
MLN4924 Promotes Self-Renewal of Limbal Stem Cells and Ocular Surface Restoration
 , ,
, , {kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Materials and Methods
2.1. Preparation of Feeder Cells
2.2. Isolation and Culture of LSCs
2.3. Crystal Violet Staining
2.4. Immunofluorescence Staining
2.5. RNA Extraction and Polymerase Chain Reaction (PCR)
2.6. Western Blotting
2.7. Corneal Epithelial Scratch Model
2.8. Statistical Analysis
3. Results
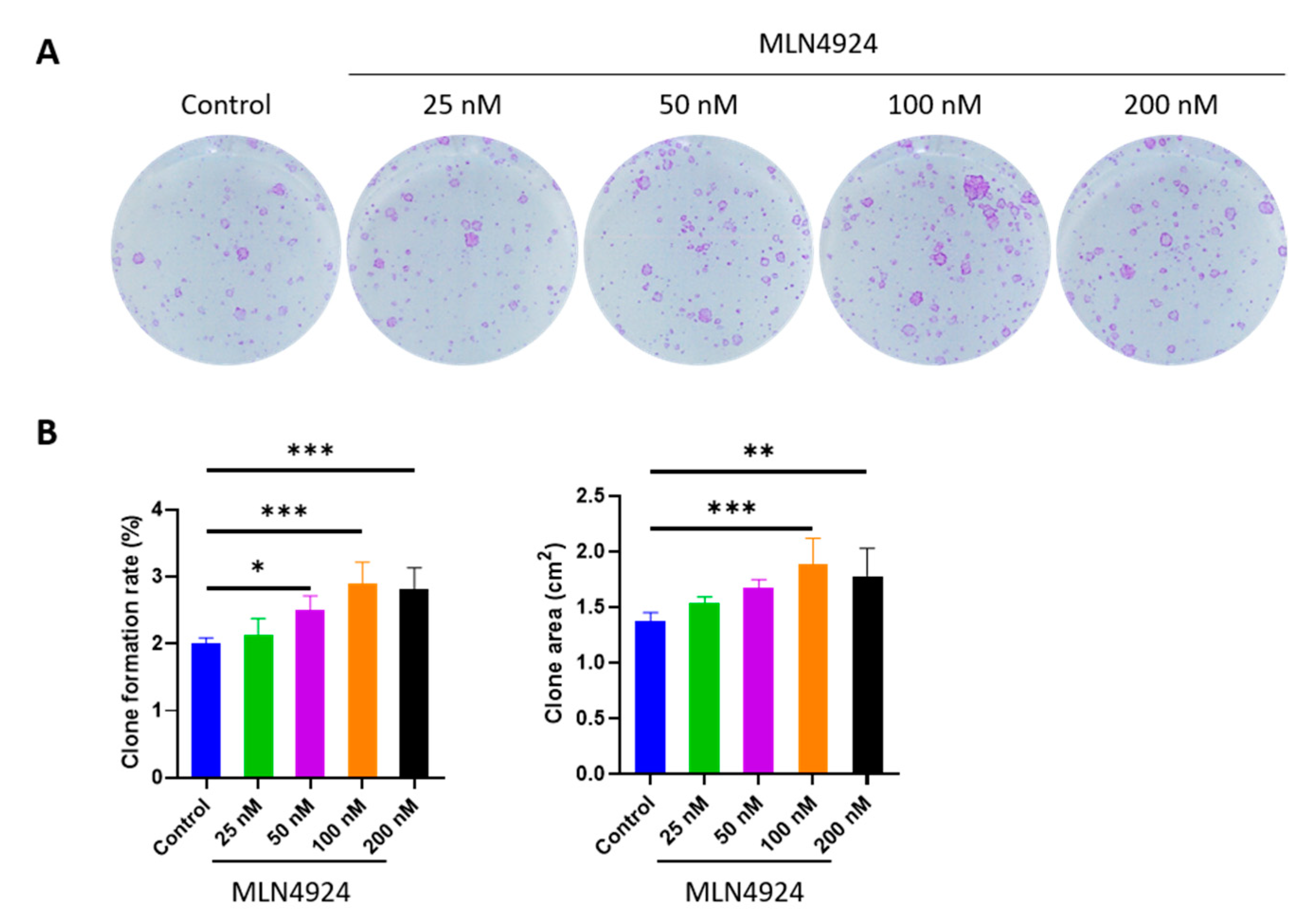
3.1. MLN4924 Promotes Clone Formation Rate and Clone Area of LSCs
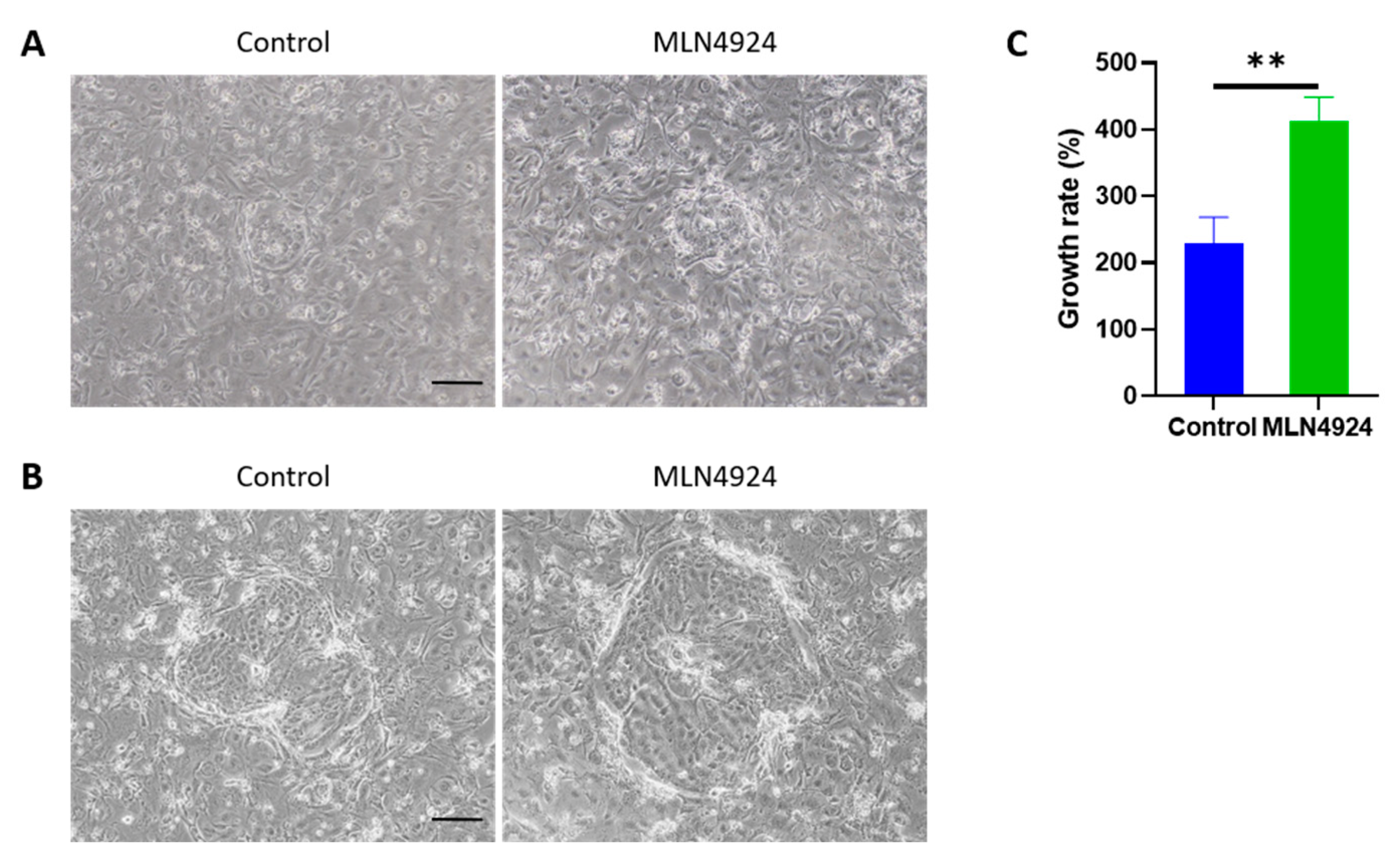
