
Should Reward Deficiency Syndrome (RDS) Be Considered an Umbrella Disorder for Mental Illness and Associated Genetic and Epigenetic Induced Dysregulation of Brain Reward Circuitry?
 , , , , , and
, , , , , and
Abstract
:1. Introduction
2. RDS Criteria
3. RDS Is a Behavioral Octopus
4. Summary
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Gondré-Lewis, M.C.; Bassey, R.; Blum, K. Pre-clinical models of reward deficiency syndrome: A behavioral octopus. Neurosci. Biobehav. Rev. 2020, 115, 164–188. [Google Scholar] [CrossRef] [PubMed]
- Blum, K.; Oscar-Berman, M.; Demetrovics, Z.; Barh, D.; Gold, M.S. Genetic Addiction Risk Score (GARS): Molecular neurogenetic evidence for predisposition to Reward Deficiency Syndrome (RDS). Mol. Neurobiol. 2014, 50, 765–796. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Blum, K.; Bowirrat, A.; Braverman, E.R.; Baron, D.; Cadet, J.L.; Kazmi, S.; Elman, I.; Thanos, P.K.; Badgaiyan, R.D.; Downs, W.B.; et al. Reward Deficiency Syndrome (RDS): A Cytoarchitectural Common Neurobiological Trait of All Addictions. Int. J. Environ. Res. Public Health 2021, 18, 11529. [Google Scholar] [CrossRef] [PubMed]
- Blum, K.; Soni, D.; Badgaiyan, R.D.; Baron, D. Overcoming reward deficiency syndrome by the induction of “dopamine homeostasis” instead of opioids for addiction: Illusion or reality? J. Osteopat. Med. 2022; ahead of print. [Google Scholar] [CrossRef]
- Blum, K.; Baron, D.; McLaughlin, T.P.K.; Giordano, J.; Ceccanti, M.D.; Badgaiyan, R.D. Summary Document Research on RDS Anti-addiction Modelling: Annotated Bibliography. J. Syst. Integr. Neurosci. 2022, 8, 2–35. [Google Scholar] [CrossRef]
- Kotyuk, E.; Magi, A.; Eisinger, A.; Király, O.; Vereczkei, A.; Barta, C.; Griffiths, M.D.; Székely, A.; Kökönyei, G.; Farkas, J.; et al. Co-occurrences of substance use and other potentially addictive behaviors: Epidemiological results from the Psychological and Genetic Factors of the Addictive Behaviors (PGA) Study. J. Behav. Addict. 2020, 9, 272–288. [Google Scholar] [CrossRef]
- Fried, L.; Modestino, E.J.; Siwicki, D.; Lott, L.; Thanos, P.K.; Baron, D.; Badgaiyan, R.D.; Ponce, J.V.; Giordano, J.; Downs, W.B.; et al. Hypodopaminergia and “Precision Behavioral Management” (PBM): It is a Generational Family Affair. Curr. Pharm. Biotechnol. 2020, 21, 528–541. [Google Scholar] [CrossRef]
- Blum, K.; Steinberg, B.; Gondré-Lewis, M.C.; Baron, D.; Modestino, E.J.; Badgaiyan, R.D.; Downs, B.W.; Bagchi, D.; Brewer, R.; McLaughlin, T.; et al. A Review of DNA Risk Alleles to Determine Epigenetic Repair of mRNA Expression to Prove Therapeutic Effectiveness in Reward Deficiency Syndrome (RDS): Embracing “Precision Behavioral Management”. Psychol. Res. Behav. Manag. 2021, 14, 2115–2134. [Google Scholar] [CrossRef] [PubMed]
- O’Connell, L.A.; Hofmann, H.A. Genes, hormones, and circuits: An integrative approach to study the evolution of social behavior. Front. Neuroendocr. 2011, 32, 320–335. [Google Scholar] [CrossRef]
- Blum, K.; Gondré-Lewis, M.; Steinberg, B.; Elman, I.; Baron, D.; Modestino, E.J.; Badgaiyan, R.D.; Gold, M.S. Our evolved unique pleasure circuit makes humans different from apes: Reconsideration of data derived from animal studies. J. Syst. Integr. Neurosci. 2018, 4. [Google Scholar] [CrossRef]
- Blum, K.; Werner, T.; Carnes, S.; Carnes, P.; Bowirrat, A.; Giordano, J.; Oscar-Berman, M.; Gold, M. Sex, Drugs, and Rock ‘N’ Roll: Hypothesizing Common Mesolimbic Activation as a Function of Reward Gene Polymorphisms. J. Psychoact. Drugs 2012, 44, 38–55. [Google Scholar] [CrossRef]
- Blum, K.; Noble, E.P.; Sheridan, P.J.; Montgomery, A.; Ritchie, T.; Jagadeeswaran, P.; Nogami, H.; Briggs, A.H.; Cohn, J.B. Allelic association of human dopamine D2 receptor gene in alcoholism. JAMA 1990, 263, 2055–2060. [Google Scholar] [CrossRef]
