Bacterial Communities in Three Parts of Intestinal Tracts of Carpenter Bees (Xylocopa tenuiscapa)

 and
and 
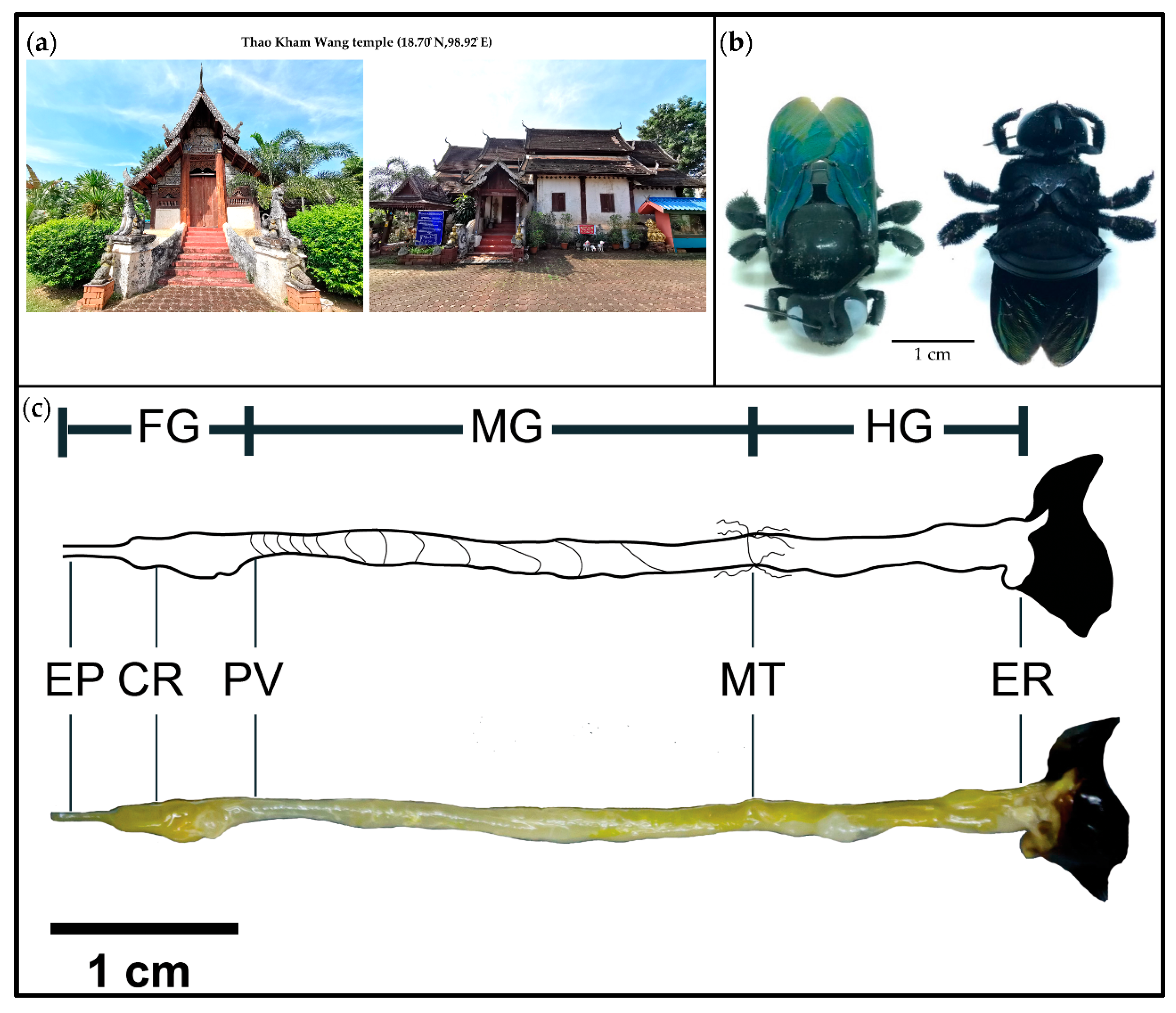{kind=link}
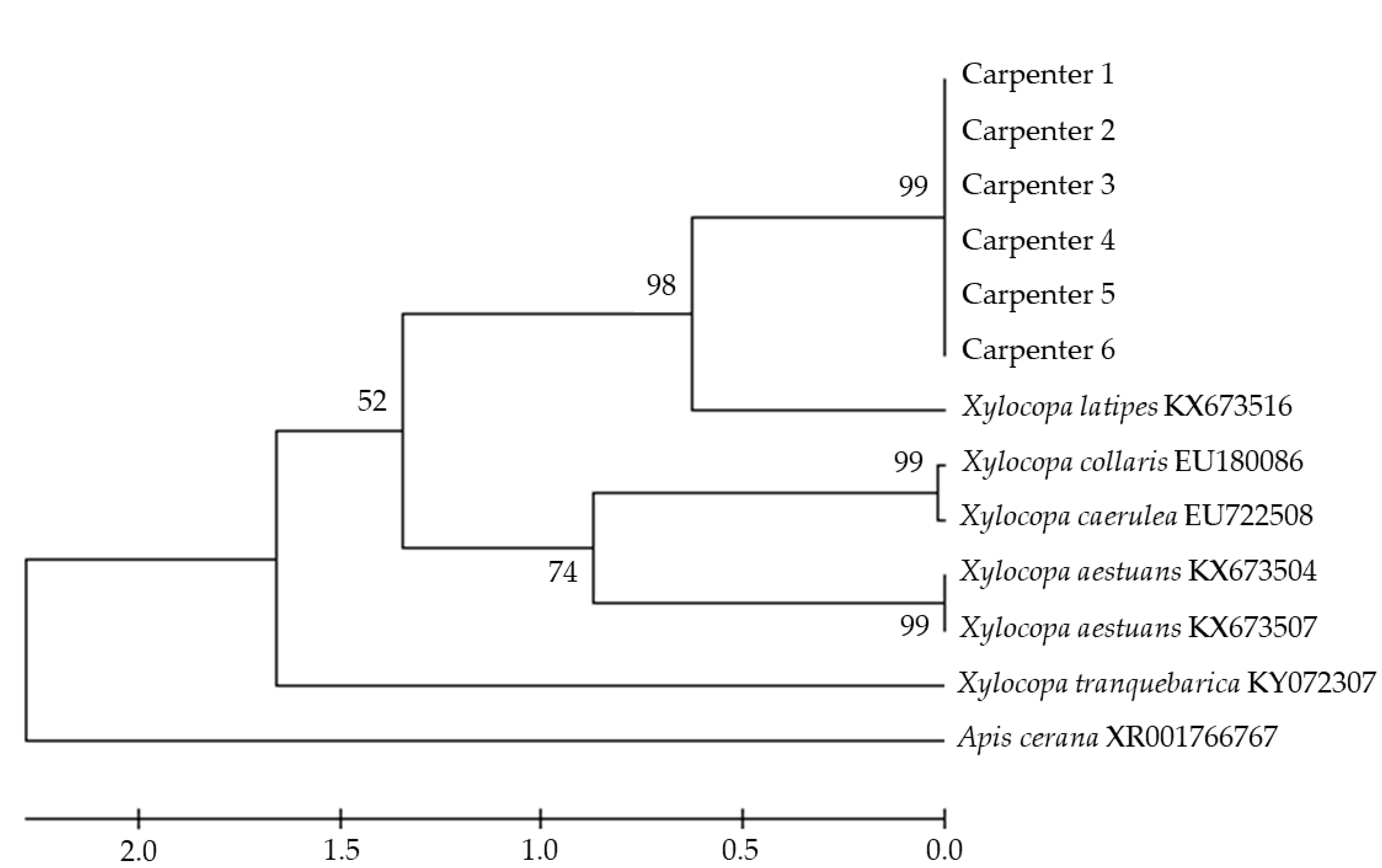{kind=link}
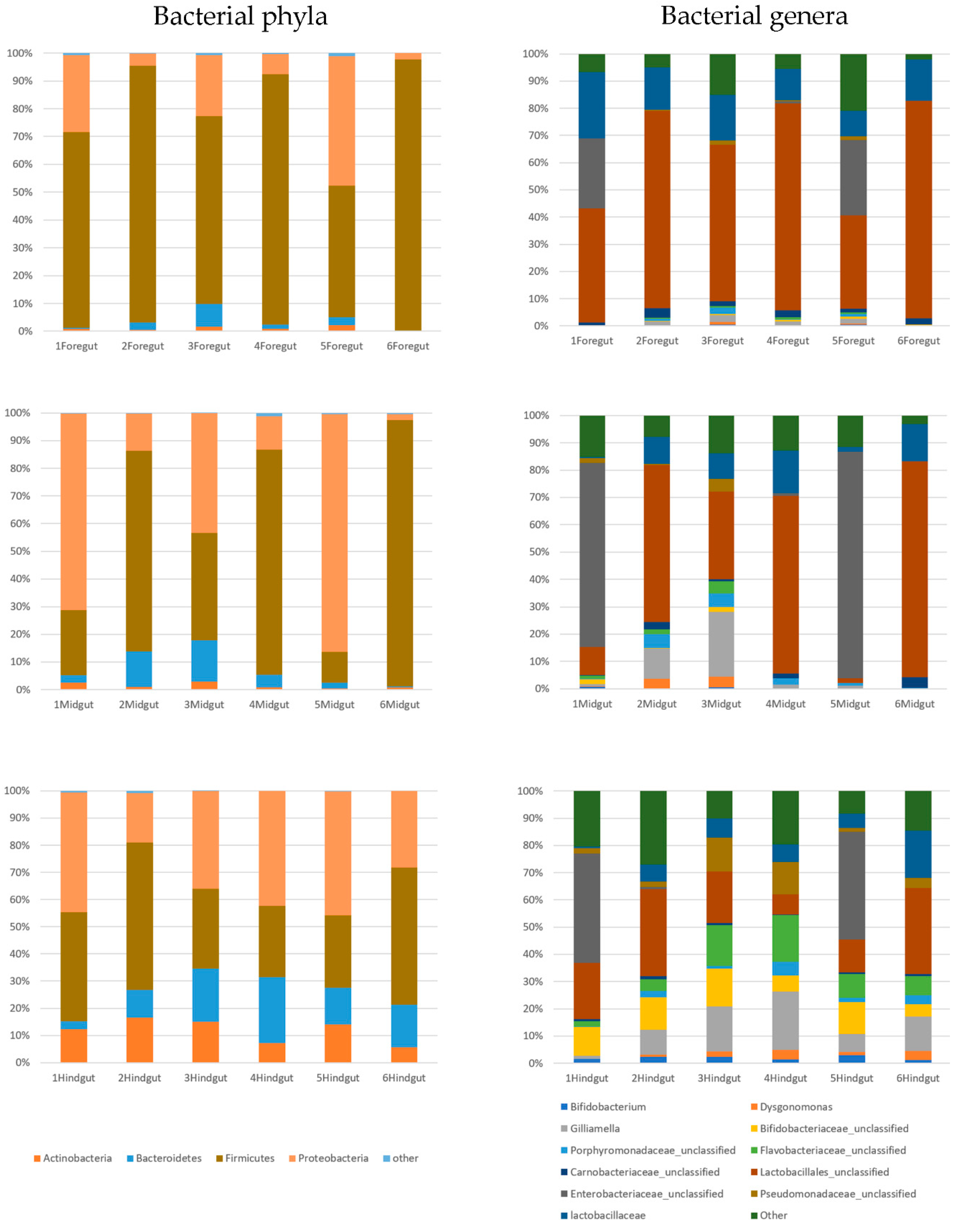{kind=link}
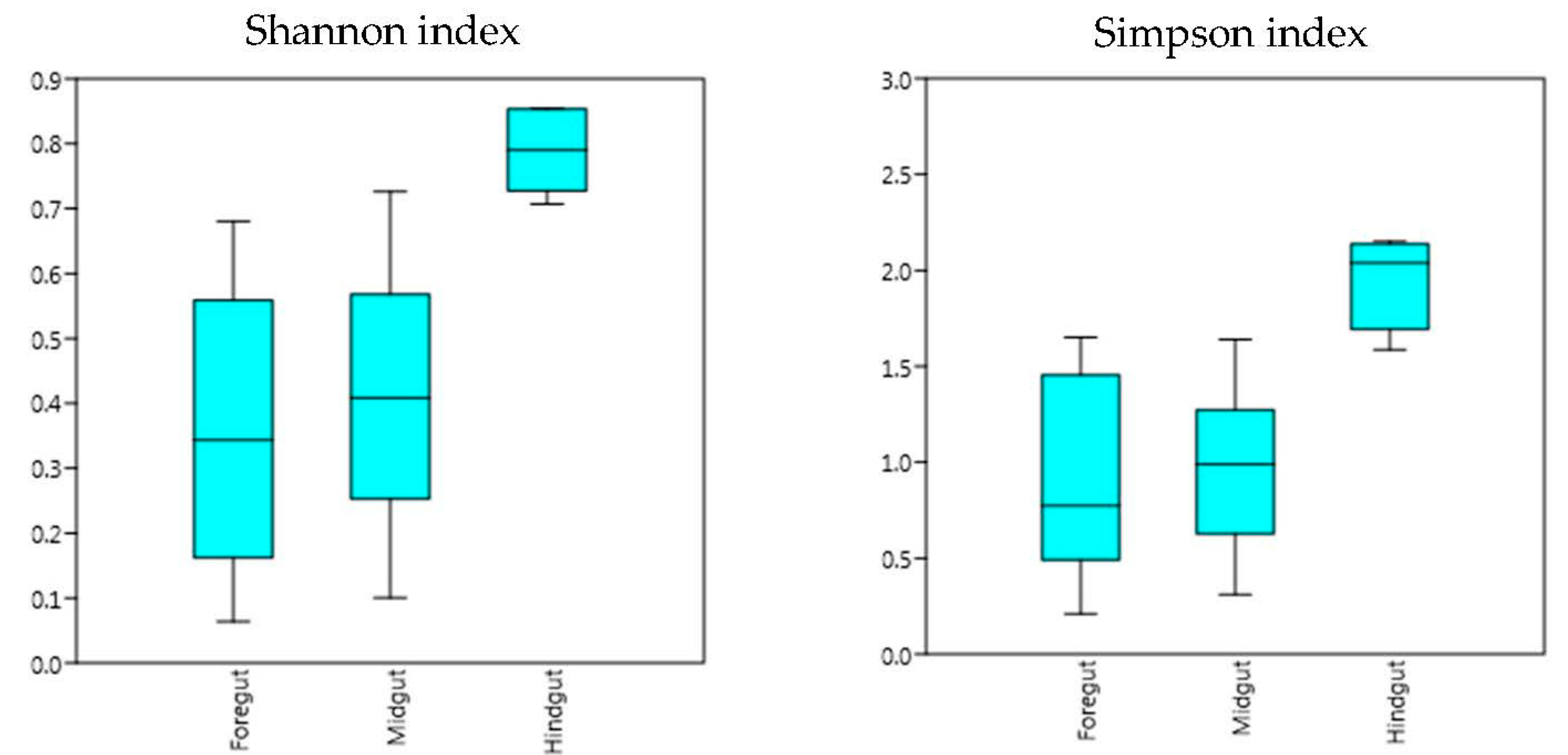{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Materials and Methods
2.1. Collection of Carpenter Bees
2.2. Classification of Carpenter Bee Samples
2.3. DNA Extraction and NGS of Gut Sections
2.3.1. Next Generation Sequencing
2.3.2. Data Analysis
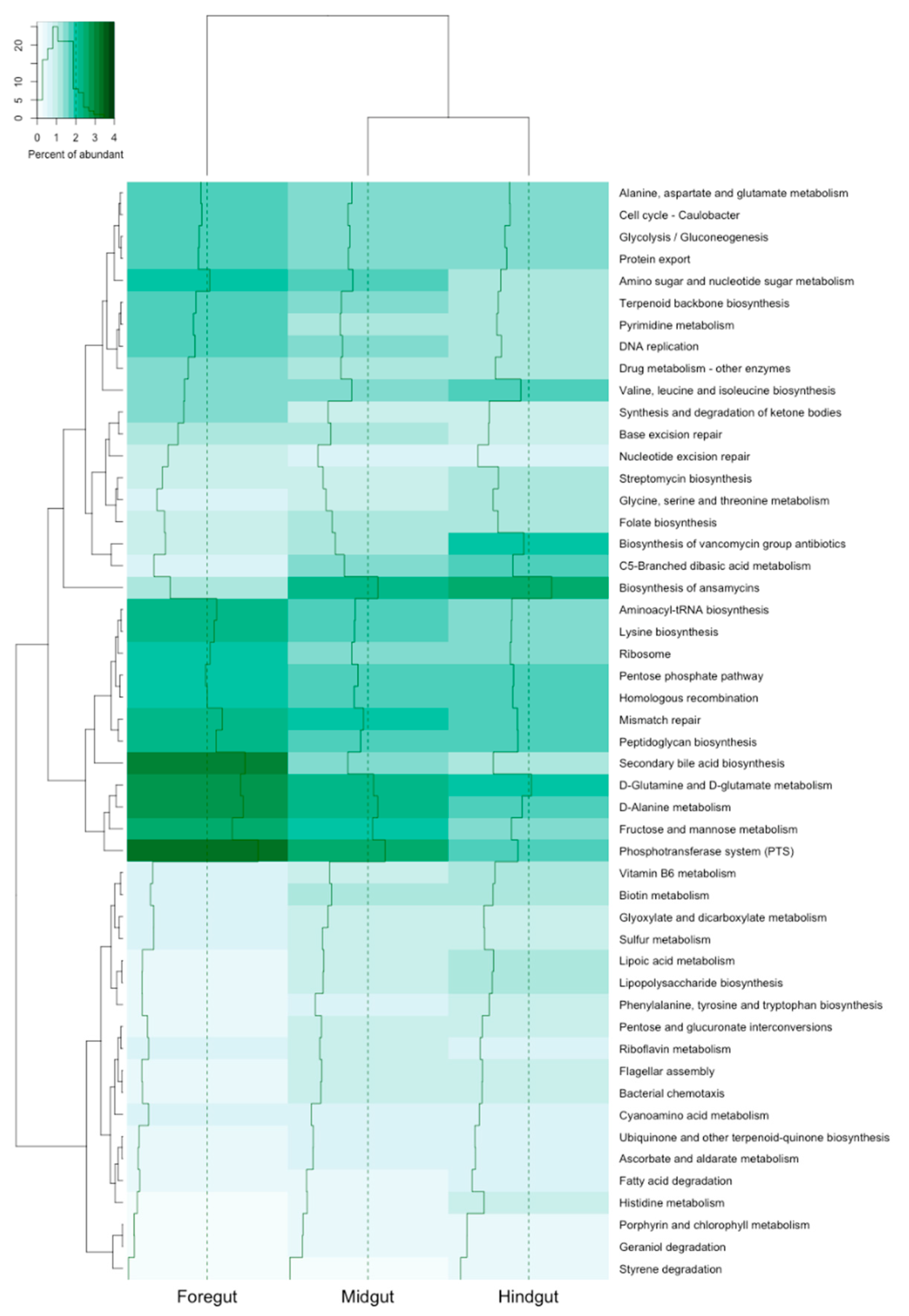
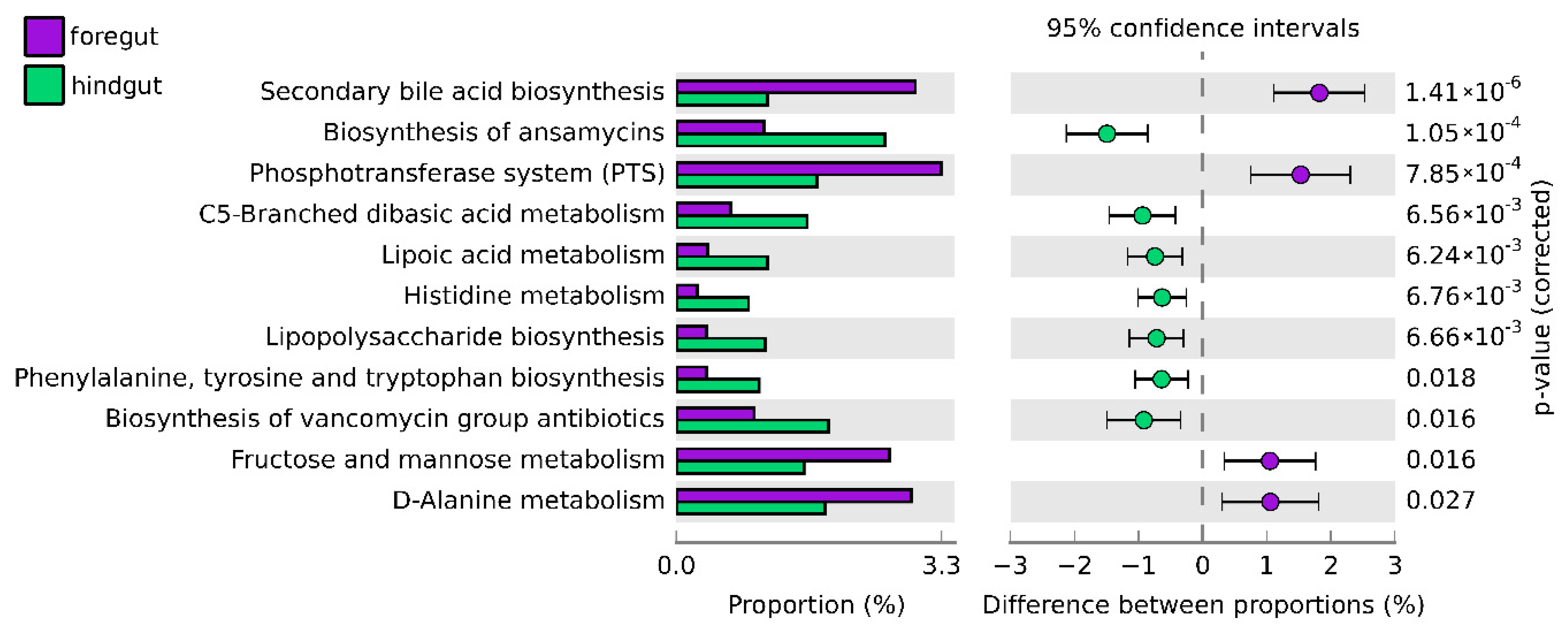
2.3.3. Prediction and Analysis of Functions of the Bacterial Microbiota
2.3.4. NGS Data Accession Number
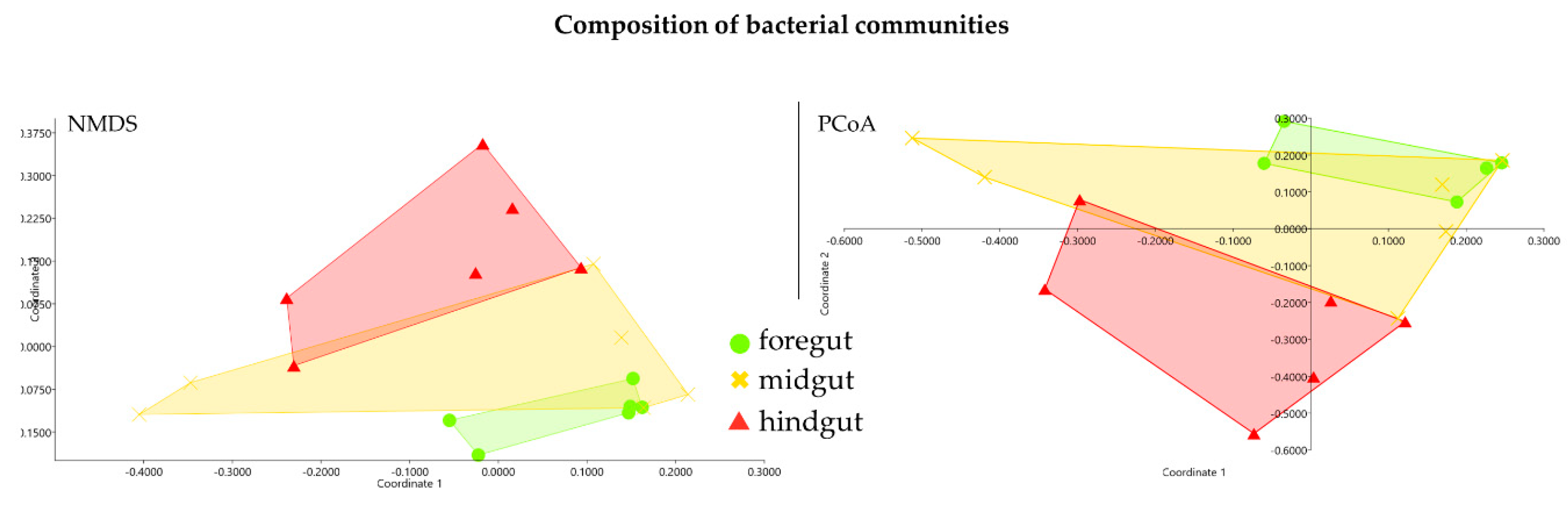
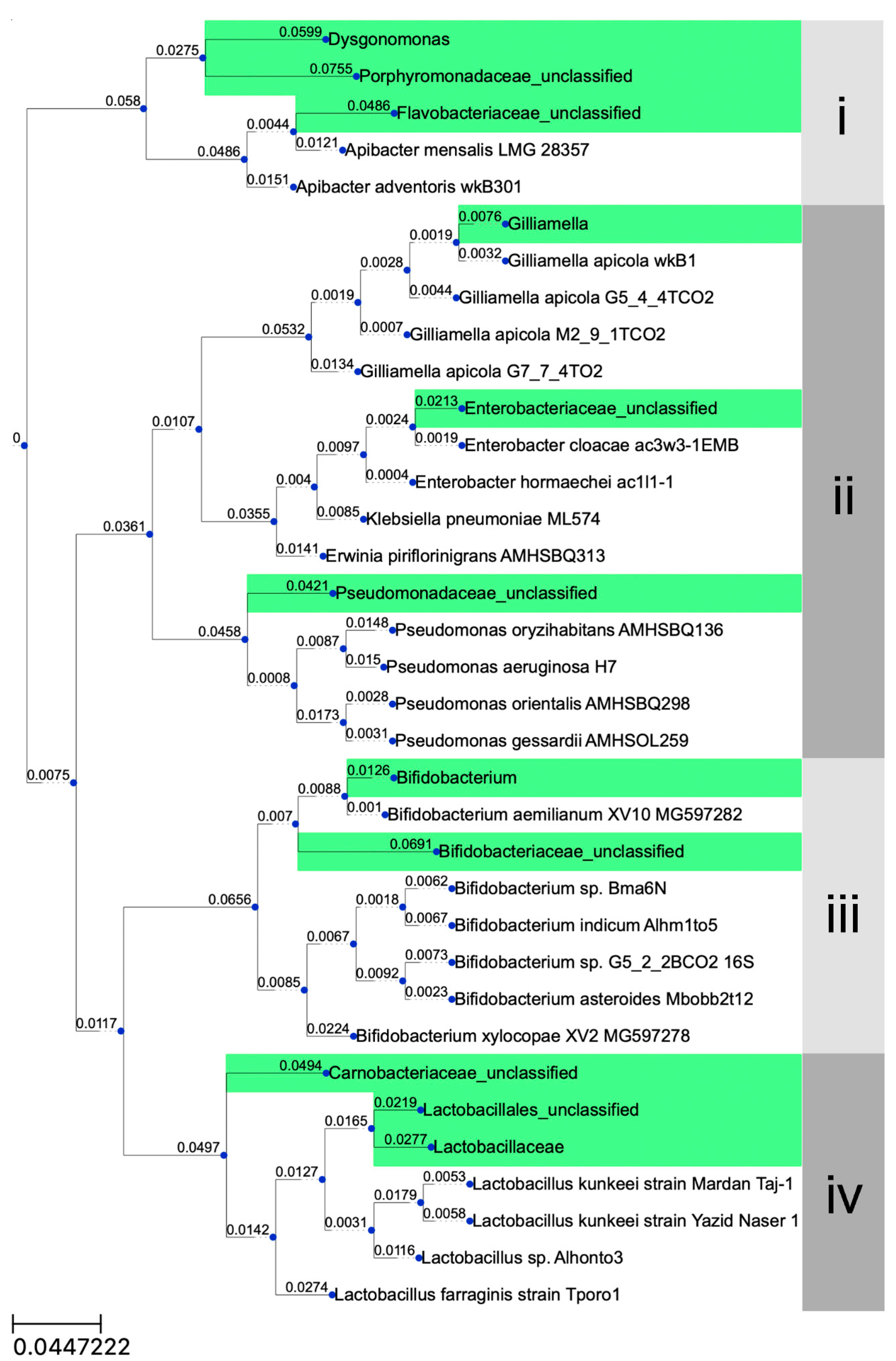
3. Results
3.1. Classification of Carpenter Bee Samples
3.2. Sequencing Results
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Conflicts of Interest
References
- Kingha, T.M.B.; Fohouo, F.N.T.; Ngakou, A.; Brueckner, D. Foraging and pollination activities of Xylocopa olivacea (Hymenoptera, Apidae) on Phaseolus vulgaris (Fabaceae) flowers at Dang (Ngaoundere-Cameroon). J. Agric. Ext. Rural. Dev. 2012, 4, 330–339. [Google Scholar]
- Azmi, W.A.; Ghazi, R.; Mohamed, N.Z. The importance of carpenter bee, Xylocopa varipuncta (Hymenoptera: Apidae as pollination agent for mangrove community of Setiu wetland, Terengganu. Sains Malays. 2012, 41, 1057–1062. [Google Scholar]
- Hongjamrassilp, W.; Warrit, N. Nesting biology of an Oriental carpenter bee, Xylocopa (Biluna) nasalis Westwood, 1838, in Thailand (Hymenoptera, Apidae, Xylocopinae). J. Hymenopt. Res. 2014, 41, 75–94. [Google Scholar]
- Stewart, A.B.; Sritongchuay, T.; Teartisup, P.; Kaewsomboon, S.; Bumrungsri, S. Habitat and landscape factors influence pollinators in a tropical megacity, Bangkok, Thailand. PeerJ 2018, 6, e5335. [Google Scholar] [CrossRef] [PubMed]