3.2. MLN4924 Improves the Proliferative Capacity of LSCs In Vitro
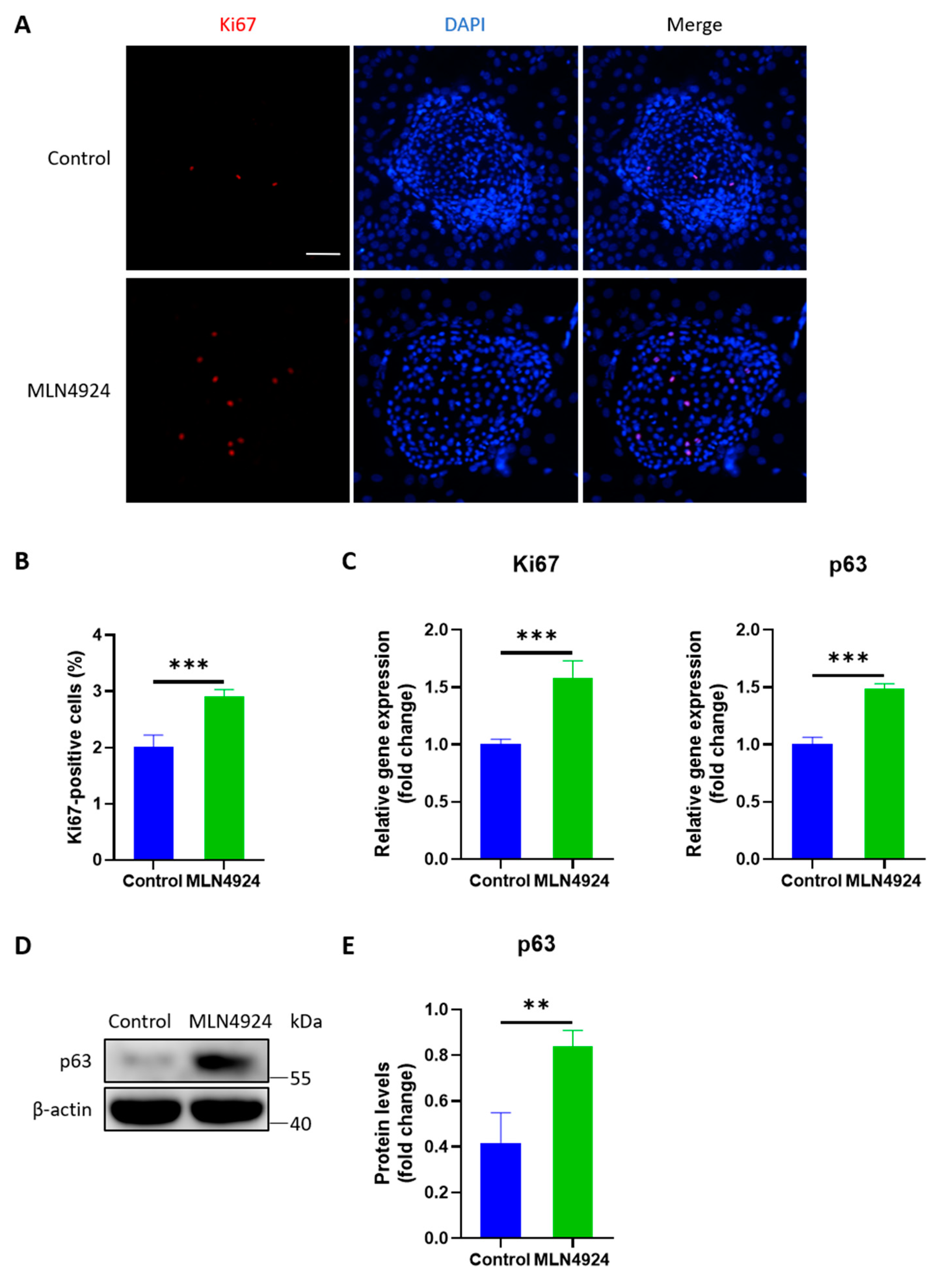
3.3. MLN4924 Increases the Expression Levels of Ki67 and p63 in LSCs
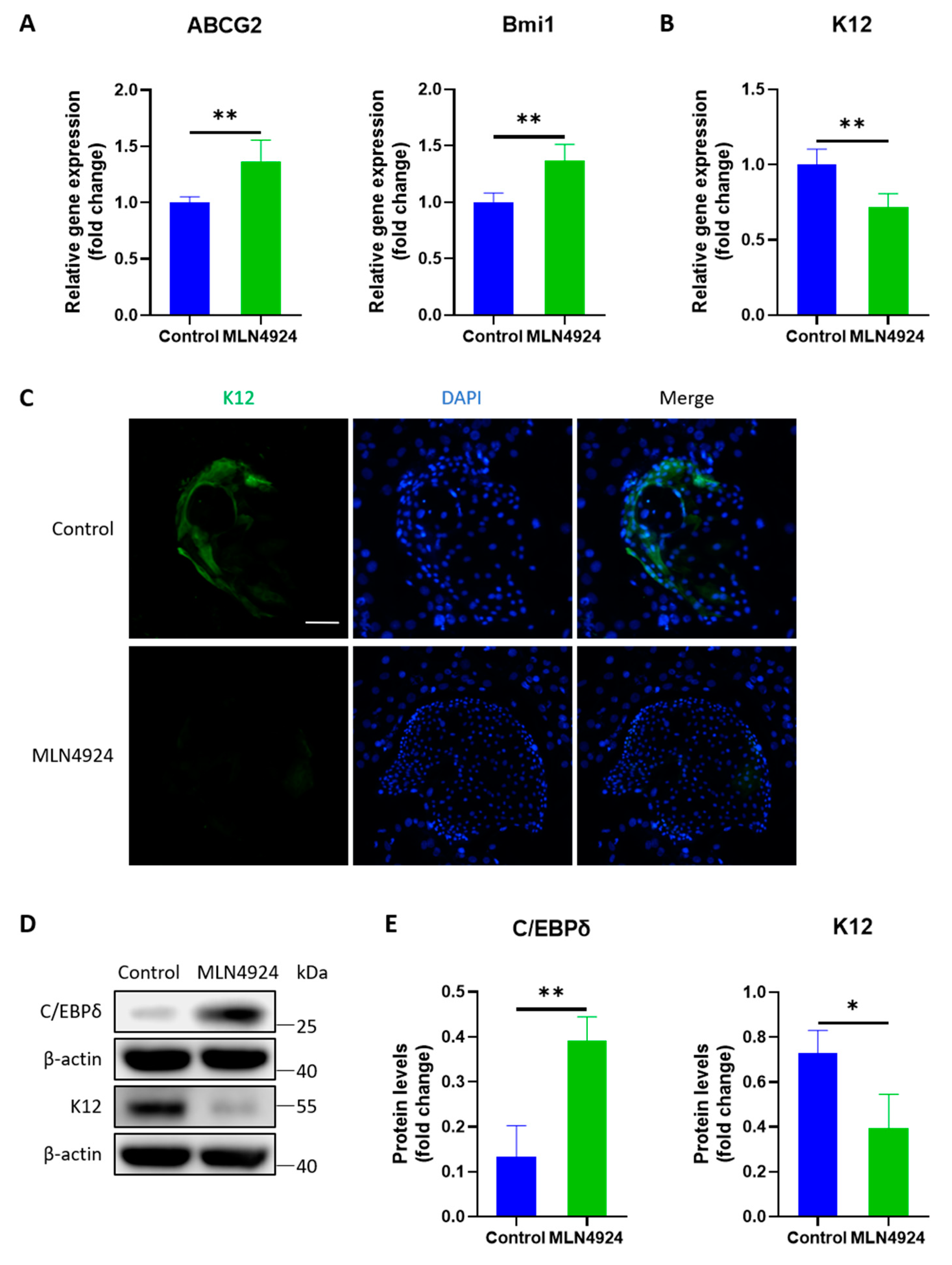
3.4. MLN4924 Enhances the Preservation of Stemness of LSCs
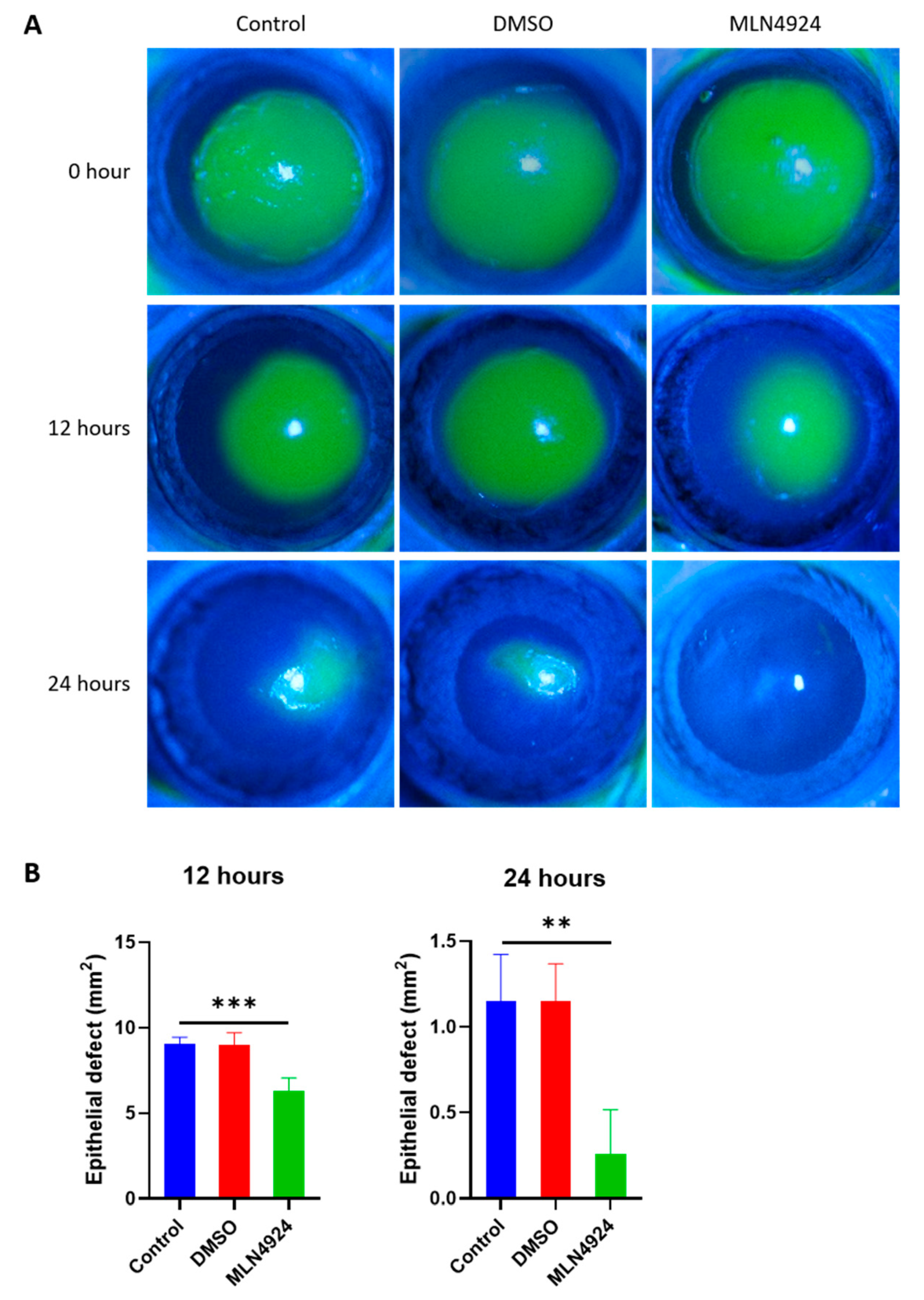
3.5. MLN4924 Promotes Corneal Wound Healing
4. Discussion
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Osei-Bempong, C.; Figueiredo, F.C.; Lako, M. The limbal epithelium of the eye—A review of limbal stem cell biology, disease and treatment. Bioessays 2013, 35, 211–219. [Google Scholar] [CrossRef] [PubMed]