- Noble, E.P.; Blum, K.; Ritchie, T.; Montgomery, A.; Sheridan, P.J. Allelic Association of the D2 Dopamine Receptor Gene with Receptor-Binding Characteristics in Alcoholism or Gene ism. Arch. Gen. Psychiatry 1991, 48, 648–654. [Google Scholar] [CrossRef]
- Caetano, R. Diffusion of an Idea: Jellinek’s Disease Concept in Latin America. Int. J. Addict. 1985, 20, 1621–1633. [Google Scholar] [CrossRef]
- Pickard, H. Is addiction a brain disease? A plea for agnosticism and heterogeneity. Psychopharmacology 2022, 239, 993–1007. [Google Scholar] [CrossRef]
- Blum, K.; Wood, R.C.; Braverman, E.R.; Chen, T.J.; Sheridan, P.J. The D2 dopamine receptor gene as a predictor of compulsive disease: Bayes’ theorem. Funct. Neurol. 1995, 10, 37–44. [Google Scholar]
- Lancaster, T.M.; Ihssen, N.; Brindley, L.M.; Linden, D.E.J. Further support for association between GWAS variant for positive emotion and reward systems. Transl. Psychiatry 2017, 7, e1018. [Google Scholar] [CrossRef] [Green Version]
- Levey, D.F.; Stein, M.B.; Wendt, F.R.; Pathak, G.A.; Zhou, H.; Aslan, M.; Quaden, R.; Harrington, K.M.; Nuñez, Y.Z.; Overstreet, C.; et al. Bi-ancestral depression GWAS in the Million Veteran Program and meta-analysis in >1.2 million individuals highlight new therapeutic directions. Nat. Neurosci. 2021, 24, 954–963. [Google Scholar] [CrossRef]
- Gupta, A.; Bowirrat, A.; Gomez, L.L.; Baron, D.; Elman, I.; Giordano, J.; Jalali, R.; Badgaiyan, R.D.; Modestino, E.J.; Gold, M.S.; et al. Hypothesizing in the Face of the Opioid Crisis Coupling Genetic Addiction Risk Severity (GARS) Testing with Electrotherapeutic Nonopioid Modalities Such as H-Wave Could Attenuate Both Pain and Hedonic Addictive Behaviors. Int. J. Environ. Res. Public Health 2022, 19, 552. [Google Scholar] [CrossRef]
- Elman, I.; Borsook, D. Common Brain Mechanisms of Chronic Pain and Addiction. Neuron 2016, 89, 11–36. [Google Scholar] [CrossRef] [Green Version]
- Gold, M.S.; Baron, D.; Bowirrat, A.; Blum, K. Neurological correlates of brain reward circuitry linked to opioid use disorder (OUD): Do homo sapiens acquire or have a reward deficiency syndrome? J. Neurol. Sci. 2020, 418, 117137. [Google Scholar] [CrossRef]
- Elman, I.; Borsook, D.; Lukas, S.E. Food Intake and Reward Mechanisms in Patients with Schizophrenia: Implications for Metabolic Disturbances and Treatment with Second-Generation Antipsychotic Agents. Neuropsychopharmacology 2006, 31, 2091–2120. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Thompson, J.; Thomas, N.; Singleton, A.; Piggott, M.; Lloyd, S.; Perry, E.K.; Morris, C.; Perry, R.H.; Ferrier, I.N.; A Court, J. D2 dopamine receptor gene (DRD2) Taql A polymorphism: Reduced dopamine D2 receptor binding in the human striatum associated with the A1 allele. Pharmacogenetics 1997, 7, 479–484. [Google Scholar] [CrossRef]
- Elman, I.; Borsook, D. Threat Response System: Parallel Brain Processes in Pain vis-à-vis Fear and Anxiety. Front. Psychiatry 2018, 9, 29. [Google Scholar] [CrossRef] [Green Version]
- LeDoux, J.E.; Pine, D.S. Using Neuroscience to Help Understand Fear and Anxiety: A Two-System Framework. Am. J. Psychiatry 2016, 173, 1083–1093. [Google Scholar] [CrossRef] [Green Version]
- Kotyuk, E.; Farkas, J.; Magi, A.; Eisinger, A.; Király, O.; Vereczkei, A.; Barta, C.; Griffiths, M.D.; Kökönyei, G.; Székely, A.; et al. The psychological and genetic factors of the addictive behaviors (PGA) study. Int. J. Methods Psychiatr. Res. 2019, 28, e1748. [Google Scholar] [CrossRef]
- Vereczkei, A.; Barta, C.; Magi, A.; Farkas, J.; Eisinger, A.; Király, O.; Belik, A.; Griffiths, M.D.; Szekely, A.; Sasvári-Székely, M.; et al. FOXN3 and GDNF Polymorphisms as Common Genetic Factors of Substance Use and Addictive Behaviors. J. Pers. Med. 2022, 12, 690. [Google Scholar] [CrossRef]
