine Twigs through M. saltuarius Feeding
3.3. Experiment of M. saltuarius Feeding on Potted Seedlings
3.4. Transmission of PWNs by Mature M. saltuarius Feeding on Non-Pinus Conifer Tree Species
4. Discussion
5. Conclusions
Author Contributions
Funding
Conflicts of Interest
References
- Mamiya, Y. Pathology of the Pine Wilt Disease Caused by Bursaphelenchus xylophilus. Annu. Rev. Phytopathol. 1983, 21, 201–220. [Google Scholar] [CrossRef] [PubMed]
- Mamiya, Y. Pine wilting disease caused by the pine wood nematode, Bursaphelenchus lignicolus, in Japan. JARQ (Jpn. Agric. Res. Quart) 1976, 10, 206–211. [Google Scholar]
- Yang, B.J.; Wang, Q.L. Distribution of the pinewood nematode in China and susceptibility of some Chinese and exotic pines to the nematode. Can. J. For. Res. 1989, 19, 1527–1530. [Google Scholar]
- La, Y.J.; Moon, Y.S.; Yeo, W.H.; Shin, S.C.; Bak, W.C. Recent status of pine wilt disease in Korea. In Proceedings of the Sustainability of Pine Forests in Relation to Pine Wilt & Decline International Symposium, Tokyo, Japan, 27–28 October 1998. [Google Scholar]
- Sun, Y.C. First report of pine wilt disease from Pinus thunbergii Parl in Sun Yat-sen Mausoleum. J. Jiangsu For. Sci. Technol. 1982, 2, 27. [Google Scholar]
- Zhao, B.G. Pine wilt disease in China. In Pine Wilt Disease; Springer: Berlin/Heidelberg, Germany, 2008; pp. 18–25. [Google Scholar]
- Linit, M.J.; Kondo, E.; Smith, M.T. Insects associated with the pinewood nematode, Bursaphelenchus xylophilus (Nematoda: Aphelenchoididae), in Missouri. Environ. Entomol. 1983, 12, 467–470. [Google Scholar] [CrossRef]
- Linit, M.J. Nemtaode-vector relationships in the pine wilt disease system. J. Nematol. 1988, 20, 227. [Google Scholar]
- Linit, M.J. Transmission of Pinewood Nematode Through Feeding Wounds of Monochamus carolinensis (Coleoptera: Cerambycidae). J. Nematol. 1990, 22, 231–236. [Google Scholar]
- Bergdahl, D.; Halik, S.; Tomminen, J.; Akar, H. Frequency of infestation of Monochamus notatus and M. scutellatus by Bursaphelenchus xylophilus in Vermont. Phytopathology 1991, 81, 120. [Google Scholar]
- Luzzi, M.; Wilkinson, R.; Tarjan, A. Transmission of the pinewood nematode, Bursaphelenchus xylophilus, to slash pine trees and log bolts by a cerambycid beetle, Monochamus titillator, in Florida. J. Nematol. 1984, 16, 37. [Google Scholar]
- Akbulut, S.; Stamps, W. Insect vectors of the pinewood nematode: A review of the biology and ecology of Monochamus species. For. Pathol. 2012, 42, 89–99. [Google Scholar] [CrossRef]
- Wingfield, M.; Blanchette, R.A. The pine-wood nematode, Bursaphelenchus xylophilus, in Minnesota and Wisconsin: Insect associates and transmission studies. Can. J. For. Res. 1983, 13, 1068–1076. [Google Scholar] [CrossRef] [Green Version]
- Mamiya, Y.; Enda, N. Transmission of bursaphelenchus lignicolus (nematoda: Aphelenchoididae) by monochamus alternatus (coleoptera: Cerambycidae). Nematologica 1972, 18, 159–162. [Google Scholar] [CrossRef]
- Sato, H.; Takeshi, S.; Kobayashi, M. Transmission of Bursaphelenchus xylophilus (Steiner et Buhrer) Nickle (Nematoda, Aphelenchoididae) by Monochamus saltuarius (Gebler)(Coleoptera, Cerambycidae). J. Jpn. For. Soc. 1987, 69, 492–496. [Google Scholar]