- DelMonte, D.W.; Kim, T. Anatomy and physiology of the cornea. J. Cataract. Refract. Surg. 2011, 37, 588–598. [Google Scholar] [CrossRef]
- Thoft, R.A.; Friend, J. The X, Y, Z hypothesis of corneal epithelial maintenance. Investig. Ophthalmol Vis Sci. 1983, 24, 1442–1443. [Google Scholar]
- Sasamoto, Y.; Ksander, B.R.; Frank, M.H.; Frank, N.Y. Repairing the corneal epithelium using limbal stem cells or alternative cell-based therapies. Expert. Opin. Biol. Ther. 2018, 18, 505–513. [Google Scholar] [CrossRef]
- Vazirani, J.; Nair, D.; Shanbhag, S.; Wurity, S.; Ranjan, A.; Sangwan, V. Limbal Stem Cell Deficiency-Demography and Underlying Causes. Am. J. Ophthalmol. 2018, 188, 99–103. [Google Scholar] [CrossRef]
- Oliva, M.S.; Schottman, T.; Gulati, M. Turning the tide of corneal blindness. Indian J. Ophthalmol. 2012, 60, 423–427. [Google Scholar] [CrossRef] [PubMed]
- Chirila, T.V.; Harkin, D. Biomaterials and Regenerative Medicine in Ophthalmology, 1st ed.; Elsevier: Amsterdam, Netherlands, 2009; Available online: https://www.elsevier.com/books/biomaterials-and-regenerative-medicine-in-ophthalmology/chirila/978-1-84569-443-2 (accessed on 29 June 2020).
