
Genetic Addiction Risk and Psychological Profiling Analyses for “Preaddiction” Severity Index

 , , , , , , , ,
, , , , , , , ,
Abstract
:1. Background
2. Reward Deficiency Syndrome Index (RDSI) and Genetics
3. Understanding GARS
4. Initial DNA Customized Studies
5. Can Early Genetic Risk Assessment of “Preaddiction” like “Prediabetes” Provide the Missing Piece to Help Overcome Substance Use Disorder?
6. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Oesterle, T.S.; Thusius, N.J.; Rummans, T.A.; Gold, M.S. Medication-Assisted Treatment for Opioid-Use Disorder. Mayo Clin. Proc. 2019, 94, 2072–2086. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Blum, K.; Baron, D.; McLaughlin, T.; Gold, M.S. Molecular neurological correlates of endorphinergic/dopaminergic mechanisms in reward circuitry linked to endorphinergic deficiency syndrome (EDS). J. Neurol. Sci. 2020, 411, 116733. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Patterson Silver Wolf, D.A.; Gold, M. Treatment resistant opioid use disorder (TROUD): Definition, rationale, and recommendations. J. Neurol. Sci. 2020, 411, 116718. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Gold, M.S.; Baron, D.; Bowirrat, A.; Blum, K. Neurological correlates of brain reward circuitry linked to opioid use disorder (OUD): Do homo sapiens acquire or have a reward deficiency syndrome? J. Neurol. Sci. 2020, 418, 117137. [Google Scholar] [CrossRef] [PubMed]
- Downs, B.W.; Blum, K.; Baron, D.; Bowirrat, A.; Lott, L.; Brewer, R.; Boyett, B.; Siwicki, D.; Roy, A.K.; Podesta, A.; et al. Death by Opioids: Are there non-addictive scientific solutions? J. Syst. Integr. Neurosci. 2019, 5. [Google Scholar] [CrossRef]
- Blum, K.; Lott, L.; Baron, D.; Smith, D.E.; Badgaiyan, R.D.; Gold, M.S. Improving naltrexone compliance and outcomes with putative pro- dopamine regulator KB220, compared to treatment as usual. J. Syst. Integr. Neurosci. 2020, 7. [Google Scholar] [CrossRef]
- Morgan, J.R.; Schackman, B.R.; Leff, J.A.; Linas, B.P.; Walley, A.Y. Injectable naltrexone, oral naltrexone, and buprenorphine utilization and discontinuation among individuals treated for opioid use disorder in a United States commercially insured population. J. Subst. Abus. Treat. 2018, 85, 90–96. [Google Scholar] [CrossRef]
- Ooteman, W.; Naassila, M.; Koeter, M.W.; Verheul, R.; Schippers, G.M.; Houchi, H.; Daoust, M.; van den Brink, W. Predicting the effect of naltrexone and acamprosate in alcohol-dependent patients using genetic indicators. Addict. Biol. 2009, 14, 328–337. [Google Scholar] [CrossRef]
- Cowen, M.S.; Adams, C.; Kraehenbuehl, T.; Vengeliene, V.; Lawrence, A.J. The acute anti-craving effect of acamprosate in alcohol-preferring rats is associated with modulation of the mesolimbic dopamine system. Addict. Biol. 2005, 10, 233–242. [Google Scholar] [CrossRef]
- Gold, M.S.; Blum, K.; Febo, M.; Baron, D.; Modestino, E.J.; Elman, I.; Badgaiyan, R.D. Molecular role of dopamine in anhedonia linked to reward deficiency syndrome (RDS) and anti- reward systems. Front. Biosci. 2018, 10, 309–325. [Google Scholar] [CrossRef]
- Blum, K.; Thanos, P.K.; Wang, G.J.; Febo, M.; Demetrovics, Z.; Modestino, E.J.; Braverman, E.R.; Baron, D.; Badgaiyan, R.D.; Gold, M.S. The Food and Drug Addiction Epidemic: Targeting Dopamine Homeostasis. Curr. Pharm. Des. 2018, 23, 6050–6061. [Google Scholar] [CrossRef]