- Haran, J.; Rousselet, J.; Tellez, D.; Roques, A.; Roux, G. Phylogeography of Monochamus galloprovincialis, the European vector of the pinewood nematode. J. Pest Sci. 2018, 91, 247–257. [Google Scholar] [CrossRef]
- Pajares, J.; Álvarez, G.; Hall, D.; Ibarra, N.; Hoch, G.; Halbig, P.; Cocoş, D.; Johansson, H.; Schroeder, M. Attractants for management of the pine sawyer beetle Monochamus sutor, a potential vector of Bursaphelenchus xylophilus. J. Appl. Entomol. 2017, 141, 97–111. [Google Scholar] [CrossRef]
- Togashi, K.; Taga, Y.; Iguchi, K.; Aikawa, T. Bursaphelenchus mucronatus (Nematoda: Aphelenchoididae) vectored by Monochamus urussovi (Coleoptera: Cerambycidae) in Hokkaido, Japan. J. For. Res. 2008, 13, 127–131. [Google Scholar] [CrossRef]
- Akbulut, S.; Keten, A.; Stamps, W.T. Population dynamics of Monochamus galloprovincialis Olivier (Coleoptera: Cerambycidae) in two pine species under laboratory conditions. J. Pest Sci. 2008, 81, 115–121. [Google Scholar] [CrossRef]
- Pajares, J.A.; Álvarez, G.; Ibeas, F.; Gallego, D.; Hall, D.R.; Farman, D.I. Identification and field activity of a male-produced aggregation pheromone in the pine sawyer beetle, Monochamus galloprovincialis. J. Chem. Ecol. 2010, 36, 570–583. [Google Scholar] [CrossRef]
- Li, Y.X. Analysis on the trend of invasion and expansion of Bursaphelenchus xylophilus. For. Pest Dis. 2018, 1, 1–4. [Google Scholar]
- Naves, A.C.G.P. Morphobiometric and Molecular Characterization of Bursaphelenchus Fuchs, 1937 (Nematoda: Aphelenchoididae) Species Associated with Pinus Pinaster Ainton in Portugal. Ph.D. Thesis, Universidade de Évora Portugal, Évora, Portugal, 2008. [Google Scholar]
- Koo, C.D.; Lee, H.Y.; Han, J.H.; Sung, J.H.; Shin, J.H. Infection behavior and distribution of Bursaphelenchus xylophilus in Pinus densiflora trees. For. Sci. Technol. 2013, 9, 81–86. [Google Scholar]
- Han, J.W.; Kim, H.K.; Kang, W.J.; Kim, G.A. Feeding and oviposition preference of the Sakhalin pine sawyer Monochamus saltuarius (Coleoptera: Cerambycidae) for various tree species. Entomol. Res. 2016, 46, 331–336. [Google Scholar] [CrossRef]
- Yu, W.Y.; Zhang, E.Y.; Zhi, Q. Monochamus saltuarius harms Pinus banksiana. Plant Prot. 1992, 18, 51. [Google Scholar]
- Han, H.; Chung, Y.J.; Shin, S.C. First report of pine wilt disease on Pinus koraiensis in Korea. Plant Dis. 2008, 92, 1251. [Google Scholar] [CrossRef] [PubMed]
- Nunes da Silva, M.; Lima, M.R.M.; Vasconcelos, M.W. Susceptibility evaluation of Picea abies and Cupressus lusitanica to the pine wood nematode (Bursaphelenchus xylophilus). Plant Pathol. 2013, 62, 1398–1406. [Google Scholar] [CrossRef]
- Shi, J.; Chen, F.; Luo, Y.Q.; Wang, Z.; Xie, B.Y. First isolation of pine wood nematode from Pinus tabuliformis forests in China. For. Pathol. 2013, 43, 59–66. [Google Scholar] [CrossRef]
- Nakamura, K.; Akiba, M.; Kanetani, S. Pine wilt disease as promising causal agent of the mass mortality of Pinus armandii Franch. var. amamiana (Koidz.) Hatusima in the field. Ecol. Res. 2001, 16, 795–801. [Google Scholar] [CrossRef]
- Zhang, X.; Stamps, W.T.; Linit, M.J. A nondestructive method of determining Bursaphelenchus xylophilus infestation of Monochamus spp. vectors. J. Nematol. 1995, 27, 36. [Google Scholar]
