
Ligand-Gated Ion Channels: Prognostic and Therapeutic Implications for Gliomas
 , , , , , ,
, , , , , , 
Abstract
:1. Introduction
2. Cells Involved in Glioma Pathogenesis
2.1. Neurons
2.2. Microglia
2.3. Astrocytes
3. Role of Chemotaxis in the Spread of Gliotic Changes
Role of Proteases
4. Role of Ion Channels in Glioma Cell Signaling
5. Role of Genetics in Faulty Ligand Channel Activity
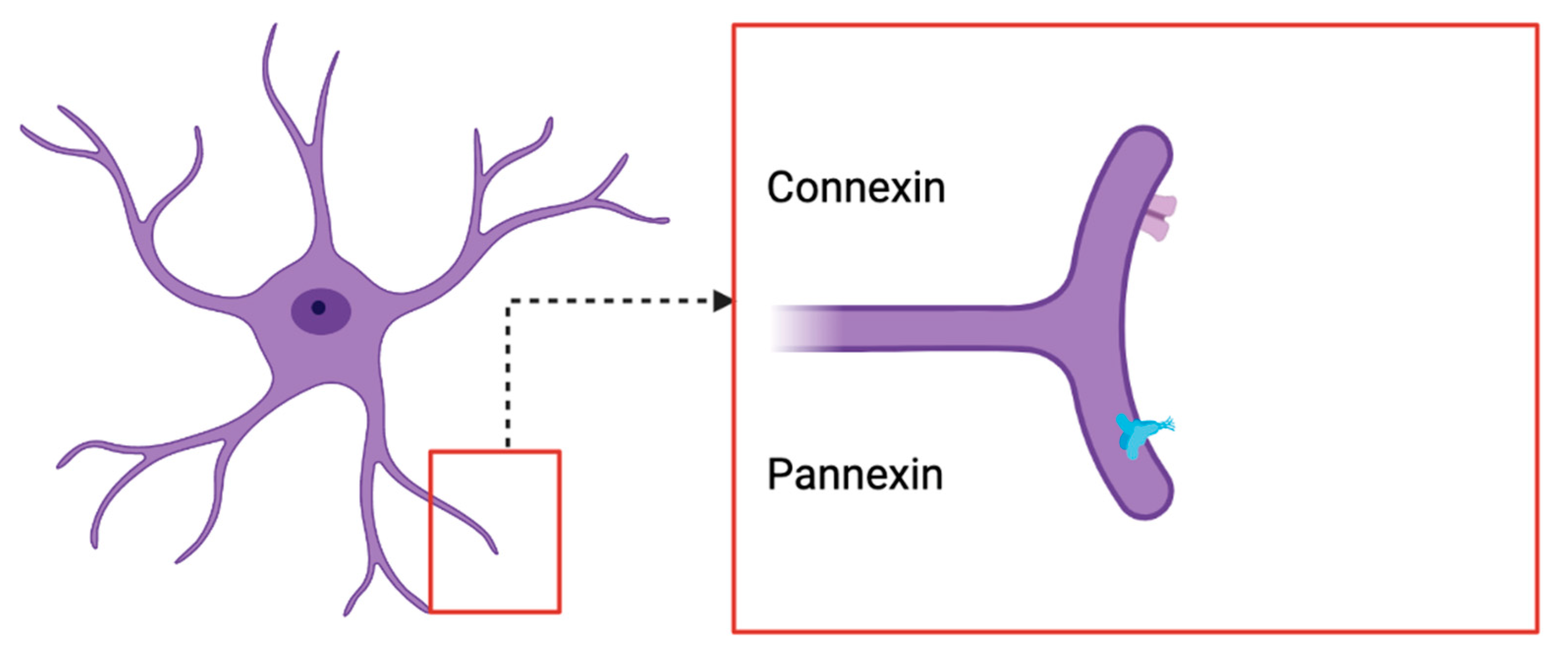
6. Overview of PANX2 Channels
7. LGICs as a Clinical Target
7.1. Purinoreceptors
7.2. Glutamate-Gated Receptors
7.3. Cys-Loop Receptors
8. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
Abbreviations
| 5-HT3 | Serotonin |
| ALS | Amyotrophic lateral sclerosis. |
| AMPA | α-amino-3-hydroxy-5-methyl-4-isoxazolepropionic acid. |
| ATP | Adenosine 5′-triphosphate. |
| BBB | Blood–brain barrier. |
| CLIC | Chloride intracellular channel proteins. |
| CNS | Central nervous system. |
| CaMK | calcium/calmodulin kinase. |
| ECD | Extracellular domain. |
| ECM | Extracellular matrix. |
| EGFR | Epidermal growth factor receptor. |
| ERG | Early response gene. |
| FAK | Focal adhesion kinase. |
| GBM | Glioblastoma. |
| GRE | Glioma-related epilepsy. |
| iCG | Ion Channel-based Gene. |
| LGIC | Ligand-gated ion channel. |
| MAPK | Mitogen-activated protein. |
| MMP-2 | matrix metalloproteinase 2. |
| NMDA | N-methyl-D-aspartate. |
| nACh | Acetylcholine. |
| Nf-kB | Nuclear factor kappa-B. |
| ROS | Reactive oxygen species. |
| SHH | Sonic hedgehog. |
| T2B | Topoisomerase-2-beta. |
| TAAs | Tumor-associated astrocytes. |
| TMD | Transmembrane domain. |
| TNF | Tumor necrosis factor. |
| TRAIL | Tumor necrosis factor-related apoptosis-inducing ligand. |
| TRPV1 | Vanilloid type-I. |
| VPA | Valproic acid. |
References
- Ohgaki, H.; Kleihues, P. Epidemiology and etiology of gliomas. Acta Neuropathol. 2005, 109, 93–108. [Google Scholar] [CrossRef]
- Louis, D.N.; Perry, A.; Wesseling, P.; Brat, D.J.; Cree, I.A.; Figarella-Branger, D.; Hawkins, C.; Ng, H.K.; Pfister, S.M.; Reifenberger, G.; et al. The 2021 WHO Classification of Tumors of the Central Nervous System: A summary. Neuro Oncol. 2021, 23, 1231–1251. [Google Scholar] [CrossRef] [PubMed]
- Liu, J.; Qu, C.; Han, C.; Chen, M.M.; An, L.J.; Zou, W. Potassium channels and their role in glioma: A mini review. Mol. Membr. Biol. 2019, 35, 76–85. [Google Scholar] [CrossRef]
- de Groot, J.; Sontheimer, H. Glutamate and the biology of gliomas. Glia 2011, 59, 1181–1189. [Google Scholar] [CrossRef]
- Wang, R.; Gurguis, C.I.; Gu, W.; Ko, E.A.; Lim, I.; Bang, H.; Zhou, T.; Ko, J.H. Ion channel gene expression predicts survival in glioma patients. Sci. Rep. 2015, 5, 11593. [Google Scholar] [CrossRef]
- Lastraioli, E.; Iorio, J.; Arcangeli, A. Ion channel expression as promising cancer biomarker. Biochim. Et Biophys. Acta (BBA)-Biomembr. 2015, 1848, 2685–2702. [Google Scholar] [CrossRef]
- Yi, B.A.; Jan, L.Y. Ion Channels. Encycl. Hum. Brain 2002, 599–615. [Google Scholar] [CrossRef]
- Alexander, S.P.H.; Mathie, A.; Peters, J.A. Ligand-Gated Ion Channels. Br. J. Pharmacol. 2011, 164, S115–S135. [Google Scholar] [CrossRef]
- Bylund, D.B. Receptor Types and Subtypes. Ref. Modul. Biomed. Sci. 2014. [Google Scholar] [CrossRef]
- Rao, R.; Shah, S.; Bhattacharya, D.; Toukam, D.K.; Caceres, R.; Pomeranz Krummel, D.A.; Sengupta, S. Ligand-Gated Ion Channels as Targets for Treatment and Management of Cancers. Front. Physiol. 2022, 13, 839437. [Google Scholar] [CrossRef]
- Ribeiro, M.P.C.; Custódio, J.B.A.; Santos, A.E. Ionotropic glutamate receptor antagonists and cancer therapy: Time to think out of the box? Cancer Chemother. Pharmacol. 2016, 79, 219–225. [Google Scholar] [CrossRef] [PubMed]
- Corsi, L.; Mescola, A.; Alessandrini, A. Glutamate Receptors and Glioblastoma Multiforme: An Old “Route” for New Perspectives. Int. J. Mol. Sci. 2019, 20, 1796. [Google Scholar] [CrossRef] [PubMed]
- Hansen, K.B.; Wollmuth, L.P.; Bowie, D.; Furukawa, H.; Menniti, F.S.; Sobolevsky, A.I.; Swanson, G.T.; Swanger, S.A.; Greger, I.H.; Nakagawa, T.; et al. Structure, Function, and Pharmacology of Glutamate Receptor Ion Channels. Pharm. Rev. 2021, 73, 298–487. [Google Scholar] [CrossRef]
- Kawate, T. P2X Receptor Activation. Adv. Exp. Med. Biol. 2017, 1051, 55–69. [Google Scholar] [CrossRef]
- Gendron, F.P.; Neary, J.T.; Theiss, P.M.; Sun, G.Y.; Gonzalez, F.A.; Weisman, G.A. Mechanisms of P2X7 receptor-mediated ERK1/2 phosphorylation in human astrocytoma cells. Am. J. Physiol. Cell Physiol. 2003, 284, C571–C581. [Google Scholar] [CrossRef]