- Trachtenberg, M.C.; Blum, K. Alcohol and Opioid Peptides: Neuropharmacological Rationale for Physical Craving of Alcohol. Am. J. Drug Alcohol Abus. 1987, 13, 365–372. [Google Scholar] [CrossRef]
- Blum, K.; Gold, M.; Modestino, E.J.; Baron, D.; Boyett, B.; Siwicki, D.; Lott, L.; Podesta, A.; Roy, A.K.; Hauser, M.; et al. Would induction of dopamine homeostasis via coupling genetic addiction risk score (GARS®) and pro-dopamine regulation benefit benzodiazepine use disorder (BUD)? J. Syst. Integr. Neurosci. 2018, 3, 1000196. [Google Scholar] [CrossRef]
- Hollander, E. Obsessive–compulsive disorder and spectrum across the life span. Int. J. Psychiatry Clin. Pract. 2005, 9, 79–86. [Google Scholar] [CrossRef]
- Blum, K.; Sheridan, P.J.; Wood, R.C.; Braverman, E.R.; Chen, T.J.; Cull, J.G.; Comings, D.E. The D2 dopamine receptor gene as a determinant of reward deficiency syndrome. J. R. Soc. Med. 1996, 89, 396–400. [Google Scholar] [CrossRef]
- Jameel, S.; Shahnawaz, M.G.; Griffiths, M.D. Smartphone addiction in students: A qualitative examination of the components model of addiction using face-to-face interviews. J. Behav. Addict. 2019, 8, 780–793. [Google Scholar] [CrossRef] [PubMed]
- Antons, S.; Brand, M.; Potenza, M.N. Neurobiology of cue-reactivity, craving, and inhibitory control in non-substance addictive behaviors. J. Neurol. Sci. 2020, 415, 116952. [Google Scholar] [CrossRef] [PubMed]
- Majuri, J.; Joutsa, J.; Johansson, J.; Voon, V.; Alakurtti, K.; Parkkola, R.; Lahti, T.; Alho, H.; Hirvonen, J.; Arponen, E.; et al. Dopamine and Opioid Neurotransmission in Behavioral Addictions: A Comparative PET Study in Pathological Gambling and Binge Eating. Neuropsychopharmacology 2017, 42, 1169–1177. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hillemacher, T.; Frieling, H.; Buchholz, V.; Hussein, R.; Bleich, S.; Meyer, C.; John, U.; Bischof, A.; Rumpf, H.-J. Alterations in DNA-methylation of the dopamine-receptor 2 gene are associated with abstinence and health care utilization in individuals with a lifetime history of pathologic gambling. Prog. Neuro-Psychopharmacol. Biol. Psychiatry 2015, 63, 30–34. [Google Scholar] [CrossRef] [PubMed]
- Hillemacher, T.; Frieling, H.; Buchholz, V.; Hussein, R.; Bleich, S.; Meyer, C.; John, U.; Bischof, A.; Rumpf, H.-J. Dopamine-receptor 2 gene-methylation and gambling behavior in relation to impulsivity. Psychiatry Res. 2016, 239, 154–155. [Google Scholar] [CrossRef]
- Lee, H.K. Prerequisites for stakeholder framework: Consumer advocacy and health protection in the digital industry. J. Behav. Addict. 2020, 9, 898–902. [Google Scholar] [CrossRef]
- Agrawal, A.; Lynskey, M.T.; Hinrichs, A.; Grucza, R.; Saccone, S.F.; Krueger, R.; Neuman, R.; Howells, W.; Fisher, S.; Fox, L.; et al. A genome-wide association study of DSM-IV cannabis dependence. Addict. Biol. 2011, 16, 514–518. [Google Scholar] [CrossRef] [Green Version]
- Chen, T.J.; Blum, K.; Mathews, D.; Fisher, L.; Schnautz, N.; Braverman, E.R.; Schoolfield, J.; Downs, B.W.; Comings, D.E. Are dopaminergic genes involved in a predisposition to pathological aggression?: Hypothesizing the importance of “super normal controls” in psychiatricgenetic research of complex behavioral disorders. Med. Hypotheses 2005, 65, 703–707. [Google Scholar] [CrossRef]
- Al-Eitan, L.N.; Rababa’H, D.M.; Alghamdi, M.A. Genetic susceptibility of opioid receptor genes polymorphism to drug addiction: A candidate-gene association study. BMC Psychiatry 2021, 21, 5. [Google Scholar] [CrossRef]