- Dillian, R.J.; Dillian, D.M. The Gut Bacteria of Insects: Nonpathogenic Interactions. Ann. Rev. Entomol. 2004, 49, 71–92. [Google Scholar] [CrossRef] [PubMed]
- Forsgren, E.; Olofsson, C.T.; Váasquez, A.; Fries, I. Novel lactic acid bacteria inhibiting Paenibacillus larvae in honey bee larvae. Apidologie 2010, 41, 91–108. [Google Scholar] [CrossRef] [Green Version]
- Olofsson, C.T.; Vasquez, A. Detection and identification of a novel lactic acid bacterial flora within the honey stomach of the honeybee Apis mellifera. Curr. Microbiol. 2008, 57, 356–363. [Google Scholar] [CrossRef]
- Promnuam, Y.; Kudo, T.; Chantawannakul, P. Actinomycetes isolated from beehives in Thailand. World J. Microbiol. Biotechnol. 2009, 25, 1685–1689. [Google Scholar] [CrossRef]
- Engel, P.; Martison, G.V.; Moran, A. Functional diversity within the simple gut microbiota of the honeybee. Proc. Natl. Acad. Sci. USA 2012, 109, 11002–11007. [Google Scholar] [CrossRef] [Green Version]
- Martinson, V.G.; Moy, J.; Moran, N.A. Establishment of characteristic gut bacteria during development of the honeybee worker. Appl. Environ. Microbiol. 2012, 78, 2830–2840. [Google Scholar] [CrossRef] [Green Version]
- Powell, J.E.; Martinson, V.G.; Urban-Mead, K.; Moran, N.A. Routes of acquisition of the gut microbiota of Apis mellifera. Appl. Environ. Microbiol. 2014, 80, 7378–7387. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Disayathanoowat, T.; Young, J.P.W.; Helgason, T.; Chantawannakul, P. T-RFLP analysis of bacterial communities in the midguts of Apis mellifera and Apis cerana honey bees in Thailand. FEMS Microbiol. Ecol. 2012, 79, 273–281. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Corby-Harris, V.; Maes, P.; Anderson, K.E. The bacterial communities associated with honey bee (Apis mellifera) Foragers. PLoS ONE 2014, 9, e95056. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Martinson, V.G.; Danforth, B.N.; Minckley, R.L.; Rueppell, O.; Tingek, S.; Moran, N.A. A simple and distinctive microbiota associated with honey bees and bumble bees. Mol. Ecol. 2011, 20, 619–628. [Google Scholar] [CrossRef]