- Guo, L.H.; Trautmann, K.; Schluesener, H.J. Expression of P2X4 receptor in rat C6 glioma by tumor-associated macrophages and activated microglia. J. Neuroimmunol. 2004, 152, 67–72. [Google Scholar] [CrossRef]
- Chen, S.; Zhang, S.; Feng, W.; Li, J.; Yuan, Y.; Li, W.; Wang, Z.; Yang, Y.; Liu, Y. Serine and glycine metabolism-related gene expression signature stratifies immune profiles of brain gliomas, and predicts prognosis and responses to immunotherapy. Front. Pharm. 2022, 13, 1072253. [Google Scholar] [CrossRef]
- He, F.; Yin, Z.; Wu, C.; Xia, Y.; Wu, M.; Li, P.; Zhang, H.; Yin, Y.; Li, N.; Zhu, G.; et al. l-Serine Lowers the Inflammatory Responses during Pasteurella multocida Infection. Infect. Immun. 2019, 87, e00677-19. [Google Scholar] [CrossRef]
- Hanahan, D.; Weinberg, R.A. Hallmarks of cancer: The next generation. Cell 2011, 144, 646–674. [Google Scholar] [CrossRef]
- Bhattacharya, D.; Gawali, V.S.; Kallay, L.; Toukam, D.K.; Koehler, A.; Stambrook, P.; Krummel, D.P.; Sengupta, S. Therapeutically leveraging GABAA receptors in cancer. Exp. Biol. Med. 2021, 246, 2128–2135. [Google Scholar] [CrossRef]
- Chen, J.; Wang, Z.; Wang, W.; Ren, S.; Xue, J.; Zhong, L.; Jiang, T.; Wei, H.; Zhang, C. SYT16 is a prognostic biomarker and correlated with immune infiltrates in glioma: A study based on TCGA data. Int. Immunopharmacol. 2020, 84, 106490. [Google Scholar] [CrossRef]
- Xu, X.; Hao, Y.; Xiong, S.; He, Z. PANX2 and brain lower grade glioma genesis: A bioinformatic analysis. Sci. Prog. 2021, 104, 368504211011836. [Google Scholar] [CrossRef]
- Piao, Y.; Lu, L.; de Groot, J. AMPA receptors promote perivascular glioma invasion via beta1 integrin-dependent adhesion to the extracellular matrix. Neuro Oncol. 2009, 11, 260–273. [Google Scholar] [CrossRef]
- Ye, Z.C.; Sontheimer, H. Glioma cells release excitotoxic concentrations of glutamate. Cancer Res. 1999, 59, 4383–4391. [Google Scholar]
- Ishiuchi, S.; Tsuzuki, K.; Yoshida, Y.; Yamada, N.; Hagimura, N.; Okado, H.; Miwa, A.; Kurihara, H.; Nakazato, Y.; Tamura, M.; et al. Blockage of Ca2+-permeable AMPA receptors suppresses migration and induces apoptosis in human glioblastoma cells. Nat. Med. 2002, 8, 971–978. [Google Scholar] [CrossRef]
- Giannone, G.; Ronde, P.; Gaire, M.; Beaudouin, J.; Haiech, J.; Ellenberg, J.; Takeda, K. Calcium rises locally trigger focal adhesion disassembly and enhance residency of focal adhesion kinase at focal adhesions. J. Biol. Chem. 2004, 279, 28715–28723. [Google Scholar] [CrossRef]
- Komuro, H.; Rakic, P. Modulation of neuronal migration by NMDA receptors. Science 1993, 260, 95–97. [Google Scholar] [CrossRef]
- van Vuurden, D.G.; Yazdani, M.; Bosma, I.; Broekhuizen, A.J.; Postma, T.J.; Heimans, J.J.; van der Valk, P.; Aronica, E.; Tannous, B.A.; Wurdinger, T.; et al. Attenuated AMPA receptor expression allows glioblastoma cell survival in glutamate-rich environment. PLoS ONE 2009, 4, e5953. [Google Scholar] [CrossRef]
- Puchalski, R.B.; Shah, N.; Miller, J.; Dalley, R.; Nomura, S.R.; Yoon, J.G.; Smith, K.A.; Lankerovich, M.; Bertagnolli, D.; Bickley, K.; et al. An anatomic transcriptional atlas of human glioblastoma. Science 2018, 360, 660–663. [Google Scholar] [CrossRef]
- Lutz, H.; Nguyen, T.A.; Joswig, J.; Rau, K.; Laube, B. NMDA Receptor Signaling Mediates cFos Expression via Top2beta-Induced DSBs in Glioblastoma Cells. Cancers 2019, 11, 306. [Google Scholar] [CrossRef]
- Laube, B.; Kuhse, J.; Betz, H. Evidence for a tetrameric structure of recombinant NMDA receptors. J. Neurosci. 1998, 18, 2954–2961. [Google Scholar] [CrossRef]
- Hardingham, G.E.; Bading, H. Synaptic versus extrasynaptic NMDA receptor signalling: Implications for neurodegenerative disorders. Nat. Rev. Neurosci. 2010, 11, 682–696. [Google Scholar] [CrossRef]
- Lau, C.G.; Zukin, R.S. NMDA receptor trafficking in synaptic plasticity and neuropsychiatric disorders. Nat. Rev. Neurosci. 2007, 8, 413–426. [Google Scholar] [CrossRef]
- Madabhushi, R.; Gao, F.; Pfenning, A.R.; Pan, L.; Yamakawa, S.; Seo, J.; Rueda, R.; Phan, T.X.; Yamakawa, H.; Pao, P.C.; et al. Activity-Induced DNA Breaks Govern the Expression of Neuronal Early-Response Genes. Cell 2015, 161, 1592–1605. [Google Scholar] [CrossRef]
- Li, L.; Hanahan, D. Hijacking the neuronal NMDAR signaling circuit to promote tumor growth and invasion. Cell 2013, 153, 86–100. [Google Scholar] [CrossRef]