- Gowin, J.L.; May, A.C.; Wittmann, M.; Tapert, S.F.; Paulus, M. Doubling Down: Increased Risk-Taking Behavior Following a Loss by Individuals with Cocaine Use Disorder Is Associated with Striatal and Anterior Cingulate Dysfunction. Biol. Psychiatry Cogn. Neurosci. Neuroimaging 2017, 2, 94–103. [Google Scholar] [CrossRef]
- Cooper, S.; Robison, A.; Mazei-Robison, M.S. Reward Circuitry in Addiction. Neurotherapeutics 2017, 14, 687–697. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Pourhamzeh, M.; Moravej, F.G.; Arabi, M.; Shahriari, E.; Mehrabi, S.; Ward, R.; Ahadi, R.; Joghataei, M.T. The Roles of Serotonin in Neuropsychiatric Disorders. Cell. Mol. Neurobiol. 2021, 42, 1671–1692. [Google Scholar] [CrossRef] [PubMed]
- Bara, A.; Ferland, J.-M.N.; Rompala, G.; Szutorisz, H.; Hurd, Y.L. Cannabis and synaptic reprogramming of the developing brain. Nat. Rev. Neurosci. 2021, 22, 423–438. [Google Scholar] [CrossRef]
- Blum, K.; McLaughlin, T.; Bowirrat, A.; Modestino, E.J.; Baron, D.; Gomez, L.L.; Ceccanti, M.; Braverman, E.R.; Thanos, P.K.; Cadet, J.L.; et al. Reward Deficiency Syndrome (RDS) Surprisingly Is Evolutionary and Found Everywhere: Is It “Blowin’ in the Wind”? J. Pers. Med. 2022, 12, 321. [Google Scholar] [CrossRef]
- Blum, K.; Febo, M.; Badgaiyan, R.D.; Baron, D.; Thanos, P.K.; Colon-Perez, L.M.; Demotrovics, Z.; Gold, M.S. Dopamine homeostasis brain functional connectivity in reward deficiency syndrome. Front. Biosci. 2017, 22, 669–691. [Google Scholar] [CrossRef] [Green Version]
- Blum, K.; Febo, M.; Badgaiyan, R.D.; Braverman, E.R.; Dushaj, K.; Li, M.; Demetrovics, Z. Neuronutrient amino-acid therapy protects against reward deficiency syndrome: Dopaminergic key to homeostasis and neuroplasticity. Curr. Pharm. Des. 2016, 22, 5837–5854. [Google Scholar] [CrossRef] [Green Version]
- Blum, K.; Gold, M.S.; Jacobs, W.; McCall, W.V.; Febo, M.; Baron, D.; Dushaj, K.; Demetrovics, Z.; Badgaiyan, R.D. Neurogenetics of acute and chronic opiate opioid abstinence treating symptoms and the cause. Front. Biosci. 2017, 22, 1247–1288. [Google Scholar] [CrossRef] [Green Version]
- Blum, K.; Febo, M.; Fried, L.; Li, M.; Dushaj, K.; Braverman, E.R.; McLaughlin, T.; Steinberg, B.; Badgaiyan, R.D. Hypothesizing That Neuropharmacological and Neuroimaging Studies of Glutaminergic-Dopaminergic Optimization Complex (KB220Z) Are Associated With “Dopamine Homeostasis” in Reward Deficiency Syndrome (RDS). Subst. Use Misuse 2017, 52, 535–547. [Google Scholar] [CrossRef] [Green Version]
- Miller, D.K.; Bowirrat, A.; Manka, M.; Miller, M.; Stokes, S.; Manka, D.; Allen, C.; Gant, C.; Downs, B.W.; Smolen, A.; et al. Acute Intravenous Synaptamine Complex Variant KB220™ “Normalizes” Neurological Dysregulation in Patients during Protracted Abstinence from Alcohol and Opiates as Observed Using Quantitative Electroencephalographic and Genetic Analysis for Reward Polymorphisms: Part 1, Pilot Study with 2 Case Reports. Postgrad. Med. 2010, 122, 188–213. [Google Scholar] [CrossRef]
- Miller, M.; Chen, A.L.; Stokes, S.D.; Silverman, S.; Bowirrat, A.; Manka, M.; Manka, D.; Miller, D.K.; Perrine, K.; Chen, T.J.; et al. Early Intervention of Intravenous KB220IV- Neuroadaptagen Amino-Acid Therapy (NAAT)™ Improves Behavioral Outcomes in a Residential Addiction Treatment Program: A Pilot Study. J. Psychoact. Drugs 2012, 44, 398–409. [Google Scholar] [CrossRef]
- Blum, K.; Modestino, E.J.; Gondre-Lewis, M.C.; Baron, K.; Steinberg, B.; Thanos, K.P.; Downs, W.B.; Siwicki, D.; Lott, L.; Braverman, R.E.; et al. Pro-Dopamine Regulator (KB220) A Fifty Year Sojourn to Combat Reward Deficiency Syndrome (RDS): Evidence Based Bibliography (Annotated). CPQ Neurol. Psychol. 2018, 1. Available online: https://www.cientperiodique.com/journal/fulltext/CPQNP/1/2/13 (accessed on 13 October 2022).