- Hurd, P.D., Jr.; Moure, J.S. A classification on the large carpenter bees (Xylocopini) (Hymenoptera: Apoidea). Univ. Calif. Publ. Entomol. 1963, 29, 1–365. [Google Scholar]
- Pakwan, C.; Kaltenpoth, M.; Weiss, B.; Jun, G.; Disayathanoowat, T. Bacterial communities associated with the ectoparasitic mites Varroa destructor and Tropilaelaps mercedesae of the honey bee (Apis mellifera). FEMS Microbiol. Ecol. 2017, 94, fix160. [Google Scholar] [CrossRef] [PubMed]
- Kwong, W.K.; Moran, N.A. Gut microbial communities of social bees. Nat. Rev. Microbiol. 2016, 14, 374–384. [Google Scholar] [CrossRef]
- Chapman, R.F.; Simpson, S.J.; Douglas, A.E. The Insects: Structure and Function, 5th ed.; Cambridge University Press: Cambridge, UK, 2013; p. 959. [Google Scholar]
- Sheffield, C.S.; Hebert, P.D.N.; Kevan, P.G.; Packer, L. DNA barcoding a regional bee (Hymenoptera: Apoidea) fauna and its potential for ecological studies. Mol. Ecol. Resour. 2009, 9, 196–207. [Google Scholar] [CrossRef] [Green Version]
- Kumar, S.; Stecher, G.; Tamura, K. MEGA7: Molecular Evolutionary Genetics Analysis version 7.0 for bigger datasets. Mol. Biol. Evol. 2016, 33, 1870–1874. [Google Scholar] [CrossRef] [Green Version]
- Daffonchio, D.; Borin, S.; Consolandi, A.; Mora, D.; Manachini, P.L.; Sorlini, C. 16S–23S rRNA internal transcribed spacers as molecular markers for the species of the 16S rRNA group I of the genus Bacillus. FEMS Microbiol. Lett. 1998, 163, 229–236. [Google Scholar] [CrossRef]
- Schloss, P.D.; Westcott, S.L.; Ryabin, T.; Hall, J.R.; Hartmann, M.; Hollister, E.B.; Lesniewski, R.A.; Oakley, B.B.; Parks, D.H.; Robinson, C.J.; et al. Introducing mothur: Open-source, platform-independent, community-supported software for describing and comparing microbial communities. Appl. Environ. Microbiol. 2009, 75, 7537–7541. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Evans, J.D.S.; Schwarz, R.; Childers, A. HoloBee Database v2016.1; Ag Data Commons: Beltsville, MD, USA, 2016. [Google Scholar]