- Kolli, S.; Ahmad, S.; Mudhar, H.S.; Meeny, A.; Lako, M.; Figueiredo, F.C. Successful application of ex vivo expanded human autologous oral mucosal epithelium for the treatment of total bilateral limbal stem cell deficiency. Stem Cells 2014, 32, 2135–2146. [Google Scholar] [CrossRef]
- Gonzalez, G.; Sasamoto, Y.; Ksander, B.R.; Frank, M.H.; Frank, N.Y. Limbal stem cells: Identity, developmental origin, and therapeutic potential. Wiley Interdiscip. Rev. Dev. Biol. 2018, 7, e303. [Google Scholar] [CrossRef]
- Zhou, M.; Li, X.M.; Lavker, R.M. Transcriptional profiling of enriched populations of stem cells versus transient amplifying cells. A comparison of limbal and corneal epithelial basal cells. J. Biol. Chem. 2006, 281, 19600–19609. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zhou, L.; Zhang, W.; Sun, Y.; Jia, L. Protein neddylation and its alterations in human cancers for targeted therapy. Cell Signal. 2018, 44, 92–102. [Google Scholar] [CrossRef]
- Xirodimas, D.P. Novel substrates and functions for the ubiquitin-like molecule NEDD8. Biochem. Soc. Trans. 2008, 36 Pt 5, 802–806. [Google Scholar] [CrossRef] [Green Version]
- Zhao, Y.; Morgan, M.A.; Sun, Y. Targeting Neddylation pathways to inactivate cullin-RING ligases for anticancer therapy. Antioxid. Redox. Signal. 2014, 21, 2383–2400. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Liddy, K.A.; White, M.Y.; Cordwell, S.J. Functional decorations: Post-translational modifications and heart disease delineated by targeted proteomics. Genome Med. 2013, 5, 20. [Google Scholar] [CrossRef] [Green Version]
- Bailly, A.P.; Perrin, A.; Serrano-Macia, M.; Maghames, C.; Leidecker, O.; Trauchessec, H.; Martinez-Chantar, M.L.; Gartner, A.; Xirodimas, D.P. The Balance between Mono- and NEDD8-Chains Controlled by NEDP1 upon DNA Damage Is a Regulatory Module of the HSP70 ATPase Activity. Cell Rep. 2019, 29, 212–224.e8. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zou, T.; Zhang, J. Diverse and pivotal roles of neddylation in metabolism and immunity. FEBS J. 2021, 288, 3884–3912. [Google Scholar] [CrossRef]
- Soucy, T.A.; Smith, P.G.; Milhollen, M.A.; Berger, A.J.; Gavin, J.M.; Adhikari, S.; Brownell, J.E.; Burke, K.E.; Cardin, D.P.; Critchley, S.; et al. An inhibitor of NEDD8-activating enzyme as a new approach to treat cancer. Nature 2009, 458, 732–736. [Google Scholar] [CrossRef] [PubMed]
- Zhao, Y.; Sun, Y. Cullin-RING Ligases as attractive anti-cancer targets. Curr. Pharm. Des. 2013, 19, 3215–3225. [Google Scholar] [CrossRef] [Green Version]
- Xie, M.; Guo, H.; Lou, G.; Yao, J.; Liu, Y.; Sun, Y.; Yang, Z.; Zheng, M. Neddylation inhibitor MLN4924 has anti-HBV activity via modulating the ERK-HNF1α-C/EBPα-HNF4α axis. J. Cell Mol. Med. 2021, 25, 840–854. [Google Scholar] [CrossRef]
- Zhou, X.; Tan, M.; Nyati, M.K.; Zhao, Y.; Wang, G.; Sun, Y. Blockage of neddylation modification stimulates tumor sphere formation in vitro and stem cell differentiation and wound healing in vivo. Proc. Natl. Acad. Sci. USA 2016, 113, E2935–E2944. [Google Scholar] [CrossRef] [Green Version]
- Skeens, H.M.; Brooks, B.P.; Holland, E.J. Congenital aniridia variant: Minimally abnormal irides with severe limbal stem cell deficiency. Ophthalmology 2011, 118, 1260–1264. [Google Scholar] [CrossRef]
- Ramos, T.; Scott, D.; Ahmad, S. An Update on Ocular Surface Epithelial Stem Cells: Cornea and Conjunctiva. Stem Cells Int. 2015, 2015, 601731. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Robertson, S.Y.T.; Roberts, J.S.; Deng, S.X. Regulation of Limbal Epithelial Stem Cells: Importance of the Niche. Int. J. Mol. Sci. 2021, 22, 11975. [Google Scholar] [CrossRef] [PubMed]
- Notara, M.; Lentzsch, A.; Coroneo, M.; Cursiefen, C. The Role of Limbal Epithelial Stem Cells in Regulating Corneal (Lymph)angiogenic Privilege and the Micromilieu of the Limbal Niche following UV Exposure. Stem Cells Int. 2018, 2018, 8620172. [Google Scholar] [CrossRef] [Green Version]