- Febo, M.; Blum, K.; Badgaiyan, R.D.; Perez, P.D.; Colon-Perez, L.M.; Thanos, P.K.; Ferris, C.F.; Kulkarni, P.; Giordano, J.; Baron, D.; et al. Enhanced functional connectivity and volume between cognitive and reward centers of naïve rodent brain produced by pro-dopaminergic agent KB220Z. PLoS ONE 2017, 12, e0174774. [Google Scholar] [CrossRef] [Green Version]
- Blum, K.; Liu, Y.; Wang, W.; Wang, Y.; Zhang, Y.; Oscar-Berman, M.; Smolen, A.; Febo, M.; Han, D.; Simpatico, T.; et al. rsfMRI effects of KB220Z™ on neural pathways in reward circuitry of abstinent genotyped heroin addicts. Postgrad. Med. 2015, 127, 232–241. [Google Scholar] [CrossRef] [PubMed]
- Blum, K.; Chen, T.J.; Morse, S.; Giordano, J.; Chen, A.L.; Thompson, J.; Allen, C.; Smolen, A.; Lubar, J.; Stice, E.; et al. Overcoming qEEG abnormalities and reward gene deficits during protracted abstinence in male psychostimulant and polydrug abusers utilizing putative dopamine D2 agonist. Postgrad. Med. 2010, 122, 214–226. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Willuhn, I.; Burgeno, L.; Groblewski, P.A.; Phillips, P.E.M. Excessive cocaine use results from decreased phasic dopamine signaling in the striatum. Nat. Neurosci. 2014, 17, 704–709. [Google Scholar] [CrossRef] [PubMed]
- Park, K.; Volkow, N.D.; Pan, Y.; Du, C. Chronic cocaine dampens dopamine signaling during cocaine intoxication and unbalances D1 over D2 receptor signaling. J. Neurosci. 2013, 33, 15827–15836. [Google Scholar] [CrossRef] [Green Version]
- Thanawala, V.J.K.A.R.G.V.; Kadam, V.J.; Ghosh, R. Enkephalinase Inhibitors: Potential Agents for the Management of Pain. Curr. Drug Targets 2008, 9, 887–894. [Google Scholar] [CrossRef] [PubMed]
- Ramírez-Sánchez, M.; Prieto, I.; Segarra, A.-B.; Martínez-Cañamero, M.; Banegas, I.; de Gasparo, M. Enkephalinase regulation. Vitam. Horm. 2019, 111, 105–129. [Google Scholar] [CrossRef]
- Kitade, T.; Odahara, Y.; Shinohara, S.; Ikeuchi, T.; Sakai, T.; Morikawa, K.; Minamikawa, M.; Toyota, S.; Kawachi, A.; Hyodo, M.; et al. Studies on the enhanced effect of acupuncture analgesia and acupuncture anesthesia by d-phenylalanine (2nd report)—Schedule of administration and clinical effects in low back pain and tooth extraction. Acupunct. Electro. Ther. Res. 1990, 15, 121–135. [Google Scholar] [CrossRef] [PubMed]
- Cheng, R.S.; Pomeranz, B. A combined treatment with d-amino acids and electroacupuncture produces a greater analgesia than either treatment alone; naloxone reverses these effects. Pain 1980, 8, 231–236. [Google Scholar] [CrossRef]
- Blum, K.; Baron, D.; McLaughlin, T.; Gold, M.S. Molecular neurological correlates of endorphinergic/dopaminergic mechanisms in reward circuitry linked to endorphinergic deficiency syndrome (EDS). J. Neurol. Sci. 2020, 411, 116733. [Google Scholar] [CrossRef]