- Liu, Z.G.; Jiang, G.; Tang, J.; Wang, H.; Feng, G.; Chen, F.; Tu, Z.; Liu, G.; Zhao, Y.; Peng, M.J.; et al. c-Fos over-expression promotes radioresistance and predicts poor prognosis in malignant glioma. Oncotarget 2016, 7, 65946–65956. [Google Scholar] [CrossRef]
- Kenig, S.; Faoro, V.; Bourkoula, E.; Podergajs, N.; Ius, T.; Vindigni, M.; Skrap, M.; Lah, T.; Cesselli, D.; Storici, P.; et al. Topoisomerase IIbeta mediates the resistance of glioblastoma stem cells to replication stress-inducing drugs. Cancer Cell Int. 2016, 16, 58. [Google Scholar] [CrossRef]
- Stebbing, M.J.; Cottee, J.M.; Rana, I. The Role of Ion Channels in Microglial Activation and Proliferation-A Complex Interplay between Ligand-Gated Ion Channels, K+ Channels, and Intracellular Ca2+. Front. Immunol. 2015, 6, 497. [Google Scholar] [CrossRef]
- Young, C.N.J.; Chira, N.; Róg, J.; Al-Khalidi, R.; Benard, M.; Galas, L.; Chan, P.; Vaudry, D.; Zabłocki, K.; Górecki, D.C. Sustained activation of P2X7 induces MMP-2-evoked cleavage and functional purinoceptor inhibition. J. Mol. Cell Biol. 2018, 10, 229–242. [Google Scholar] [CrossRef]
- Drill, M.; Powell, K.L.; Kan, L.K.; Jones, N.C.; O’Brien, T.J.; Hamilton, J.A.; Monif, M. Inhibition of purinergic P2X receptor 7 (P2X7R) decreases granulocyte-macrophage colony-stimulating factor (GM-CSF) expression in U251 glioblastoma cells. Sci. Rep. 2020, 10, 14844. [Google Scholar] [CrossRef]
- Bergamin, L.S.; Capece, M.; Salaro, E.; Sarti, A.C.; Falzoni, S.; Pereira, M.S.L.; De Bastiani, M.A.; Scholl, J.N.; Battastini, A.M.O.; Di Virgilio, F. Role of the P2X7 receptor in in vitro and in vivo glioma tumor growth. Oncotarget 2019, 10, 4840–4856. [Google Scholar] [CrossRef]
- Adinolfi, E.; Callegari, M.G.; Ferrari, D.; Bolognesi, C.; Minelli, M.; Wieckowski, M.R.; Pinton, P.; Rizzuto, R.; Di Virgilio, F. Basal Activation of the P2X7 ATP Receptor Elevates Mitochondrial Calcium and Potential, Increases Cellular ATP Levels, and Promotes Serum-independent Growth. Mol. Biol. Cell 2005, 16, 3260–3272. [Google Scholar] [CrossRef]
- Mai, Y.; Guo, Z.; Yin, W.; Zhong, N.; Dicpinigaitis, P.V.; Chen, R. P2X Receptors: Potential Therapeutic Targets for Symptoms Associated With Lung Cancer—A Mini Review. Front. Oncol. 2021, 11, 691956. [Google Scholar] [CrossRef]
- Giovannoni, F.; Quintana, F.J. The Role of Astrocytes in CNS Inflammation. Trends Immunol. 2020, 41, 805–819. [Google Scholar] [CrossRef]
- Nieland, L.; Morsett, L.M.; Broekman, M.L.D.; Breakefield, X.O.; Abels, E.R. Extracellular Vesicle-Mediated Bilateral Communication between Glioblastoma and Astrocytes. Trends Neurosci. 2021, 44, 215–226. [Google Scholar] [CrossRef]
- Tabatabaee, M.; Menard, F. Glutamate Signaling and Filopodiagenesis of Astrocytoma Cells in Brain Cancers: Survey and Questions. Cells 2022, 11, 2657. [Google Scholar] [CrossRef]
- Lim, D.; Semyanov, A.; Genazzani, A.; Verkhratsky, A. Calcium signaling in neuroglia. Inter-Organellar Ca2+ Signaling in Health and Disease-Part A. Int. Rev. Cell Mol. Biol. 2021, 362, 1–53. [Google Scholar] [CrossRef]
- Sofroniew, M.V. Astrogliosis. Cold Spring Harb. Perspect. Biol. 2015, 7, a020420. [Google Scholar] [CrossRef]
- Brandao, M.; Simon, T.; Critchley, G.; Giamas, G. Astrocytes, the rising stars of the glioblastoma microenvironment. Glia 2018, 67, 779–790. [Google Scholar] [CrossRef]
- Kim, J.-K.; Jin, X.; Sohn, Y.-W.; Jin, X.; Jeon, H.-Y.; Kim, E.-J.; Ham, S.W.; Jeon, H.-M.; Chang, S.-Y.; Oh, S.-Y.; et al. Tumoral RANKL activates astrocytes that promote glioma cell invasion through cytokine signaling. Cancer Lett. 2014, 353, 194–200. [Google Scholar] [CrossRef]
- Becher, O.J.; Hambardzumyan, D.; Fomchenko, E.I.; Momota, H.; Mainwaring, L.; Bleau, A.-M.; Katz, A.M.; Edgar, M.; Kenney, A.M.; Cordon-Cardo, C.; et al. Gli Activity Correlates with Tumor Grade in Platelet-Derived Growth Factor–Induced Gliomas. Cancer Res. 2008, 68, 2241–2249. [Google Scholar] [CrossRef]
- Guan, X.; Hasan, M.N.; Maniar, S.; Jia, W.; Sun, D. Reactive Astrocytes in Glioblastoma Multiforme. Mol. Neurobiol. 2018, 55, 6927–6938. [Google Scholar] [CrossRef]
- Sforna, L.; Cenciarini, M.; Belia, S.; D’Adamo, M.C.; Pessia, M.; Franciolini, F.; Catacuzzeno, L. The role of ion channels in the hypoxia-induced aggressiveness of glioblastoma. Front. Cell. Neurosci. 2015, 8, 467. [Google Scholar] [CrossRef] [PubMed]
- Bazargani, N.; Attwell, D. Astrocyte calcium signaling: The third wave. Nat. Neurosci. 2016, 19, 182–189. [Google Scholar] [CrossRef] [PubMed]
- Olsen, M.L.; Schade, S.; Lyons, S.A.; Amaral, M.D.; Sontheimer, H. Expression of Voltage-Gated Chloride Channels in Human Glioma Cells. J. Neurosci. 2003, 23, 5572–5582. [Google Scholar] [CrossRef]