- Blum, K.; Madigan, M.A.; Fried, L.; Braverman, E.R.; Giordano, J.; Badgaiyan, R.D. Coupling genetic addiction risk score (GARS) and pro dopamine regulation (KB220) to combat substance use disorder (SUD). J. Addict. Rehabil. Med. 2017, 1, 26–33. [Google Scholar] [CrossRef]
- Vereczkei, A.; Barta, C.; Magi, A.; Farkas, J.; Eisinger, A.; Király, O.; Belik, A.; Griffiths, M.D.; Szekely, A.; Sasvári-Székely, M.; et al. FOXN3 and GDNF Polymorphisms as Common Genetic Factors of Substance Use and Addictive Behaviors. J. Pers. Med. 2022, 12, 690. [Google Scholar] [CrossRef]
- Blum, K.; Briggs, A.H.; Elston, S.F.; DeLallo, L. Psychogenetics of drug seeking behavior. Subst. Alcohol Actions Misuse 1980, 1, 255–257. [Google Scholar]
- Madigan, M.A.; Gupta, A.; Bowirrat, A.; Baron, D.; Badgaiyan, R.D.; Elman, I.; Dennen, C.A.; Braverman, E.R.; Gold, M.S.; Blum, K. Precision Behavioral Management (PBM) and Cognitive Control as a Potential Therapeutic and Prophylactic Modality for Reward Deficiency Syndrome (RDS): Is There Enough Evidence? Int. J. Environ. Res. Public Health 2022, 19, 6395. [Google Scholar] [CrossRef]
- Krebs, G.; Heyman, I. Obsessive-compulsive disorder in children and adolescents. Arch. Dis. Child. 2015, 100, 495–499. [Google Scholar] [CrossRef] [Green Version]
- Blum, K.; Sheridan, P.J.; Wood, R.C.; Braverman, E.R.; Chen, T.J.; Cull, J.G.; Comings, D.E. The D2 dopamine receptor gene as a determinant of reward deficiency syndrome. J. R. Soc. Med. 1996, 89, 396–400. [Google Scholar] [CrossRef] [Green Version]
- Jameel, S.; Shahnawaz, M.G.; Griffiths, M.D. Smartphone addiction in students: A qualitative examination of the components model of addiction using face-to-face interviews. J. Behav. Addict. 2019, 8, 780–793. [Google Scholar] [CrossRef]
- Cunningham-Williams, R.M.; Gattis, M.N.; Dore, P.M.; Shi, P.; Spitznagel, E.L., Jr. Towards DSM-V: Considering other withdrawal-like symptoms of pathological gambling disorder. Int. J. Methods Psychiatr. Res. 2009, 18, 13–22. [Google Scholar] [CrossRef]
- Romanczuk-Seiferth, N.; van den Brink, W.; Goudriaan, A.E. From symptoms to neurobiology: Pathological gambling in the light of the new classification in DSM-5. Neuropsychobiology 2014, 70, 95–102. [Google Scholar] [CrossRef]
- Orford, J.; Morison, V.; Somers, M. Drinking and gambling: A comparison with implications for theories of addiction. Drug Alcohol Rev. 1996, 15, 47–56. [Google Scholar] [CrossRef] [PubMed]
- Grant, J.E.; Chamberlain, S.R. Expanding the definition of addiction: DSM-5 vs. ICD-11. CNS Spectr. 2016, 21, 300–303. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Brick, L.A.; Keller, M.C.; Knopik, V.S.; McGeary, J.E.; Palmer, R.H.C. Shared additive genetic variation for alcohol dependence among subjects of African and European ancestry. Addict. Biol. 2019, 24, 132–144. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Blum, K.; Wood, R.C.; Braverman, E.R.; Chen, T.J.; Sheridan, P.J. The D2 dopamine receptor gene as a predictor of compulsive disease: Bayes’ theorem. Funct. Neurol. 1995, 10, 37–44. [Google Scholar] [PubMed]
- Blum, K.; McLaughlin, T.; Bowirrat, A.; Modestino, E.J.; Baron, D.; Gomez, L.L.; Ceccanti, M.; Braverman, E.R.; Thanos, P.K.; Cadet, J.L.; et al. Reward Deficiency Syndrome (RDS) Surprisingly Is Evolutionary and Found Everywhere: Is It “Blowin’ in the Wind”? J. Pers. Med. 2022, 12, 321. [Google Scholar] [CrossRef]