- Löytynoja, A.; Goldman, N. webPRANK: A phylogeny-aware multiple sequence aligner with interactive alignment browser. BMC Bioinform. 2010, 11, 579. [Google Scholar]
- Langille, M.G.; Zaneveld, J.; Caporaso, J.G.; McDonald, D.; Knights, D.; Reyes, J.A.; Clemente, J.C.; Burkepile, D.E.; Vega Thurber, R.L.; Knight, R.; et al. Predictive functional profiling of microbial communities using 16S rRNA marker gene sequences. Nat. Biotechnol. 2013, 31, 814–821. [Google Scholar] [CrossRef] [PubMed]
- Alberoni, D.; Gaggìa, F.; Baffoni, L.; Modesto, M.M.; Biavati, B.; Di Gioia, D. Bifidobacterium xylocopae sp. nov. and Bifidobacterium aemilianum sp. nov., from the carpenter bee (Xylocopa violacea) digestive tract. Syst. Appl. Microbiol. 2019, 42, 205–216. [Google Scholar] [CrossRef]
- Yun, J.H.; Roh, S.W.; Whon, T.W.; Jang, M.J.; Kim, M.S.; Kim, Y.J.; Shin, N.R.; Kim, S.H.; Lee, W.J.; Bae, J.W. Insect gut bacterial diversity determined by environmental, habitat, diet, developmental stage and physiology of host. J. Appl. Environ. Microbiol. 2014, 80, 5254–5264. [Google Scholar] [CrossRef] [Green Version]
- Brinkmann, N.; Martens, R.; Tebbe, C.C. Origin and diversity of metabolically active gut bacteria from laboratory-bred larvae of Manduca sexta (Sphingidae, Lepidoptera, Insecta). Environ. Microbiol. 2008, 78, 7189–7196. [Google Scholar] [CrossRef] [Green Version]
- Tikhe, C.V.; Sethi, A.; Delatte, J.; Husseneder, C. Isolation and assessment of gut bacteria from the Formosan subterranean termite, Coptotermes formosanus (Isoptera: Rhinotermitidae) for paratransgenesis research and application. Insect Sci. 2017, 24, 93–102. [Google Scholar] [CrossRef]
- Wang, W.W.; He, C.; Cui, J.; Wang, H.D.; Li, M.L. Comparative analysis of the composition of intestinal bacterial communities in Dastarcus helophoroides fed different diets. J. Insect Sci. 2014, 14, 111. [Google Scholar] [CrossRef]