- Setti, M.; Savalli, N.; Osti, D.; Richichi, C.; Angelini, M.; Brescia, P.; Fornasari, L.; Carro, M.S.; Mazzanti, M.; Pelicci, G. Functional Role of CLIC1 Ion Channel in Glioblastoma-Derived Stem/Progenitor Cells. JNCI J. Natl. Cancer Inst. 2013, 105, 1644–1655. [Google Scholar] [CrossRef] [PubMed]
- Roussos, E.T.; Condeelis, J.S.; Patsialou, A. Chemotaxis in cancer. Nat. Rev. Cancer 2011, 11, 573–587. [Google Scholar] [CrossRef] [PubMed]
- Annesley, S.J.; Fisher, P.R. Dictyostelium discoideum—A model for many reasons. Mol. Cell. Biochem. 2009, 329, 73–91. [Google Scholar] [CrossRef] [PubMed]
- Joyce, J.A.; Pollard, J.W. Microenvironmental regulation of metastasis. Nat. Rev. Cancer 2008, 9, 239–252. [Google Scholar] [CrossRef]
- Karve, I.P.; Taylor, J.M.; Crack, P.J. The contribution of astrocytes and microglia to traumatic brain injury. Br. J. Pharmacol. 2016, 173, 692–702. [Google Scholar] [CrossRef]
- Jha, M.K.; Jeon, S.; Suk, K. Glia as a Link between Neuroinflammation and Neuropathic Pain. Immune Netw. 2012, 12, 41–47. [Google Scholar] [CrossRef] [PubMed]
- Park, K.C.; Dharmasivam, M.; Richardson, D.R. The Role of Extracellular Proteases in Tumor Progression and the Development of Innovative Metal Ion Chelators That Inhibit Their Activity. Int. J. Mol. Sci. 2020, 21, 6805. [Google Scholar] [CrossRef] [PubMed]
- Gialeli, C.; Theocharis, A.D.; Karamanos, N.K. Roles of matrix metalloproteinases in cancer progression and their pharmacological targeting. FEBS J. 2011, 278, 16–27. [Google Scholar] [CrossRef] [PubMed]
- Mohamed, M.M.; Sloane, B.F. Multifunctional enzymes in cancer. Nat. Rev. Cancer 2006, 6, 764–775. [Google Scholar] [CrossRef]
- DeClerck, Y.A.; Mercurio, A.M.; Stack, M.S.; Chapman, H.A.; Zutter, M.M.; Muschel, R.J.; Raz, A.; Matrisian, L.M.; Sloane, B.F.; Noel, A.; et al. Proteases, Extracellular Matrix, and Cancer. Am. J. Pathol. 2004, 164, 1131–1139. [Google Scholar] [CrossRef]
- Breznik, B.; Motaln, H.; Lah Turnšek, T. Proteases and cytokines as mediators of interactions between cancer and stromal cells in tumours. Biol. Chem. 2017, 398, 709–719. [Google Scholar] [CrossRef]
- López-Otín, C.; Matrisian, L.M. Emerging roles of proteases in tumour suppression. Nat. Rev. Cancer 2007, 7, 800–808. [Google Scholar] [CrossRef]
- Egeblad, M.; Werb, Z. New functions for the matrix metalloproteinases in cancer progression. Nat. Rev. Cancer 2002, 2, 161–174. [Google Scholar] [CrossRef]
- Gocheva, V.; Zeng, W.; Ke, D.; Klimstra, D.; Reinheckel, T.; Peters, C.; Hanahan, D.; Joyce, J.A. Distinct roles for cysteine cathepsin genes in multistage tumorigenesis. Genes Dev. 2006, 20, 543–556. [Google Scholar] [CrossRef]
- Venkataramani, V.; Tanev, D.I.; Strahle, C.; Studier-Fischer, A.; Fankhauser, L.; Kessler, T.; Körber, C.; Kardorff, M.; Ratliff, M.; Xie, R.; et al. Glutamatergic synaptic input to glioma cells drives brain tumour progression. Nature 2019, 573, 532–538. [Google Scholar] [CrossRef]
- Venkataramani, V.; Yang, Y.; Schubert, M.C.; Reyhan, E.; Tetzlaff, S.K.; Wißmann, N.; Botz, M.; Soyka, S.J.; Beretta, C.A.; Pramatarov, R.L.; et al. Glioblastoma hijacks neuronal mechanisms for brain invasion. Cell 2022, 185, 2899–2917.e2831. [Google Scholar] [CrossRef]
- Tsuchihashi, K.; Okazaki, S.; Ohmura, M.; Ishikawa, M.; Sampetrean, O.; Onishi, N.; Wakimoto, H.; Yoshikawa, M.; Seishima, R.; Iwasaki, Y.; et al. The EGF Receptor Promotes the Malignant Potential of Glioma by Regulating Amino Acid Transport System xc(—). Cancer Res. 2016, 76, 2954–2963. [Google Scholar] [CrossRef] [PubMed]
- Suina, K.; Tsuchihashi, K.; Yamasaki, J.; Kamenori, S.; Shintani, S.; Hirata, Y.; Okazaki, S.; Sampetrean, O.; Baba, E.; Akashi, K.; et al. Epidermal growth factor receptor promotes glioma progression by regulating xCT and GluN2B-containing N-methyl-d-aspartate-sensitive glutamate receptor signaling. Cancer Sci. 2018, 109, 3874–3882. [Google Scholar] [CrossRef]
- Griffin, M.; Khan, R.; Basu, S.; Smith, S. Ion Channels as Therapeutic Targets in High Grade Gliomas. Cancers 2020, 12, 3068. [Google Scholar] [CrossRef]
- Bi, W.L.; Beroukhim, R. Beating the odds: Extreme long-term survival with glioblastoma. Neuro Oncol. 2014, 16, 1159–1160. [Google Scholar] [CrossRef]
- Krex, D.; Klink, B.; Hartmann, C.; von Deimling, A.; Pietsch, T.; Simon, M.; Sabel, M.; Steinbach, J.P.; Heese, O.; Reifenberger, G.; et al. Long-term survival with glioblastoma multiforme. Brain 2007, 130, 2596–2606. [Google Scholar] [CrossRef]