- Shuto, Y.; Watanabe, H. Attractants from a vector, Monochamus alternatus, for the pine wood nematode. Agric. Biol. Chem. 1987, 51, 1457–1458. [Google Scholar]
- Vrain, T.C.; Wakarchuk, D.A.; Levesque, A.C.; Hamilton, R.I. Intraspecific rDNA restriction fragment length polymorphism in the Xiphinema americanum group. Fundam. Appl. Nematol. 1992, 15, 563–573. [Google Scholar]
- Ferris, V.R. Variation in spacer ribosomal DNA in some cyst-forming species of plant parasitic nematodes. Fundam. Appl. Nematol. 1993, 16, 177–184. [Google Scholar]
- Chen, F.M.; Ye, J.R.; Tang, J.; Wu, X.Q. A Study on Detection Technique of Bursaphelenchus xylophilus and B. mucronatus by RAPD. J. Nanjing For. Univ. 2005, 29, 25–28. [Google Scholar]
- Green, M.; Rott, M.; Leal, I.; Humble, L.; Allen, E. Application of a real-time PCR method for the detection of pine wood nematode, Bursaphelenchus xylophilus, in wood samples from lodgepole pine. Nematology 2007, 9, 351–362. [Google Scholar] [CrossRef]
- Li, R.J.; Xu, F.Y.; Zhang, P. Host Preference by Monochamus alternatus (Hope) during Maturation Feeding on Pine Species and Masson Pine Provenances. Chin. For. Sci. Technol. 2003, 2, 91–98. [Google Scholar]
- Kubozono, M.; Sone, K.; Kawauchi, H.; Tsuji, M. Resistance to the pine wilt disease and initial invasion of the pine wood nematode, Bursaphelenchus xylophilus (Steiner et Buhrer) Nickle, into seedlings of resistant clones of Pinus thunbergii Parl. Bull. Kagoshima Univ. For. 1998, 26, 37–41. [Google Scholar]
- Ohsawa, M.; Akiba, M. Possible altitude and temperature limits on pine wilt disease: The reproduction of vector sawyer beetles (Monochamus alternatus), survival of causal nematode (Bursaphelenchus xylophilus), and occurrence of damage caused by the disease. Eur. J. For. Res. 2014, 133, 225–233. [Google Scholar]
- Zhang, K. Research Advances of Pine Wood Nematode Disease in China. World For. Res. 2010, 23, 59–63. [Google Scholar]
- Huang, J.; Zhang, J.; Li, M.J.; Xia, T.F. Seasonal variations in the incidence of Monochamus alternatus adults (Coleoptera: Cerambycidae) and other major Coleoptera: A two-year monitor in the pine forests of Hangzhou, Eastern China. Scand. J. For. Res. 2015, 30, 507–515. [Google Scholar] [CrossRef]
- Du, H.B.; Wu, Z.F.; Zhang, N.; Zong, S.W.; Meng, X.J. Characteristics of Extreme Temperature and Precipitation Events over Dandong During the Last Six Decades. Sci. Geogr. Sin. 2013, 33, 473–480. [Google Scholar]
- Zhao, L.L.; Wei, W.; Zhang, X.Y.; Kulhavy, D.; Sun, J.H. Low temperature induces two growth-arrested stages and change of secondary metabolites in Bursaphelenchus xylophilus. Nematology 2007, 9, 663–670. [Google Scholar] [CrossRef] [Green Version]
- Sheng, M.L. Two new species of the genus Ischnoceros Gravenhorst (Hymenoptera, Ichneumonidae) parasitizing woodborers with a key to species of Ischnoceros known in China. Acta Zootaxonomica Sin. 2008, 33, 508–513. [Google Scholar]
- Zhen, D.; Xu, F.Y.; Ge, M.H.; Zhu, K.G. Studies on the Relationship of Amino Acid Content of Different Masson Pine Provenances and Their Resistance to Pine Wood Nematode (PWN). For. Res. 1998, 3, 6–12. [Google Scholar]


{kind=link}