- Broderick, N.A.; Raffa, K.F.; Goodman, R.M.; Handelsman, J. Census of the bacterial community of gypsy moth larval midgut by using culturing and culture independent methods. Appl. Environ. Microbiol. 2004, 70, 293–300. [Google Scholar] [CrossRef] [Green Version]
- Babendreier, D.; Joller, D.; Romeis, J.; Bigler, F.; Widmer, F. Bacterial community structures in honeybee intestines and their response to two insecticidal proteins. FEMS Microbiol. Ecol. 2007, 59, 600–610. [Google Scholar] [CrossRef]
- Carivenu, P.C.; Powell, J.E.; Koch, H.; Winfree, R.; Moran, N.A. Variation in gut microbial communities and its association with pathogen infection in wild bumble bees (Bombus). ISME J. 2014, 8, 2369–2379. [Google Scholar] [CrossRef]
- Lukjancenko, O.; Ussery, D.; Wassenaar, T.M. Comparitive genomics of Bifidobacterium, Lactobacillus and related probiotic genera. Microb. Ecol. 2012, 63, 651–673. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ouwehand, A.C.; Salminen, S.; Isolauri, E. Probiotics: An overview of beneficial effects. Anton. Leeuw. 2002, 82, 279–289. [Google Scholar] [CrossRef]
- Vásquez, A.; Olofsson, T.C.; Sammataro, D. A scientific note on the lactic acid bacterial flora in honeybees in the USA—A comparison with bees from Sweden. Apidologie 2009, 40, 26–28. [Google Scholar] [CrossRef] [Green Version]
- Killer, J.; Dubná, S.; Sedláček, I.; Svec, P. Lactobacillus apis sp. nov., from the stomach of honeybees (Apis mellifera), having an in vitro inhibitory effect on the causative agents of American and European foulbrood. Int. J. Syst. Evol. Microbiol. 2014, 64, 152–157. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bottacini, F.; Milani, C.; Turroni, F.; Sa’nchez, B.; Foroni, E.; Duranti, S.; Serafini, F.; Viappiani, A.; Strati, F.; Ferrarini, A.; et al. Bifidobacterium asteroides PRL2011 genome analysis reveals clues for colonization of the insect gut. PLoS ONE 2012, 7, e44229. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kwong, W.K.; Mancenido, A.L.; Moran, N.A. Genome sequences of Lactobacillus sp. strains wkB8 and wkB10, members of the Firm-5 clade, from honey bee guts. Genome Announc. 2014, 2, 1–2. [Google Scholar] [CrossRef] [Green Version]