- Yoon, J.J.; Ismail, S.; Sherwin, T. Limbal stem cells: Central concepts of corneal epithelial homeostasis. World J. Stem Cells 2014, 6, 391–403. [Google Scholar] [CrossRef]
- Chung, E.H.; Bukusoglu, G.; Zieske, J.D. Localization of corneal epithelial stem cells in the developing rat. Investig. Ophthalmol. Vis. Sci. 1992, 33, 2199–2206. [Google Scholar]
- Pellegrini, G.; Golisano, O.; Paterna, P.; Lambiase, A.; Bonini, S.; Rama, P.; De Luca, M. Location and clonal analysis of stem cells and their differentiated progeny in the human ocular surface. J. Cell Biol. 1999, 145, 769–782. [Google Scholar] [CrossRef]
- Goldberg, M.F.; Bron, A.J. Limbal palisades of Vogt. Trans. Am. Ophthalmol. Soc. 1982, 80, 155–171. [Google Scholar]
- Di Girolamo, N. Moving epithelia: Tracking the fate of mammalian limbal epithelial stem cells. Prog. Retin. Eye Res. 2015, 48, 203–225. [Google Scholar] [CrossRef]
- Gericke, A.; Wasielica-Poslednik, J.; Zimmermann, M.; Musayeva, A. Expansion und Transplantation limbaler Stammzellen zur Regeneration der kornealen Oberfläche [Expansion and Transplantation of Limbal Stem Cells for Corneal Surface Regeneration]. Klin. Monbl. Augenheilkd. 2019, 236, 777–783. [Google Scholar] [CrossRef]
- Park, H.S.; Ju, U.I.; Park, J.W.; Song, J.Y.; Shin, D.H.; Lee, K.H.; Jeong, L.S.; Yu, J.; Lee, H.W.; Cho, J.Y.; et al. PPARγ neddylation essential for adipogenesis is a potential target for treating obesity. Cell Death Differ. 2016, 23, 1296–1311. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zou, J.; Ma, W.; Li, J.; Littlejohn, R.; Zhou, H.; Kim, I.M.; Fulton, D.J.R.; Chen, W.; Weintraub, N.L.; Zhou, J.; et al. Neddylation mediates ventricular chamber maturation through repression of Hippo signaling. Proc. Natl. Acad. Sci. USA 2018, 115, E4101–E4110. [Google Scholar] [CrossRef] [Green Version]
- Li, L.; Cao, Y.; Wu, H.; Ye, X.; Zhu, Z.; Xing, G.; Shen, C.; Barik, A.; Zhang, B.; Xie, X.; et al. Enzymatic Activity of the Scaffold Protein Rapsyn for Synapse Formation. Neuron 2016, 92, 1007–1019. [Google Scholar] [CrossRef] [Green Version]
- Deng, Q.; Zhang, J.; Gao, Y.; She, X.; Wang, Y.; Wang, Y.; Ge, X. MLN4924 protects against bleomycin-induced pulmonary fibrosis by inhibiting the early inflammatory process. Am. J. Transl. Res. 2017, 9, 1810–1821. [Google Scholar]
- Ai, T.J.; Sun, J.Y.; Du, L.J.; Shi, C.; Li, C.; Sun, X.N.; Liu, Y.; Li, L.; Xia, Z.; Jia, L.; et al. Inhibition of neddylation by MLN4924 improves neointimal hyperplasia and promotes apoptosis of vascular smooth muscle cells through p53 and p62. Cell Death Differ. 2018, 25, 319–329. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bhatia, S.; Pavlick, A.C.; Boasberg, P.; Thompson, J.A.; Mulligan, G.; Pickard, M.D.; Faessel, H.; Dezube, B.J.; Hamid, O. A phase I study of the investigational NEDD8-activating enzyme inhibitor pevonedistat (TAK-924/MLN4924) in patients with metastatic melanoma. Investig. New Drugs 2016, 34, 439–449. [Google Scholar] [CrossRef] [Green Version]
- Khalife, J.; Radomska, H.S.; Santhanam, R.; Huang, X.; Neviani, P.; Saultz, J.; Wang, H.; Wu, Y.-Z.; Alachkar, H.; Anghelina, M.; et al. Pharmacological targeting of miR-155 via the NEDD8-activating enzyme inhibitor MLN4924 (Pevonedistat) in FLT3-ITD acute myeloid leukemia. Leukemia 2015, 29, 1981–1992. [Google Scholar] [CrossRef] [Green Version]
- Whiteside, T.L. The tumor microenvironment and its role in promoting tumor growth. Oncogene 2008, 27, 5904–5912. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zhou, L.; Jiang, Y.; Luo, Q.; Li, L.; Jia, L. Neddylation: A novel modulator of the tumor microenvironment. Mol. Cancer 2019, 18, 77. [Google Scholar] [CrossRef] [Green Version]