- Berridge, K.C.; Robinson, T.E. Liking, wanting, and the incentive-sensitization theory of addiction. Am. Psychol. 2016, 71, 670–679. [Google Scholar] [CrossRef]
- Kibaly, C.; Alderete, J.A.; Liu, S.H.; Nasef, H.S.; Law, P.Y.; Evans, C.J.; Cahill, C.M. Oxycodone in the Opioid Epidemic: High ‘Liking’, ‘Wanting’, and Abuse Liability. Cell. Mol. Neurobiol. 2021, 41, 899–926. [Google Scholar] [CrossRef]
- Blum, K.; Gardner, E.; Oscar-Berman, M.; Gold, M. “Liking” and “wanting” linked to Reward Deficiency Syndrome (RDS): Hypothesizing differential responsivity in brain reward circuitry. Curr. Pharm. Des. 2012, 18, 113–118. [Google Scholar] [CrossRef] [Green Version]
- File, D.; Bőthe, B.; File, B.; Demetrovics, Z. The Role of Impulsivity and Reward Deficiency in “Liking” and “Wanting” of Potentially Problematic Behaviors and Substance Uses. Front. Psychiatry 2022, 13, 820836. [Google Scholar] [CrossRef]
- Cox, D.J.; Dolan, S.B.; Johnson, P.; Johnson, M.W. Delay and probability discounting in cocaine use disorder: Comprehensive examination of money, cocaine, and health outcomes using gains and losses at multiple magnitudes. Exp. Clin. Psychopharmacol. 2020, 28, 724–738. [Google Scholar] [CrossRef]
- Ferrari, C.; Lega, C.; Tamietto, M.; Nadal, M.; Cattaneo, Z. I find you more attractive … after (prefrontal cortex) stimulation. Neuropsychologia 2015, 72, 87–93. [Google Scholar] [CrossRef]
- Lindgren, E.; Gray, K.; Miller, G.; Tyler, R.; Wiers, C.E.; Volkow, N.D.; Wang, G.J. Food addiction: A common neurobiological mechanism with drug abuse. Front. Biosci. 2018, 23, 811–836. [Google Scholar] [CrossRef] [Green Version]
- Müller, C.P. Serotonin and consciousness—A reappraisal. Behav. Brain Res. 2022, 432, 113970. [Google Scholar] [CrossRef]
- Vanyukov, M.M.; Tarter, R.E.; Kirillova, G.P.; Kirisci, L.; Reynolds, M.D.; Kreek, M.J.; Conway, K.P.; Maher, B.S.; Iacono, W.G.; Bierut, L.; et al. Common liability to addiction and “gateway hypothesis”: Theoretical, empirical and evolutionary perspective. Drug Alcohol Depend. 2012, 123 (Suppl. S1), S3–S17. [Google Scholar] [CrossRef] [Green Version]
- Barnett, E.J.; Biederman, J.; Doyle, A.E.; Hess, J.; DiSalvo, M.; Faraone, S.V. Identifying Pediatric Mood Disorders From Transdiagnostic Polygenic Risk Scores: A Study of Children and Adolescents. J. Clin. Psychiatry 2022, 83, 21m14180. [Google Scholar] [CrossRef]
- Ferland, J.N.; Hynes, T.J.; Hounjet, C.D.; Lindenbach, D.; Vonder Haar, C.; Adams, W.K.; Phillips, A.G.; Winstanley, C.A. Prior Exposure to Salient Win-Paired Cues in a Rat Gambling Task Increases Sensitivity to Cocaine Self-Administration and Suppresses Dopamine Efflux in Nucleus Accumbens: Support for the Reward Deficiency Hypothesis of Addiction. J. Neurosci. 2019, 39, 1842–1854. [Google Scholar] [CrossRef] [Green Version]
- Blum, K.; Bowirrat, A.; Braverman, E.R.; Baron, D.; Cadet, J.L.; Kazmi, S.; Elman, I.; Thanos, P.K.; Badgaiyan, R.D.; Downs, W.B.; et al. Reward Deficiency Syndrome (RDS): A Cytoarchitectural Common Neurobiological Trait of All Addictions. Int. J. Environ. Res. Public Health 2021, 18, 11529. [Google Scholar] [CrossRef]
- Gardner, E.L. Addiction and brain reward and antireward pathways. Adv. Psychosom. Med. 2011, 30, 22–60. [Google Scholar] [CrossRef] [Green Version]