- Blum, K.; Modestino, E.J.; Gondré-Lewis, M.; Downs, B.W.; Baron, D.; Steinberg, B.; Siwicki, D.; Giordano, J.; McLaughlin, T.; Neary, J.; et al. “Dopamine homeostasis” requires balanced polypharmacy: Issue with destructive, powerful dopamine agents to combat America’s drug epidemic. J. Syst. Integr. Neurosci. 2017, 3. [Google Scholar] [CrossRef] [Green Version]
- Blum, K.; Briggs, A.H.; Elston, S.F.A.; DeLallo, L.; Sheridan, P.J.; Sar, M. Reduced Leucine-Enkephalin—Like Immunoreactive Substance in Hamster Basal Ganglia After Long-Term Ethanol Exposure. Science 1982, 216, 1425–1427. [Google Scholar] [CrossRef] [PubMed]
- Blum, K.; Elston, S.F.; DeLallo, L.; Briggs, A.H.; Wallace, J.E. Ethanol acceptance as a function of genotype amounts of brain [Met]enkephalin. Proc. Natl. Acad. Sci. USA 1983, 80, 6510–6512. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Blum, K.; Briggs, A.H.; Trachtenberg, M.C.; Delallo, L.; Wallace, J.E. Enkephalinase inhibition: Regulation of ethanol intake in genetically predisposed mice. Alcohol 1987, 4, 449–456. [Google Scholar] [CrossRef]
- Blum, K.; Trachtenberg, M.C.; Elliott, C.E.; Dingler, M.L.; Sexton, R.L.; Samuels, A.I.; Cataldie, L. Enkephalinase inhibition and precursor amino acid loading improves inpatient treatment of alcohol and polydrug abusers: Double-blind placebo-controlled study of the nutritional adjunct SAAVE. Alcohol 1988, 5, 481–493. [Google Scholar] [CrossRef]
- Brown, R.J.; Blum, K.; Trachtenberg, M.C. Neurodynamics of Relapse Prevention: A Neuronutrient Approach to Outpatient DUI Offenders. J. Psychoact. Drugs 1990, 22, 173–187. [Google Scholar] [CrossRef] [PubMed]
- Casey, B.J.; Craddock, N.; Cuthbert, B.N.; Hyman, S.E.; Lee, F.S.; Ressler, K.J. DSM-5 and RDoC: Progress in psychiatry research? Nat. Rev. Neurosci. 2013, 14, 810–814. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kótyuk, E.; Urbán, R.; Hende, B.; Richman, M.; Magi, A.; Király, O.; Barta, C.; Griffiths, M.D.; Potenza, M.N.; Badgaiyan, R.D.; et al. Development and validation of the Reward Deficiency Syndrome Questionnaire (RDSQ-29). J. Psychopharmacol. 2022, 36, 409–422. [Google Scholar] [CrossRef]
- Blum, K.; McLaughlin, T.; Modestino, E.J.; Baron, D.; Bowirrat, A.; Brewer, R.; Steinberg, B.; Roy, A.K.; Febo, M.; Badgaiyan, R.D.; et al. Epigenetic Repair of Terrifying Lucid Dreams by Enhanced Brain Reward Functional Connectivity and Induction of Dopaminergic Homeo—static Signaling. Curr. Psychopharmacol. 2021, 10, 170–180. [Google Scholar] [CrossRef] [PubMed]
- Blum, K.; Bowirrat, A.; Lewis, M.C.; Simpatico, T.A.; Ceccanti, M.; Steinberg, B.; Modestino, E.J.; Thanos, P.K.; Baron, D.; McLaughlin, T.; et al. Exploration of Epigenetic State Hyperdopaminergia (Surfeit) and Genetic Trait Hypodopaminergia (Deficit) during Adolescent Brain Development. Curr. Psychopharmacol. 2021, 10, 181–196. [Google Scholar] [CrossRef]