- Palmieri, C.; Rudraraju, B.; Monteverde, M.; Lattanzio, L.; Gojis, O.; Brizio, R.; Garrone, O.; Merlano, M.; Syed, N.; Lo Nigro, C.; et al. Methylation of the calcium channel regulatory subunit alpha2delta-3 (CACNA2D3) predicts site-specific relapse in oestrogen receptor-positive primary breast carcinomas. Br. J. Cancer 2012, 107, 375–381. [Google Scholar] [CrossRef] [PubMed]
- Venet, D.; Dumont, J.E.; Detours, V. Most random gene expression signatures are significantly associated with breast cancer outcome. PLoS Comput. Biol. 2011, 7, e1002240. [Google Scholar] [CrossRef] [PubMed]
- Saotome, K.; Singh, A.K.; Sobolevsky, A.I. Determining the Crystal Structure of TRPV6. In Calcium Entry Channels in Non-Excitable Cells; Kozak, J.A., Putney, J.W., Jr., Eds.; CRC Press/Taylor & Francis: Boca Raton, FL, USA, 2018; pp. 275–292. [Google Scholar] [CrossRef]
- Wang, H.Y.; Li, J.Y.; Liu, X.; Yan, X.Y.; Wang, W.; Wu, F.; Liang, T.Y.; Yang, F.; Hu, H.M.; Mao, H.X.; et al. A three ion channel genes-based signature predicts prognosis of primary glioblastoma patients and reveals a chemotherapy sensitive subtype. Oncotarget 2016, 7, 74895–74903. [Google Scholar] [CrossRef] [PubMed]
- Hervey-Jumper, S.L.; Berger, M.S. Maximizing safe resection of low- and high-grade glioma. J. Neurooncol. 2016, 130, 269–282. [Google Scholar] [CrossRef] [PubMed]
- Takayasu, T.; Kurisu, K.; Esquenazi, Y.; Ballester, L.Y. Ion Channels and Their Role in the Pathophysiology of Gliomas. Mol. Cancer 2020, 19, 1959–1969. [Google Scholar] [CrossRef] [PubMed]
- McKinnon, C.; Nandhabalan, M.; Murray, S.A.; Plaha, P. Glioblastoma: Clinical presentation, diagnosis, and management. BMJ 2021, 374, n1560. [Google Scholar] [CrossRef] [PubMed]
- Aasen, T.; Mesnil, M.; Naus, C.C.; Lampe, P.D.; Laird, D.W. Gap junctions and cancer: Communicating for 50 years. Nat. Rev. Cancer 2016, 16, 775–788. [Google Scholar] [CrossRef] [PubMed]
- Dahl, G.; Levine, E.; Rabadan-Diehl, C.; Werner, R. Cell/cell channel formation involves disulfide exchange. Eur. J. Biochem. 1991, 197, 141–144. [Google Scholar] [CrossRef] [PubMed]
- Penuela, S.; Gehi, R.; Laird, D.W. The biochemistry and function of pannexin channels. Biochim. Et Biophys. Acta (BBA)-Biomembr. 2013, 1828, 15–22. [Google Scholar] [CrossRef]
- Foote, C.I.; Zhou, L.; Zhu, X.; Nicholson, B.J. The Pattern of Disulfide Linkages in the Extracellular Loop Regions of Connexin 32 Suggests a Model for the Docking Interface of Gap Junctions. J. Cell Biol. 1998, 140, 1187–1197. [Google Scholar] [CrossRef]
- Lohman, A.W.; Isakson, B.E. Differentiating connexin hemichannels and pannexin channels in cellular ATP release. FEBS Lett. 2014, 588, 1379–1388. [Google Scholar] [CrossRef]
- Boassa, D.; Nguyen, P.; Hu, J.; Ellisman, M.H.; Sosinsky, G.E. Pannexin2 oligomers localize in the membranes of endosomal vesicles in mammalian cells while Pannexin1 channels traffic to the plasma membrane. Front. Cell. Neurosci. 2015, 8, 468. [Google Scholar] [CrossRef]
- Giaume, C.; Leybaert, L.; Naus, C.C.; Sáez, J.C. Connexin and pannexin hemichannels in brain glial cells: Properties, pharmacology, and roles. Front. Pharmacol. 2013, 4, 88. [Google Scholar] [CrossRef]
- Liang, Y.; Diehn, M.; Watson, N.; Bollen, A.W.; Aldape, K.D.; Nicholas, M.K.; Lamborn, K.R.; Berger, M.S.; Botstein, D.; Brown, P.O.; et al. Gene expression profiling reveals molecularly and clinically distinct subtypes of glioblastoma multiforme. Proc. Natl. Acad. Sci. USA 2005, 102, 5814–5819. [Google Scholar] [CrossRef]
- Scemes, E.; Velíšková, J. Exciting and not so exciting roles of pannexins. Neurosci. Lett. 2019, 695, 25–31. [Google Scholar] [CrossRef] [PubMed]
- Lai, C.P.K.; Bechberger, J.F.; Naus, C.C. Pannexin2 as a novel growth regulator in C6 glioma cells. Oncogene 2009, 28, 4402–4408. [Google Scholar] [CrossRef] [PubMed]
- Burnstock, G. Purinergic Signalling and Neurological Diseases: An Update. CNS Neurol. Disord.-Drug Targets 2017, 16, 257–265. [Google Scholar] [CrossRef] [PubMed]
- Galvão, R.P.; Zong, H. Inflammation and Gliomagenesis: Bi-Directional Communication at Early and Late Stages of Tumor Progression. Curr. Pathobiol. Rep. 2013, 1, 19–28. [Google Scholar] [CrossRef]
- Morshed, R.A.; Young, J.S.; Hervey-Jumper, S.L.; Berger, M.S. The management of low-grade gliomas in adults. J. Neurosurg. Sci. 2019, 63, 450–457. [Google Scholar] [CrossRef]
- Di Virgilio, F.; Schmalzing, G.; Markwardt, F. The Elusive P2X7 Macropore. Trends Cell Biol. 2018, 28, 392–404. [Google Scholar] [CrossRef]