- Pellegrino, N.E.; Guven, A.; Gray, K.; Shah, P.; Kasture, G.; Nastke, M.D.; Thakurta, A.; Gesta, S.; Vishnudas, V.K.; Narain, N.R.; et al. The Next Frontier: Translational Development of Ubiquitination, SUMOylation, and NEDDylation in Cancer. Int. J. Mol. Sci. 2022, 23, 3480. [Google Scholar] [CrossRef]
- Petroski, M.D. Mechanism-based neddylation inhibitor. Chem. Biol. 2010, 17, 6–8. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Liao, H.; Liu, X.J.; Blank, J.L.; Bouck, D.C.; Bernard, H.; Garcia, K.; Lightcap, E.S. Quantitative proteomic analysis of cellular protein modulation upon inhibition of the NEDD8-activating enzyme by MLN4924. Mol. Cell Proteom. 2011, 10, M111.009183. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Milhollen, M.A.; Narayanan, U.; Soucy, T.A.; Veiby, P.O.; Smith, P.G.; Amidon, B. Inhibition of NEDD8-activating enzyme induces rereplication and apoptosis in human tumor cells consistent with deregulating CDT1 turnover. Cancer Res. 2011, 71, 3042–3051. [Google Scholar] [CrossRef] [Green Version]
- Zhang, Y.; Shi, C.C.; Zhang, H.P.; Li, G.Q.; Li, S.S. MLN4924 suppresses neddylation and induces cell cycle arrest, senescence, and apoptosis in human osteosarcoma. Oncotarget 2016, 7, 45263–45274. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Tong, S.; Si, Y.; Yu, H.; Zhang, L.; Xie, P.; Jiang, W. MLN4924 (Pevonedistat), a protein neddylation inhibitor, suppresses proliferation and migration of human clear cell renal cell carcinoma. Sci. Rep. 2017, 7, 5599. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zhou, H.; Su, H.; Chen, W. Neddylation Regulates Class IIa and III Histone Deacetylases to Mediate Myoblast Differentiation. Int. J. Mol. Sci. 2021, 22, 9509. [Google Scholar] [CrossRef]
- Li, L.; Liu, B.; Dong, T.; Lee, H.W.; Yu, J.; Zheng, Y.; Gao, H.; Zhang, Y.; Chu, Y.; Liu, G.; et al. Neddylation pathway regulates the proliferation and survival of macrophages. Biochem. Biophys. Res. Commun. 2013, 432, 494–498. [Google Scholar] [CrossRef]
- Wu, M.H.; Hsu, W.B.; Chen, M.H.; Shi, C.S. Inhibition of Neddylation Suppresses Osteoclast Differentiation and Function In Vitro and Alleviates Osteoporosis In Vivo. Biomedicines 2022, 10, 2355. [Google Scholar] [CrossRef]
- Albayrak, E.; Uslu, M.; Akgol, S.; Tuysuz, E.C.; Kocabas, F. Small molecule-mediated modulation of ubiquitination and neddylation improves HSC function ex vivo. J. Cell Physiol. 2021, 236, 8122–8136. [Google Scholar] [CrossRef]
- Kammergruber, E.; Rahn, C.; Nell, B.; Gabner, S.; Egerbacher, M. Morphological and immunohistochemical characteristics of the equine corneal epithelium. Vet. Ophthalmol. 2019, 22, 778–790. [Google Scholar] [CrossRef]
- Liu, T.; Wang, Y.; Duan, H.Y.; Qu, M.L.; Yang, L.L.; Xu, Y.Y.; Zang, X.J.; Zhou, Q.J. Effects of preservation time on proliferative potential of human limbal stem/progenitor cells. Int. J. Ophthalmol. 2012, 5, 549–554. [Google Scholar] [CrossRef]
- Busin, M.; Breda, C.; Bertolin, M.; Bovone, C.; Ponzin, D.; Ferrari, S.; Barbaro, V.; Elbadawy, H.M. Corneal Epithelial Stem Cells Repopulate the Donor Area within 1 Year from Limbus Removal for Limbal Autograft. Ophthalmology 2016, 123, 2481–2488. [Google Scholar] [CrossRef] [PubMed]
- Takahashi, K.; Yamanaka, S. Induction of pluripotent stem cells from mouse embryonic and adult fibroblast cultures by defined factors. Cell 2006, 126, 663–676. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Quinn, L.M.; Secombe, J.; Hime, G.R. Myc in stem cell behaviour: Insights from Drosophila. Adv. Exp. Med. Biol. 2013, 786, 269–285. [Google Scholar] [CrossRef] [PubMed]
- Aporowicz, M.; Czopnik, P.; Kubicka, E.; Piotrowska, A.; Dziegiel, P.; Bolanowski, M.; Domoslawski, P. Minichromosome Maintenance Proteins MCM-3, MCM-5, MCM-7, and Ki-67 as Proliferative Markers in Adrenocortical Tumors. Anticancer Res. 2019, 39, 1151–1159. [Google Scholar] [CrossRef] [Green Version]
- da Silva, J.N.L.; Ranzi, A.D.; Carvalho, C.T.; Scheide, T.V.; Strey, Y.T.M.; Graziottin, T.M.; Bica, C.G. Cell Cycle Markers in the Evaluation of Bladder Cancer. Pathol. Oncol. Res. 2020, 26, 175–181. [Google Scholar] [CrossRef] [PubMed]