- Ellegaard, K.M.; Tamarit, D.; Javelind, E. Extensive intra-phylotype diversity in Lactobacilli and Bifidobacteria from the honeybee gut. BMC Genom. 2015, 16, 284. [Google Scholar] [CrossRef] [Green Version]
- Butler, È.; Alsterfjord, M.; Olofsson, T.C.; Karlsson, C.; Malmström, J.; Vásquez, A. Proteins of novel lactic acid bacteria from Apis mellifera: An insight into the production of known extracellular proteins during microbial stress. BMC Microbiol. 2013, 13, 235. [Google Scholar] [CrossRef] [Green Version]
- Vasquez, A.; Forsgren, E.; Fries, I.; Paxton, R.J.; Flaberg, E. Symbionts as major modulators of insect health: Lactic acid bacteria and honeybees. PLoS ONE 2012, 7, e33188. [Google Scholar] [CrossRef]
- Evans, J.; Lopéz, D. Bacterial probiotics induce an immune response in the honey bee (Hymenoptera: Apidae). J. Econ. Entomol 2004, 97, 752–756. [Google Scholar] [CrossRef] [PubMed]
- Kwong, W.K.; Moran, N.A. Cultivation and characterization of the gut symbionts of honey bees and bumble bees: Description of Snodgrassella alvi gen. nov., sp. nov., a member of the family Neisseriaceae of the Betaproteobacteria, and Gilliamella apicola gen. nov., sp. nov., a member of Orbaceae fam. nov., Orbales ord. nov., a sister taxon to the order ‘Enterobacteriales’ of the Gammaproteobacteria. Int. J. Syst. Evol. Microbiol. 2013, 63, 2008–2018. [Google Scholar] [PubMed]
- Kwong, W.K.; Engel, P.; Koch, H.; Moran, N.A. Genomics and host specialization of honey bee and bumble bee gut symbionts. Proc. Natl. Acad. Sci. USA 2014, 111, 11509–11514. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Barker, R.J.; Lehner, Y. Acceptance and sustenance value of naturally occurring sugars fed to newly emerged adult workers of honey bees (Apis mellifera L.). J. Exp. Zool. 1974, 187, 277–285. [Google Scholar] [CrossRef]