- Blum, K.; Bowirrat, A.; Gomez, L.L.; Downs, B.W.; Bagchi, D.; Barh, D.; Modestino, E.J.; Baron, D.; McLaughlin, T.; Thanos, P.; et al. Why haven’t we solved the addiction crisis? J. Neurol. Sci. 2022, 442, 120404. [Google Scholar] [CrossRef] [PubMed]
- Blum, K.; Febo, M.; Smith, D.E.; Roy, A.K., 3rd; Demetrovics, Z.; Cronjé, F.J.; Femino, J.; Agan, G.; Fratantonio, J.L.; Pandey, S.C.; et al. Neurogenetic and epigenetic correlates of adolescent predisposition to and risk for addictive behaviors as a function of prefrontal cortex dysregulation. J. Child Adolesc. Psychopharmacol. 2015, 25, 286–292. [Google Scholar] [CrossRef] [PubMed]



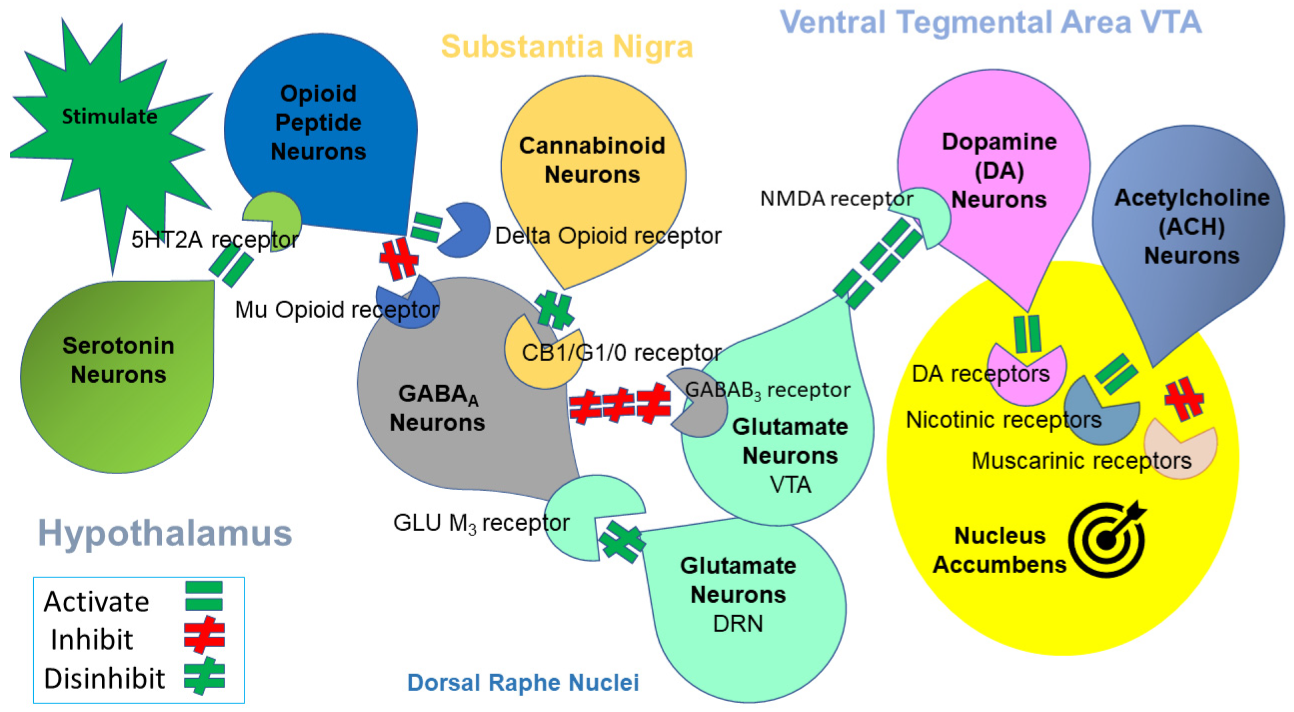{kind=link}
{kind=link}
| GARS Score of 4 | With 1–2 Criteria | Mild |
|---|---|---|
| GARS Score of 4–6 | with 3 or more Criteria | Moderate |
| GARS Score of 7–9 | with 1–2 Criteria | Moderate |
| GARS Score of 7–9 | with 3 or more Criteria | Severe |
| GARS Score of 10 or more | with 1–2 Criteria | Severe |
| GARS Score of 10 or more | with 3 or more Criteria | Profound |
| Criteria Set ONE—DSM-5 Disorders | ||
| A present diagnosis, past diagnosis, or history of these behavioral disorders | ||
| Substance Use Process Disorders | Disorders: Alcohol Use Disorder, Opioid Use Disorder, Cannabis Use Disorder; Sedative, Hypnotic, Anxiolytic Use Disorder; Cocaine Use Disorder, Amphetamine Use Disorder, Hallucinogen Use Disorder, Nicotine Use Disorder, Inhalant Use Disorder, Other, Unknown Substance Use Disorder | |
| Specifiers: Mild, Moderate, Severe, Early Remission (6–12 months), Sustained Remission (12+ months), in a Controlled Environment, on Maintenance Therapy | ||
| Process Disorders | Gambling, Sex, Other Specified Process Disorders | |
| Depressive (and related) Disorders | Major Depression, Dysthymia, Disruptive Mood Dysregulation, SUD/Medication/Medical Condition Induced Depressive Disorder, Disruptive Premenstrual Dysphoric Disorder | |
| Anxiety Disorders | Generalized Anxiety Disorder, Social Anxiety, Panic Attack | |
| Disorder, Separation Anxiety, Selective Mutism, Specific Phobia, | ||
| SUD/Medication/Medical Condition Induced Anxiety | ||
| Trauma and Stress Disorders | Reactive Attachment, Disinhibited Social Engagement, Post-Traumatic Stress Disorder (PTSD), Acute Stress Disorders | |
| Disruptive, Impulse Control, and Conduct Disorders | Oppositional Defiant Disorder, Intermittent Explosive Disorder, Conduct Disorder, Pyromania, Kleptomania | |
| Personality Disorders | General Personality Disorder, Paranoid Personality Disorder, Schizoid/Schizotypal Personality Disorder, Anti-Social Personality Disorder, Borderline Personality Disorder, Histrionic Personality Disorder, Narcissistic, Personality Disorder, Avoidant Personality Disorder, Dependent Personality Disorder | |
| Obsessive Compulsive Disorders and Related Disorders | Trichotillomania, Excoriation Disorder, SUD/Medical/Medication Induced Obsessive Compulsive Disorder (OCD), other Medical Condition, Induced Personality Disorder | |