- Douguet, L.; Janho dit Hreich, S.; Benzaquen, J.; Seguin, L.; Juhel, T.; Dezitter, X.; Duranton, C.; Ryffel, B.; Kanellopoulos, J.; Delarasse, C.; et al. A small-molecule P2RX7 activator promotes anti-tumor immune responses and sensitizes lung tumor to immunotherapy. Nat. Commun. 2021, 12. [Google Scholar] [CrossRef]
- North, R.A. P2X3receptors and peripheral pain mechanisms. J. Physiol. 2004, 554, 301–308. [Google Scholar] [CrossRef]
- Kaan, T.K.Y.; Yip, P.K.; Patel, S.; Davies, M.; Marchand, F.; Cockayne, D.A.; Nunn, P.A.; Dickenson, A.H.; Ford, A.P.D.W.; Zhong, Y.; et al. Systemic blockade of P2X3 and P2X2/3 receptors attenuates bone cancer pain behaviour in rats. Brain 2010, 133, 2549–2564. [Google Scholar] [CrossRef]
- Stepulak, A.; Luksch, H.; Gebhardt, C.; Uckermann, O.; Marzahn, J.; Sifringer, M.; Rzeski, W.; Staufner, C.; Brocke, K.S.; Turski, L. Expression of glutamate receptor subunits in human cancers. Histochem. Cell Biol. 2009, 132, 435–445. [Google Scholar] [CrossRef]
- Gallo, S.; Vitacolonna, A.; Crepaldi, T. NMDA Receptor and Its Emerging Role in Cancer. Int. J. Mol. Sci. 2023, 24, 2540. [Google Scholar] [CrossRef] [PubMed]
- Lee, C.-Y.; Lai, H.-Y.; Chiu, A.; Chan, S.-H.; Hsiao, L.-P.; Lee, S.-T. The effects of antiepileptic drugs on the growth of glioblastoma cell lines. J. Neuro-Oncol. 2016, 127, 445–453. [Google Scholar] [CrossRef] [PubMed]
- Tatsuoka, J.; Sano, E.; Hanashima, Y.; Yagi, C.; Yamamuro, S.; Sumi, K.; Hara, H.; Takada, K.; Kanemaru, K.; Komine-Aizawa, S.; et al. Anti-tumor effects of perampanel in malignant glioma cells. Oncol Lett. 2022, 24, 421. [Google Scholar] [CrossRef]
- Hung, T.-Y.; Huang, H.-Y.I.; Wu, S.-N.; Huang, C.-W. Depressive effectiveness of vigabatrin (γ-vinyl-GABA), an antiepileptic drug, in intermediate-conductance calcium-activated potassium channels in human glioma cells. BMC Pharmacol. Toxicol. 2021, 22, 6. [Google Scholar] [CrossRef]
- Sperling, S.; Aung, T.; Martin, S.; Rohde, V.; Ninkovic, M. Riluzole: A potential therapeutic intervention in human brain tumor stem-like cells. Oncotarget 2017, 8, 96697–96709. [Google Scholar] [CrossRef] [PubMed]
- Herner, A.; Sauliunaite, D.; Michalski, C.W.; Erkan, M.; Oliveira, T.D.; Abiatari, I.; Kong, B.; Esposito, I.; Friess, H.; Kleeff, J. Glutamate increases pancreatic cancer cell invasion and migration via AMPA receptor activation and Kras-MAPK signaling. Int. J. Cancer 2011, 129, 2349–2359. [Google Scholar] [CrossRef]
- North, W.G.; Gao, G.; Jensen, A.; Memoli, V.A.; Du, J. NMDA receptors are expressed by small-cell lung cancer and are potential targets for effective treatment. Clin. Pharmacol. Adv. Appl. 2010, 2, 31–40. [Google Scholar]
- North, W.G.; Liu, F.; Tian, R.; Abbasi, H.; Akerman, B. NMDA receptors are expressed in human ovarian cancer tissues and human ovarian cancer cell lines. Clin. Pharmacol. Adv. Appl. 2015, 7, 111–117. [Google Scholar] [CrossRef]
- Kleinerman, R.A.; Brinton, L.A.; Hoover, R.; Fraumeni, J.F., Jr. Diazepam use and progression of breast cancer. Cancer Res. 1984, 44, 1223–1225. [Google Scholar]
- Phiel, C.J.; Zhang, F.; Huang, E.Y.; Guenther, M.G.; Lazar, M.A.; Klein, P.S. Histone deacetylase is a direct target of valproic acid, a potent anticonvulsant, mood stabilizer, and teratogen. J. Biol. Chem. 2001, 276, 36734–36741. [Google Scholar] [CrossRef]
- Stella, M.; Baiardi, G.; Pasquariello, S.; Sacco, F.; Dellacasagrande, I.; Corsaro, A.; Mattioli, F.; Barbieri, F. Antitumor Potential of Antiepileptic Drugs in Human Glioblastoma: Pharmacological Targets and Clinical Benefits. Biomedicines 2023, 11, 582. [Google Scholar] [CrossRef] [PubMed]
- Gottlicher, M.; Minucci, S.; Zhu, P.; Kramer, O.H.; Schimpf, A.; Giavara, S.; Sleeman, J.P.; Lo Coco, F.; Nervi, C.; Pelicci, P.G.; et al. Valproic acid defines a novel class of HDAC inhibitors inducing differentiation of transformed cells. EMBO J. 2001, 20, 6969–6978. [Google Scholar] [CrossRef]
- Van Nifterik, K.A.; Van den Berg, J.; Slotman, B.J.; Lafleur, M.V.; Sminia, P.; Stalpers, L.J. Valproic acid sensitizes human glioma cells for temozolomide and gamma-radiation. J. Neurooncol. 2012, 107, 61–67. [Google Scholar] [CrossRef] [PubMed]
- Han, W.; Guan, W. Valproic Acid: A Promising Therapeutic Agent in Glioma Treatment. Front. Oncol. 2021, 11, 687362. [Google Scholar] [CrossRef] [PubMed]