- Kotyuk, E.; Magi, A.; Eisinger, A.; Király, O.; Vereczkei, A.; Barta, C.; Griffiths, M.D.; Székely, A.; Kökönyei, G.; Farkas, J.; et al. Co-occurrences of substance use and other potentially addictive behaviors: Epidemiological results from the Psychological and Genetic Factors of the Addictive Behaviors (PGA) Study. J. Behav. Addict. 2020, 9, 272–288. [Google Scholar] [CrossRef]
- Karlsson Linnér, R.; Mallard, T.T.; Barr, P.B.; Sanchez-Roige, S.; Madole, J.W.; Driver, M.N.; Poore, H.E.; de Vlaming, R.; Grotzinger, A.D.; Tielbeek, J.J.; et al. Multivariate analysis of 1.5 million people identifies genetic associations with traits related to self-regulation and addiction. Nat. Neurosci. 2021, 24, 1367–1376. [Google Scholar] [CrossRef]
- Uhl, G.R.; Drgon, T.; Johnson, C.; Liu, Q.R. Addiction genetics and pleiotropic effects of common haplotypes that make polygenic contributions to vulnerability to substance dependence. J. Neurogenet. 2009, 23, 272–282. [Google Scholar] [CrossRef] [PubMed]
- Sheerin, C.M.; Bountress, K.E.; Meyers, J.L.; Saenz de Viteri, S.S.; Shen, H.; Maihofer, A.X.; Duncan, L.E.; Amstadter, A.B. Shared molecular genetic risk of alcohol dependence and posttraumatic stress disorder (PTSD). Psychol. Addict. Behav. 2020, 34, 613–619. [Google Scholar] [CrossRef] [PubMed]
- Wetherill, L.; Lai, D.; Johnson, E.C.; Anokhin, A.; Bauer, L.; Bucholz, K.K.; Dick, D.M.; Hariri, A.R.; Hesselbrock, V.; Kamarajan, C.; et al. Genome-wide association study identifies loci associated with liability to alcohol and drug dependence that is associated with variability in reward-related ventral striatum activity in African- and European-Americans. Genes Brain Behav. 2019, 18, e12580. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Reyes-Gibby, C.C.; Yuan, C.; Wang, J.; Yeung, S.C.; Shete, S. Gene network analysis shows immune-signaling and ERK1/2 as novel genetic markers for multiple addiction phenotypes: Alcohol, smoking and opioid addiction. BMC Syst. Biol. 2015, 9, 25. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Weinstein, A.; Weinstein, Y. Exercise addiction- diagnosis, bio-psychological mechanisms and treatment issues. Curr. Pharm. Des. 2014, 20, 4062–4069. [Google Scholar] [CrossRef]
- Müller, A.; Loeber, S.; Söchtig, J.; Te Wildt, B.; De Zwaan, M. Risk for exercise dependence, eating disorder pathology, alcohol use disorder and addictive behaviors among clients of fitness centers. J. Behav. Addict. 2015, 4, 273–280. [Google Scholar] [CrossRef] [Green Version]
- Yau, Y.H.; Potenza, M.N. Gambling disorder and other behavioral addictions: Recognition and treatment. Harv. Rev. Psychiatry 2015, 23, 134–146. [Google Scholar] [CrossRef]
- Caldeira, K.M.; Arria, A.M.; O’Grady, K.E.; Vincent, K.B.; Robertson, C.; Welsh, C.J. Risk factors for gambling and substance use among recent college students. Drug Alcohol Depend. 2017, 179, 280–290. [Google Scholar] [CrossRef]
- Lang, M.; Leménager, T.; Streit, F.; Fauth-Bühler, M.; Frank, J.; Juraeva, D.; Witt, S.H.; Degenhardt, F.; Hofmann, A.; Heilmann-Heimbach, S.; et al. Genome-wide association study of pathological gambling. Eur. Psychiatry 2016, 36, 38–46. [Google Scholar] [CrossRef]