- Ludvigsen, J.; Porcellato, D.; Amdam, G.V.; Rudi, K. Addressing the diversity of the honeybee gut symbiont Gilliamella: Description of Gilliamella apis sp. nov., isolated from the gut of honeybees (Apis mellifera). Int. J. Syst. Evol. Microbiol. 2018, 68, 1762–1770. [Google Scholar] [CrossRef]
- Rizzi, A.; Crotti, E.; Borruso, L.; Jucker, C.; Lupi, D.; Colombo, M.; Daffonchio, D. Characterization of the bacterial community associated with larvae and adults of Anoplophora chinensis collected in Italy by culture and culture-independent methods. BioMed Res. Intertional 2013, 2013, 420287. [Google Scholar]
- Xia, X.; Gurr, G.M.; Vasseur, L.; Zheng, D.; Zhong, H.; Qin, B.; Lin, J.; Wang, Y.; Song, F.; Li, Y.; et al. Metagenomic sequencing of diamondback moth gut microbiome unveils key holobiont adaptations for herbivory. Front. Microbial. 2017, 8, 663. [Google Scholar] [CrossRef]
- Kato, T.; Yamazaki, K.; Nakajima, M.; Date, Y.; Kikuchi, J.; Hase, K.; Ohno, H.; Yamazaki, K. Oral administration of Porphyromonas gingivalis alters the gut microbiome and serum metabolome. ASM J. 2018, 3. [Google Scholar] [CrossRef] [Green Version]
- Hadapad, A.B.; Shettigar, S.K.G.; Hire, R. Bacterial communities in the gut of wild and mass-reared Zeugodacus cucurbitae and Bactrocera dorsalis revealed by metagenomic sequencing. BMC Microbiol. 2019, 19, 1–11. [Google Scholar] [CrossRef]
- Praet, J.; Aerts, M.; Brandt, E.D.; Meeus, I.; Smagghe, G.; Vandamme, P. Apibacter mensalis sp. nov.: A rare member of the bumblebee gut microbiota. Int. J. Syst. Evol. Microbiol. 2016, 66, 1645–1651. [Google Scholar] [CrossRef]
- Kwong, W.K.; Moran, N.A. Apibacter adventoris gen. nov., sp. nov., a member of the phylum Bacteroidetes isolated from honey bees. Int. J. Syst. Evol. Microbiol. 2016, 66, 1323–1329. [Google Scholar] [CrossRef] [PubMed]