| Code of Beetles | First Detection of Nematodes on Twig (Days after Beetle Emergence) | Total Number of Nematodes Transmitted to Twig | Number of Nematodes Retained in Dead Beetle | Number of Nematodes Transmitted Per Day (Max-Min) | Last Detection of Nematodes (Days after Beetle Emergence) | Number of Days of Nematode Presence in the Feeding Wound during the Entire Feeding Period |
|---|---|---|---|---|---|---|
| 1 | 12 | 49 | 0 | 21–1 | 48 | 21 |
| 2 | 18 | 64 | 0 | 50–2 | 48 | 18 |
| 3 | 9 | 100 | 5 | 43–1 | 27 | 12 |
| 4 | 9 | 118 | 21,000 | 116–2 | 21 | 12 |
| 5 | 15 | 63 | 33 | 38–1 | 48 | 21 |
| 6 | 12 | 44 | 586 | 23–1 | 24 | 18 |
| 7 | 6 | 69 | 0 | 22–1 | 33 | 21 |
| 8 | 9 | 69 | 3450 | 27–1 | 36 | 9 |
| 9 | 15 | 550 | 0 | 550–550 | 9 | 12 |
| 10 | 15 | 20 | 0 | 9–1 | 24 | 27 |
| Treatment | No. | Date of Death | Isolation of B. xylophilus from: | |||||
|---|---|---|---|---|---|---|---|---|
| Needles | Upper Stems | Middle Stems | Lower Stems | Branches | Roots | |||
| Treated trees in the experimental cage (with Monochamus saltuarius) | 1 | August 31 | 573 ± 130 | 13,250 ± 2300 | 8760 ± 1750 | 8890 ± 1230 | 15,340 ± 4430 | 130 ± 75 |
| 2 | October 3 | 0 | 0 | 3200 ± 550 | 2250 ± 300 | 3595 ± 376 | 63 ± 10 | |
| 3 | August 31 | 330 ± 80 | 17,275 ± 5310 | 13,381 ± 3333 | 14,750 ± 2788 | 25,627 ± 8358 | 5112 ± 58 | |
| 4 | / | / | / | / | / | / | / | |
| 5 | October 26 | 119 ± 27 | 810 ± 120 | 573 ± 50 | 0 | 1139 ± 298 | 0 | |
| Treated trees in the experimental cage (without Monochamus saltuarius) | 1 | / | / | / | / | / | / | / |
| 2 | / | / | / | / | / | / | / | |
| 3 | / | / | / | / | / | / | / | |
| 4 | / | / | / | / | / | / | / | |
| 5 | / | / | / | / | / | / | / | |
| Tree Species | Bark Area Consumed/3 Days (mm2) | Number of Bursaphelenchus xylophilus Entering Twigs in 3 Days |
|---|---|---|
| Picea asperata * | (694 ± 7) a | (11 ± 1) a |
| Picea pungens * | (650 ± 12) a | (17 ± 3) a |
| Abies fabri * | (517 ± 15) a | (13 ± 5) a |
| Abies holophylla * | (444 ± 10) a | (11 ± 2) a |
| Juniperus formosana | (316 ± 7) b | (11 ± 3) a |
| Sabina chinensis | (308 ± 5) b | (16 ± 1) a |
| Larix principis | (335 ± 20) b | (11 ± 9) a |
| Larix olgensis | (314 ± 18) b | (17 ± 1) a |
| Podocarpus macrophyllus▲ | (205 ± 4) c | (13 ± 2) a |
| Taxus chinensis▲ | (139 ± 3) c | (9 ± 6) a |
| F | 736.6 | 1.421 |
| P | <0.0001 | 0.244 |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Li, M.; Li, H.; Sheng, R.-C.; Sun, H.; Sun, S.-H.; Chen, F.-M. The First Record of Monochamus saltuarius (Coleoptera; Cerambycidae) as Vector of Bursaphelenchus xylophilus and Its New Potential Hosts in China. Insects 2020, 11, 636. https://doi.org/10.3390/insects11090636
Li M, Li H, Sheng R-C, Sun H, Sun S-H, Chen F-M. The First Record of Monochamus saltuarius (Coleoptera; Cerambycidae) as Vector of Bursaphelenchus xylophilus and Its New Potential Hosts in China. Insects. 2020; 11(9):636. https://doi.org/10.3390/insects11090636
Chicago/Turabian StyleLi, Min, Huan Li, Ruo-Cheng Sheng, Hui Sun, Shou-Hui Sun, and Feng-Mao Chen. 2020. "The First Record of Monochamus saltuarius (Coleoptera; Cerambycidae) as Vector of Bursaphelenchus xylophilus and Its New Potential Hosts in China" Insects 11, no. 9: 636. https://doi.org/10.3390/insects11090636