- Su, J.M.; Murray, J.C.; McNall-Knapp, R.Y.; Bowers, D.C.; Shah, S.; Adesina, A.M.; Paulino, A.C.; Jo, E.; Mo, Q.; Baxter, P.A.; et al. A phase 2 study of valproic acid and radiation, followed by maintenance valproic acid and bevacizumab in children with newly diagnosed diffuse intrinsic pontine glioma or high-grade glioma. Pediatr. Blood Cancer 2020, 67, e28283. [Google Scholar] [CrossRef] [PubMed]
- Li, L.; Zhang, C.; Wang, Z.; Guo, Y.; Wang, Y.; Fan, X.; Jiang, T. Expression changes in ion channel and immunity genes are associated with glioma-related epilepsy in patients with diffuse gliomas. J. Cancer Res. Clin. Oncol. 2022, 148, 2793–2802. [Google Scholar] [CrossRef]
- Szoka, L.; Palka, J. Capsaicin up-regulates pro-apoptotic activity of thiazolidinediones in glioblastoma cell line. Biomed Pharm. 2020, 132, 110741. [Google Scholar] [CrossRef]
- Liu, T.; Wang, G.; Tao, H.; Yang, Z.; Wang, Y.; Meng, Z.; Cao, R.; Xiao, Y.; Wang, X.; Zhou, J. Capsaicin mediates caspases activation and induces apoptosis through P38 and JNK MAPK pathways in human renal carcinoma. BMC Cancer 2016, 16, 790. [Google Scholar] [CrossRef]
- Kim, J.Y.; Kim, E.H.; Kim, S.U.; Kwon, T.K.; Choi, K.S. Capsaicin sensitizes malignant glioma cells to TRAIL-mediated apoptosis via DR5 upregulation and survivin downregulation. Carcinogenesis 2010, 31, 367–375. [Google Scholar] [CrossRef]
- Amantini, C.; Ballarini, P.; Caprodossi, S.; Nabissi, M.; Morelli, M.B.; Lucciarini, R.; Cardarelli, M.A.; Mammana, G.; Santoni, G. Triggering of transient receptor potential vanilloid type 1 (TRPV1) by capsaicin induces Fas/CD95-mediated apoptosis of urothelial cancer cells in an ATM-dependent manner. Carcinogenesis 2009, 30, 1320–1329. [Google Scholar] [CrossRef]
- Pattni, B.S.; Chupin, V.V.; Torchilin, V.P. New Developments in Liposomal Drug Delivery. Chem. Rev. 2015, 115, 10938–10966. [Google Scholar] [CrossRef] [PubMed]
- Zhu, Y.; Liang, J.; Gao, C.; Wang, A.; Xia, J.; Hong, C.; Zhong, Z.; Zuo, Z.; Kim, J.; Ren, H.; et al. Multifunctional ginsenoside Rg3-based liposomes for glioma targeting therapy. J. Control. Release 2021, 330, 641–657. [Google Scholar] [CrossRef] [PubMed]
- Ying, M.; Zhan, C.; Wang, S.; Yao, B.; Hu, X.; Song, X.; Zhang, M.; Wei, X.; Xiong, Y.; Lu, W. Liposome-Based Systemic Glioma-Targeted Drug Delivery Enabled by All-d Peptides. ACS Appl. Mater. Interfaces 2016, 8, 29977–29985. [Google Scholar] [CrossRef]
- Wang, G.; Wang, J.J.; Chen, X.L.; Du, S.M.; Li, D.S.; Pei, Z.J.; Lan, H.; Wu, L.B. The JAK2/STAT3 and mitochondrial pathways are essential for quercetin nanoliposome-induced C6 glioma cell death. Cell Death Dis. 2013, 4, e746. [Google Scholar] [CrossRef] [PubMed]
- Xu, H.; Li, C.; Wei, Y.; Zheng, H.; Zheng, H.; Wang, B.; Piao, J.G.; Li, F. Angiopep-2-modified calcium arsenite-loaded liposomes for targeted and pH-responsive delivery for anti-glioma therapy. Biochem. Biophys. Res. Commun. 2021, 551, 14–20. [Google Scholar] [CrossRef]
- Mojarad-Jabali, S.; Farshbaf, M.; Hemmati, S.; Sarfraz, M.; Motasadizadeh, H.; Shahbazi Mojarrad, J.; Atyabi, F.; Zakeri-Milani, P.; Valizadeh, H. Comparison of three synthetic transferrin mimetic small peptides to promote the blood-brain barrier penetration of vincristine liposomes for improved glioma targeted therapy. Int. J. Pharm. 2022, 613, 121395. [Google Scholar] [CrossRef]
- Zheng, Z.; Zhang, J.; Jiang, J.; He, Y.; Zhang, W.; Mo, X.; Kang, X.; Xu, Q.; Wang, B.; Huang, Y. Remodeling tumor immune microenvironment (TIME) for glioma therapy using multi-targeting liposomal codelivery. J. Immunother. Cancer 2020, 8, e000207. [Google Scholar] [CrossRef]
- Wang, X.; Zhao, Y.; Dong, S.; Lee, R.J.; Yang, D.; Zhang, H.; Teng, L. Cell-Penetrating Peptide and Transferrin Co-Modified Liposomes for Targeted Therapy of Glioma. Molecules 2019, 24, 3540. [Google Scholar] [CrossRef]




{kind=link}
{kind=link}
{kind=link}
| Receptor | Receptor Subtype | Ligand | Ionic Conductance |
|---|---|---|---|
| Ionotropic Glutamate | AMPA | L-Glutamate | Na+, K+, Ca2+ |
| Kainate | L-Glutamate | Na+, K+ | |
| NMDA | L-Glutamate | Na+, K+, Ca2+ | |
| ATP-gated channel | P2X | ATP | Na+, K+, Ca2+ |
| Cys-Loop | nACh | Acetylcholine | Na+, K+, Ca2+ |
| 5-HT3 | Serotonin | Na+, K+ | |
| GABAA | GABA | Cl−, HCO3− | |
| Glycine | Glycine | Cl− |
| Gene | Gene Description | Weight |
|---|---|---|
| CACNA1D | calcium channel, voltage-dependent, L type, alpha 1D subunit | −1 |
| CLCN6 | chloride channel, voltage-sensitive 6 | −1 |
| CLIC1 | chloride intracellular channel 1 | 1 |
| CLIC4 | chloride intracellular channel 4 | 1 |
| GLRB | glycine receptor, beta | −1 |
| GRIA2 | glutamate receptor, ionotropic, AMPA 2 | −1 |