- Risner, V.A.; Benca-Bachman, C.E.; Bertin, L.; Smith, A.K.; Kaprio, J.; McGeary, J.E.; Chesler, E.; Knopik, V.S.; Friedman, N.P.; Palmer, R.H.C. Multi-Polygenic Analysis of Nicotine Dependence in Individuals of European Ancestry. Nicotine Tob. Res. 2021, 23, 2102–2109. [Google Scholar] [CrossRef]
- Muvvala, S.B.; O’Malley, S.S.; Rosenheck, R. Multiple Psychiatric Morbidity and Continued Use of Naltrexone for Alcohol Use Disorder. Am. J. Addict. 2021, 30, 55–64. [Google Scholar] [CrossRef]
- Singh, D.; Saadabadi, A. Naltrexone. In StatPearls; StatPearls Publishing: Treasure Island, FL, USA, 2022. [Google Scholar]
- Theriot, J.; Sabir, S.; Azadfard, M. Opioid Antagonists. In StatPearls; StatPearls Publishing: Treasure Island, FL, USA, 2022. [Google Scholar]
- Ward, S.; Smith, N.; Bowden-Jones, H. The use of naltrexone in pathological and problem gambling: A UK case series. J. Behav. Addict. 2018, 7, 827–833. [Google Scholar] [CrossRef]
- Grant, J.E.; Potenza, M.N.; Kraus, S.W.; Petrakis, I.L. Naltrexone and Disulfiram Treatment Response in Veterans With Alcohol Dependence and Co-Occurring Problem-Gambling Features. J. Clin. Psychiatry 2017, 78, e1299–e1306. [Google Scholar] [CrossRef]
- Weera, M.M.; Fields, M.A.; Tapp, D.N.; Grahame, N.J.; Chester, J.A. Effects of Nicotine on Alcohol Drinking in Female Mice Selectively Bred for High or Low Alcohol Preference. Alcohol Clin. Exp. Res. 2018, 42, 432–443. [Google Scholar] [CrossRef]
- Ginley, M.K.; Rash, C.J.; Olmstead, T.A.; Petry, N.M. Contingency management treatment in cocaine using methadone maintained patients with and without legal problems. Drug Alcohol Depend. 2017, 180, 208–214. [Google Scholar] [CrossRef]
- Blum, K.; Bailey, J.; Gonzalez, A.M.; Oscar-Berman, M.; Liu, Y.; Giordano, J.; Braverman, E.; Gold, M. Neuro-Genetics of Reward Deficiency Syndrome (RDS) as the Root Cause of “Addiction Transfer”: A New Phenomenon Common after Bariatric Surgery. J. Genet. Syndr. Gene Ther. 2011, 2012, S2-001. [Google Scholar] [CrossRef] [Green Version]
- Kim, H.S.; Hodgins, D.C.; Garcia, X.; Ritchie, E.V.; Musani, I.; McGrath, D.S.; von Ranson, K.M. A systematic review of addiction substitution in recovery: Clinical lore or empirically based? Clin. Psychol. Rev. 2021, 89, 102083. [Google Scholar] [CrossRef]
- Khokhar, J.Y.; Dwiel, L.L.; Henricks, A.M.; Doucette, W.T.; Green, A.I. The link between schizophrenia and substance use disorder: A unifying hypothesis. Schizophr. Res. 2018, 194, 78–85. [Google Scholar] [CrossRef]
- Shaw, M.; Black, D.W. Internet addiction: Definition, assessment, epidemiology and clinical management. CNS Drugs 2008, 22, 353–365. [Google Scholar] [CrossRef]
- Brakoulias, V.; Starcevic, V.; Albert, U.; Arumugham, S.S.; Bailey, B.E.; Belloch, A.; Borda, T.; Dell’Osso, L.; Elias, J.A.; Falkenstein, M.J.; et al. The rates of co-occurring behavioural addictions in treatment-seeking individuals with obsessive-compulsive disorder: A preliminary report. Int. J. Psychiatry Clin. Pract. 2020, 24, 173–175. [Google Scholar] [CrossRef]
- Di Nicola, M.; Tedeschi, D.; De Risio, L.; Pettorruso, M.; Martinotti, G.; Ruggeri, F.; Swierkosz-Lenart, K.; Guglielmo, R.; Callea, A.; Ruggeri, G.; et al. Co-occurrence of alcohol use disorder and behavioral addictions: Relevance of impulsivity and craving. Drug Alcohol Depend. 2015, 148, 118–125. [Google Scholar] [CrossRef]