- Pouget, C.; Hergalant, S.; Lardenois, E.; Lacomme, S.; Houlgatte, R.; Carpentier, C.; Dehais, C.; Rech, F.; Taillandier, L.; Sanson, M.; et al. Ki-67 and MCM6 labeling indices are correlated with overall survival in anaplastic oligodendroglioma, IDH1-mutant and 1p/19q-codeleted: A multicenter study from the French POLA network. Brain Pathol. 2020, 30, 465–478. [Google Scholar] [CrossRef]
- Koster, M.I.; Kim, S.; Mills, A.A.; DeMayo, F.J.; Roop, D.R. p63 is the molecular switch for initiation of an epithelial stratification program. Genes Dev. 2004, 18, 126–131. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- DeYoung, M.P.; Johannessen, C.M.; Leong, C.O.; Faquin, W.; Rocco, J.W.; Ellisen, L.W. Tumor-specific p73 up-regulation mediates p63 dependence in squamous cell carcinoma. Cancer Res. 2006, 66, 9362–9368. [Google Scholar] [CrossRef] [Green Version]
- Ramsey, M.R.; He, L.; Forster, N.; Ory, B.; Ellisen, L.W. Physical association of HDAC1 and HDAC2 with p63 mediates transcriptional repression and tumor maintenance in squamous cell carcinoma. Cancer Res. 2011, 71, 4373–4379. [Google Scholar] [CrossRef] [Green Version]
- Kukal, S.; Guin, D.; Rawat, C.; Bora, S.; Mishra, M.K.; Sharma, P.; Paul, P.R.; Kanojia, N.; Grewal, G.K.; Kukreti, S.; et al. Multidrug efflux transporter ABCG2: Expression and regulation. Cell Mol. Life Sci. 2021, 78, 6887–6939. [Google Scholar] [CrossRef]
- Wei, L.Y.; Wu, Z.X.; Yang, Y.; Zhao, M.; Ma, X.Y.; Li, J.S.; Yang, D.H.; Chen, Z.S.; Fan, Y.F. Overexpression of ABCG2 confers resistance to pevonedistat, an NAE inhibitor. Exp. Cell Res. 2020, 388, 111858. [Google Scholar] [CrossRef]
- Kathawala, R.J.; Espitia, C.M.; Jones, T.M.; Islam, S.; Gupta, P.; Zhang, Y.K.; Chen, Z.S.; Carew, J.S.; Nawrocki, S.T. ABCG2 Overexpression Contributes to Pevonedistat Resistance. Cancers 2020, 12, 429. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hedberg, M.L.; Goh, G.; Chiosea, S.I.; Bauman, J.E.; Freilino, M.L.; Zeng, Y.; Wang, L.; Diergaarde, B.B.; Gooding, W.E.; Lui, V.W.; et al. Genetic landscape of metastatic and recurrent head and neck squamous cell carcinoma. J. Clin. Investig. 2016, 126, 169–180, Erratum in J. Clin. Investig. 2016, 126, 1606. [Google Scholar] [CrossRef] [Green Version]
- López-Arribillaga, E.; Rodilla, V.; Pellegrinet, L.; Guiu, J.; Iglesias, M.; Roman, A.C.; Gutarra, S.; González, S.; Muñoz-Cánoves, P.; Fernández-Salguero, P.; et al. Bmi1 regulates murine intestinal stem cell proliferation and self-renewal downstream of Notch. Development 2015, 142, 41–50. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Jian, X.; He, H.; Zhu, J.; Zhang, Q.; Zheng, Z.; Liang, X.; Chen, L.; Yang, M.; Peng, K.; Zhang, Z.; et al. Hsa_circ_001680 affects the proliferation and migration of CRC and mediates its chemoresistance by regulating BMI1 through miR-340. Mol. Cancer 2020, 19, 20. [Google Scholar] [CrossRef] [PubMed]





Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Li, Q.; Shen, Y.; Wu, S.; Wei, H.; Zou, J.; Xu, S.; Ling, Q.; Kang, M.; Huang, H.; Chen, X.; et al. MLN4924 Promotes Self-Renewal of Limbal Stem Cells and Ocular Surface Restoration. J. Pers. Med. 2023, 13, 379. https://doi.org/10.3390/jpm13030379
Li Q, Shen Y, Wu S, Wei H, Zou J, Xu S, Ling Q, Kang M, Huang H, Chen X, et al. MLN4924 Promotes Self-Renewal of Limbal Stem Cells and Ocular Surface Restoration. Journal of Personalized Medicine. 2023; 13(3):379. https://doi.org/10.3390/jpm13030379
Chicago/Turabian StyleLi, Qingjian, Yankun Shen, Shinan Wu, Hong Wei, Jie Zou, Sanhua Xu, Qian Ling, Min Kang, Hui Huang, Xu Chen, and et al. 2023. "MLN4924 Promotes Self-Renewal of Limbal Stem Cells and Ocular Surface Restoration" Journal of Personalized Medicine 13, no. 3: 379. https://doi.org/10.3390/jpm13030379