| Schizophrenic Disorders | Schizophrenic Disorders: Schizophrenia, Schizoaffective Disorder, Schizophreniform Disorder, Delusional Disorder, Brief Psychotic Disorder, MH/Medical Catalonia, SUD/Medication/Medical Condition Induced Psychotic Disorder | |
| Dissociative Disorders | Dissociative Identity Disorder, Dissociative Amnesia, Depersonalization/Derealization Disorder | |
| Other Not Otherwise Specified (NOS) Disorders | Gender Dysphoric Disorder | |
| Paraphilic Disorders | ||
| Spectrum Disorders | Attention Deficient Disorder, Attention Deficient/Hyperactivity Disorder, Tourette’s Syndrome Autism | |
| Criteria Set TWO | ||
| Reported history of these symptoms: | ||
| Novelty seeking | This trait is associated with exploratory activity in response to novel stimulation, impulsive decision making, extravagance in approach to reward cues, quick loss of temper, and avoidance of frustration. | |
| Impulsivity | The construct of impulsivity includes at least two independent components: first, acting without an appropriate amount of deliberation, which may or may not be functional; and second, choosing short-term gains over long-term ones. | |
| Difficulty feeling reward (Anhedonia) | Either a reduced ability to experience pleasure or a diminished interest in engaging in pleasurable activities. | |
| Motivational Anhedonia | Decrease in motivation to participate in pleasurable activities | |
| Rumination, Obsessive, and Intrusive Negative Thoughts | Possible causes and consequences, as opposed to its solutions | |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Blum, K.; Dennen, C.A.; Elman, I.; Bowirrat, A.; Thanos, P.K.; Badgaiyan, R.D.; Downs, B.W.; Bagchi, D.; Baron, D.; Braverman, E.R.; et al. Should Reward Deficiency Syndrome (RDS) Be Considered an Umbrella Disorder for Mental Illness and Associated Genetic and Epigenetic Induced Dysregulation of Brain Reward Circuitry? J. Pers. Med. 2022, 12, 1719. https://doi.org/10.3390/jpm12101719
Blum K, Dennen CA, Elman I, Bowirrat A, Thanos PK, Badgaiyan RD, Downs BW, Bagchi D, Baron D, Braverman ER, et al. Should Reward Deficiency Syndrome (RDS) Be Considered an Umbrella Disorder for Mental Illness and Associated Genetic and Epigenetic Induced Dysregulation of Brain Reward Circuitry? Journal of Personalized Medicine. 2022; 12(10):1719. https://doi.org/10.3390/jpm12101719
Chicago/Turabian StyleBlum, Kenneth, Catherine A. Dennen, Igor Elman, Abdalla Bowirrat, Panayotis K. Thanos, Rajendra D. Badgaiyan, B. William Downs, Debasis Bagchi, David Baron, Eric R. Braverman, and et al. 2022. "Should Reward Deficiency Syndrome (RDS) Be Considered an Umbrella Disorder for Mental Illness and Associated Genetic and Epigenetic Induced Dysregulation of Brain Reward Circuitry?" Journal of Personalized Medicine 12, no. 10: 1719. https://doi.org/10.3390/jpm12101719