- Luo, C.; Li, Y.; Chen1, Y.; Fu, C.; Long, W.; Xiao, X.; Liao, H.; Yang, Y. Bamboo lignocellulose degradation by gut symbiotic microbiota of the bamboo snout beetle Cyrtotrachelus buqueti. Biotechnol. Biofuels 2019, 12, 70. [Google Scholar] [CrossRef] [PubMed]
- Akami, M.; Njintang, N.Y.; Gbaye, O.A.; Andongma, A.A.; Rashid, M.A.; Niu, C.Y.; Nukenine, E.N. Gut bacteria of the cowpea beetle mediate its resistance to dichlorvos and susceptibility to Lippia adoensis essential oil. Sci. Rep. 2019, 9, 1–13. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Jing, T.; Qi, F.; Wang, Z. Most dominant roles of insect gut bacteria: Digestion, detoxification, or essential nutrient provision? Microbiome 2020, 8, 1–20. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Engel, P.; Moran, N.A. The gut microbiota of insects–diversity in structure and function. FEMS Microbiol. Rev. 2013, 37, 699–735. [Google Scholar] [CrossRef] [PubMed]








© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Subta, P.; Yodsuwan, P.; Yongsawas, R.; In-on, A.; Warrit, N.; Panha, S.; Khongphinitbunjong, K.; Chantawannakul, P.; Attasopa, K.; Disayathanoowat, T. Bacterial Communities in Three Parts of Intestinal Tracts of Carpenter Bees (Xylocopa tenuiscapa). Insects 2020, 11, 497. https://doi.org/10.3390/insects11080497
Subta P, Yodsuwan P, Yongsawas R, In-on A, Warrit N, Panha S, Khongphinitbunjong K, Chantawannakul P, Attasopa K, Disayathanoowat T. Bacterial Communities in Three Parts of Intestinal Tracts of Carpenter Bees (Xylocopa tenuiscapa). Insects. 2020; 11(8):497. https://doi.org/10.3390/insects11080497
Chicago/Turabian StyleSubta, Phakamas, Phongsathon Yodsuwan, Rujipas Yongsawas, Ammarin In-on, Natapot Warrit, Somsak Panha, Kitiphong Khongphinitbunjong, Panuwan Chantawannakul, Korrawat Attasopa, and Terd Disayathanoowat. 2020. "Bacterial Communities in Three Parts of Intestinal Tracts of Carpenter Bees (Xylocopa tenuiscapa)" Insects 11, no. 8: 497. https://doi.org/10.3390/insects11080497