- Nguyen, C.; Mondoloni, S.; Le Borgne, T.; Centeno, I.; Come, M.; Jehl, J.; Solié, C.; Reynolds, L.M.; Durand-de Cuttoli, R.; Tolu, S.; et al. Nicotine inhibits the VTA-to-amygdala dopamine pathway to promote anxiety. Neuron 2021, 109, 2604–2615.e9. [Google Scholar] [CrossRef] [PubMed]
- Hogg, R.C.; Raggenbass, M.; Bertrand, D. Nicotinic acetylcholine receptors: From structure to brain function. Rev. Physiol. Biochem. Pharmacol. 2003, 147, 73–120. [Google Scholar] [CrossRef]
- Blum, K.; Modestino, E.J.; Gondre-Lewis, M.; Chapman, E.J.; Neary, J.; Siwicki, D.; Baron, D.; Hauser, M.; Smith, D.E.; Roy, A.K.; et al. The Benefits of Genetic Addiction Risk Score (GARS™) Testing in Substance Use Disorder (SUD). Int. J. Genom. Data Min. 2018, 2018, 115. [Google Scholar] [CrossRef] [PubMed]
- Cosker, E.; Schwitzer, T.; Ramoz, N.; Ligier, F.; Lalanne, L.; Gorwood, P.; Schwan, R.; Laprévote, V. The effect of interactions between genetics and cannabis use on neurocognition. A review. Prog. Neuro-Psychopharmacol. Biol. Psychiatry 2018, 82, 95–106. [Google Scholar] [CrossRef]
- Hartman, C.A.; Hopfer, C.J.; Haberstick, B.; Rhee, S.H.; Crowley, T.J.; Corley, R.P.; Hewitt, J.K.; Ehringer, M.A. The association between cannabinoid receptor 1 gene (CNR1) and cannabis dependence symptoms in adolescents and young adults. Drug Alcohol Depend. 2009, 104, 11–16. [Google Scholar] [CrossRef] [Green Version]
- Comings, D.E.; Muhleman, D.; Gade, R.; Johnson, P.; Verde, R.; Saucier, G.; MacMurray, J. Cannabinoid receptor gene (CNR1): Association with i.v. drug use. Mol. Psychiatry 1997, 2, 161–168. [Google Scholar] [CrossRef]
- Noble, E.P.; Blum, K.; Ritchie, T.; Montgomery, A.; Sheridan, P.J. Allelic association of the D2 dopamine receptor gene with receptor-binding characteristics in alcoholism. Arch. Gen. Psychiatry 1991, 48, 648–654. [Google Scholar] [CrossRef]
- Hirvonen, M.M.; Laakso, A.; Någren, K.; Rinne, J.O.; Pohjalainen, T.; Hietala, J. C957T polymorphism of dopamine D2 receptor gene affects striatal DRD2 in vivo availability by changing the receptor affinity. Synapse 2009, 63, 907–912. [Google Scholar] [CrossRef]
- Voisey, J.; Swagell, C.D.; Hughes, I.P.; van Daal, A.; Noble, E.P.; Lawford, B.R.; Young, R.M.; Morris, C.P. A DRD2 and ANKK1 haplotype is associated with nicotine dependence. Psychiatry Res. 2012, 196, 285–289. [Google Scholar] [CrossRef] [Green Version]
- Connor, J.P.; Young, R.M.; Lawford, B.R.; Saunders, J.B.; Ritchie, T.L.; Noble, E.P. Heavy nicotine and alcohol use in alcohol dependence is associated with D2 dopamine receptor (DRD2) polymorphism. Addict. Behav. 2007, 32, 310–319. [Google Scholar] [CrossRef]
- Gilbert, D.; McClernon, J.; Rabinovich, N.; Sugai, C.; Plath, L.; Asgaard, G.; Zuo, Y.; Huggenvik, J.; Botros, N. Effects of quitting smoking on EEG activation and attention last for more than 31 days and are more severe with stress, dependence, DRD2 A1 allele, and depressive traits. Nicotine Tob. Res. 2004, 6, 249–267. [Google Scholar] [CrossRef]
- Ponce, G.; Jimenez-Arriero, M.A.; Rubio, G.; Hoenicka, J.; Ampuero, I.; Ramos, J.A.; Palomo, T. The A1 allele of the DRD2 gene (TaqI A polymorphisms) is associated with antisocial personality in a sample of alcohol-dependent patients. Eur. Psychiatry 2003, 18, 356–360. [Google Scholar] [CrossRef]