| GRID1 | glutamate receptor, ionotropic, delta 1 | −1 |
| KCNAB1 | potassium voltage-gated channel, shaker-related subfamily, beta member 1 | −1 |
| KCNB1 | potassium voltage-gated channel, Shab-related subfamily, member 1 | −1 |
| KCND2 | potassium voltage-gated channel, Shal-related subfamily, member 2 | −1 |
| KCNJ10 | potassium inwardly-rectifying channel, subfamily J, member 10 | −1 |
| KCNMA1 | potassium large conductance calcium-activated channel, subfamily M, alpha member 1 | −1 |
| KCNN3 | potassium intermediate/small conductance calcium-activated channel, subfamily N, member 3 | −1 |
| KCNQ5 | potassium voltage-gated channel, KQT-like subfamily, member 5 | −1 |
| NALCN | sodium leak channel, non-selective | −1 |
| P2RX7 | purinergic receptor P2X, ligand-gated ion channel, 7 | −1 |
| SCN1A | sodium channel, voltage-gated, type I, alpha subunit | −1 |
| VDAC2 | voltage-dependent anion channel 2 | −1 |
| Study | Title | Status | Receptor Type | Conditions |
|---|---|---|---|---|
| NCT02082821 (Boston, MA, USA) | A P2X7R Single Nucleotide Mutation Promotes Chronic Allograft Vasculopathy | Completed | P2RX7 | Cardiac Allograft Vasculopathy |
| NCT03918616 (Pisa, Italy) | P2X7 Receptor, Inflammation, and Neurodegenerative Diseases | Completed | P2RX7 | Neuro-Degenerative Disease |
| NCT02293811 (Alpes-Maritimes, France) | Decoding of the Expression of Tumor Suppressor P2RX7 in Inflammatory and Malignant Colonic Mucosa (P2RX7) | Unknown | P2RX7 | Crohn’s Disease-Associated Colorectal Adenocarcinoma |
| NCT04122937 (Pisa, Italy) | Defining Inflammation Related to Peritoneal Carcinomatosis in Women With Ovarian or Colon Cancer (CarFlog) | Completed | P2RX7 | Peritoneal Carcinomatosis Ovarian Cancer Colon Cancer |
| NCT05225883 (Lyon, France) | GWAS in NMDAR Encephalitis | Recruiting | Glutamate NMDA | Autoimmune Encephalitis |
| NCT05503264 (Birmingham, AL, USA) | A Study To Evaluate The Efficacy, Safety, Pharmacokinetics, And Pharmacodynamics Of Satralizumab In Patients With Anti-N-Methyl-D-Aspartic Acid Receptor (NMDAR) Or Anti-Leucine-Rich Glioma-Inactivated 1 (LGI1) Encephalitis (Cielo) | Recruiting | Glutamate NMDA | NMDAR Autoimmune Encephalitis LGI1 Autoimmune Encephalitis |
| NCT02654964 (Washington, D.C., MA, USA) | Cancer Stem Cell High-Throughput Drug Screening Study | Recruiting | Glutamate NMDA | GBM |
| NCT02363933 (Durham, NC, USA) | Perampanel in Seizure Patients With Primary Glial Brain Tumors | Completed | Glutamate AMPA | Brain Tumor, Primary |
| NCT03636958 (Paca, France; Marseille, France) | Efficacy and Safety of Perampanel in Combination in Glioma-refractory Epilepsy | Withdrawn | Glutamate AMPA | Refractory Epilepsy |
| NCT03229278 (New Brunswick, NJ, USA) | Trigriluzole with Nivolumab and Pembrolizumab in Treating Patients with Metastatic of Unresectable Solid Malignancies or Lymphoma | Completed | Cys-loop | Lymphoma Unresectable or metastatic malignancies |
| NCT00879437 (Dallas, Fort Worth, Houston, and San Antonio TX, USA; Oklahoma City, OK, USA) | Valproic acid, radiation, and bevacizumab in children with high-grade gliomas or diffuse intrinsic pontine glioma | Completed | Cys-loop | Pediatric high-grade glioma or brainstem glioma |
| NCT00302159 (Bethesda, MA, USA; Philadelphia, PA, USA; Richmond, VA, USA) | Valproic acid with temozolomide and radiation therapy to treat brain tumors | Completed | Cys-loop | GBM High-grade giomas |
| NCT03048084 (The Hague, Netherlands; Rotterdand, Netherlands; Amsterdam, Netherlands) | Seizure treatment in glioma (STING) | Recruiting | Cys-loop | Glioma |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Hey, G.; Rao, R.; Carter, A.; Reddy, A.; Valle, D.; Patel, A.; Patel, D.; Lucke-Wold, B.; Pomeranz Krummel, D.; Sengupta, S. Ligand-Gated Ion Channels: Prognostic and Therapeutic Implications for Gliomas. J. Pers. Med. 2023, 13, 853. https://doi.org/10.3390/jpm13050853
Hey G, Rao R, Carter A, Reddy A, Valle D, Patel A, Patel D, Lucke-Wold B, Pomeranz Krummel D, Sengupta S. Ligand-Gated Ion Channels: Prognostic and Therapeutic Implications for Gliomas. Journal of Personalized Medicine. 2023; 13(5):853. https://doi.org/10.3390/jpm13050853
Chicago/Turabian StyleHey, Grace, Rohan Rao, Ashley Carter, Akshay Reddy, Daisy Valle, Anjali Patel, Drashti Patel, Brandon Lucke-Wold, Daniel Pomeranz Krummel, and Soma Sengupta. 2023. "Ligand-Gated Ion Channels: Prognostic and Therapeutic Implications for Gliomas" Journal of Personalized Medicine 13, no. 5: 853. https://doi.org/10.3390/jpm13050853