- Balldin, J.; Berglund, K.J.; Berggren, U.; Wennberg, P.; Fahlke, C. TAQ1A1 Allele of the DRD2 Gene Region Contribute to Shorter Survival Time in Alcohol Dependent Individuals When Controlling for the Influence of Age and Gender. A Follow-up Study of 18 Years. Alcohol Alcohol. 2018, 53, 216–220. [Google Scholar] [CrossRef] [Green Version]
- Lawford, B.R.; Young, R.M.; Noble, E.P.; Sargent, J.; Rowell, J.; Shadforth, S.; Zhang, X.; Ritchie, T. The D(2) dopamine receptor A(1) allele and opioid dependence: Association with heroin use and response to methadone treatment. Am. J. Med. Genet. 2000, 96, 592–598. [Google Scholar] [CrossRef]
- Perez de los Cobos, J.; Baiget, M.; Trujols, J.; Sinol, N.; Volpini, V.; Banuls, E.; Calafell, F.; Luquero, E.; del Rio, E.; Alvarez, E. Allelic and genotypic associations of DRD2 TaqI A polymorphism with heroin dependence in Spanish subjects: A case control study. Behav. Brain Funct. 2007, 3, 25. [Google Scholar] [CrossRef] [Green Version]
- Noble, E.P.; Blum, K.; Khalsa, M.E.; Ritchie, T.; Montgomery, A.; Wood, R.C.; Fitch, R.J.; Ozkaragoz, T.; Sheridan, P.J.; Anglin, M.D.; et al. Allelic association of the D2 dopamine receptor gene with cocaine dependence. Drug Alcohol Depend. 1993, 33, 271–285. [Google Scholar] [CrossRef]
- Blum, K.; Baron, D.; Badgaiyan, R.D.; Gold, M.S. Can Chronic Consumption of Caffeine by Increasing D2/D3 Receptors Offer Benefit to Carriers of the DRD2 A1 Allele in Cocaine Abuse? EC Psychol. Psychiatry 2019, 8, 318–321. [Google Scholar]
- D’Souza, U.M.; Russ, C.; Tahir, E.; Mill, J.; McGuffin, P.; Asherson, P.J.; Craig, I.W. Functional effects of a tandem duplication polymorphism in the 5′flanking region of the DRD4 gene. Biol. Psychiatry 2004, 56, 691–697. [Google Scholar] [CrossRef]
- Gervasini, G.; Gordillo, I.; García-Herráiz, A.; Flores, I.; Jiménez, M.; Monge, M.; Carrillo, J.A. Influence of dopamine polymorphisms on the risk for anorexia nervosa and associated psychopathological features. J. Clin. Psychopharmacol. 2013, 33, 551–555. [Google Scholar] [CrossRef]
- Thomson, C.J.; Hanna, C.W.; Carlson, S.R.; Rupert, J.L. The -521 C/T variant in the dopamine-4-receptor gene (DRD4) is associated with skiing and snowboarding behavior. Scand. J. Med. Sci. Sport. 2013, 23, e108–e113. [Google Scholar] [CrossRef] [PubMed]
- Villalba, K.; Devieux, J.G.; Rosenberg, R.; Cadet, J.L. DRD2 and DRD4 genes related to cognitive deficits in HIV-infected adults who abuse alcohol. Behav. Brain Funct. 2015, 11, 25. [Google Scholar] [CrossRef] [PubMed]
- Blum, K.; Kazmi, S.; Modestino, E.J.; Downs, B.W.; Bagchi, D.; Baron, D.; McLaughlin, T.; Green, R.; Jalali, R.; Thanos, P.K.; et al. A Novel Precision Approach to Overcome the “Addiction Pandemic” by Incorporating Genetic Addiction Risk Severity (GARS) and Dopamine Homeostasis Restoration. J. Pers. Med. 2021, 11, 212. [Google Scholar] [CrossRef] [PubMed]
- Lachowicz, M.; Chmielowiec, J.; Chmielowiec, K.; Suchanecka, A.; Masiak, J.; Michałowska-Sawczyn, M.; Mroczek, B.; Mierzecki, A.; Ciechanowicz, I.; Grzywacz, A. Significant association of DRD2 and ANKK1 genes with rural heroin dependence and relapse in men. Ann. Agric. Environ. Med. 2020, 27, 269–273. [Google Scholar] [CrossRef] [PubMed]
- Freels, T.G.; Gabriel, D.B.K.; Lester, D.B.; Simon, N.W. Risky decision-making predicts dopamine release dynamics in nucleus accumbens shell. Neuropsychopharmacology 2020, 45, 266–275. [Google Scholar] [CrossRef] [PubMed]
- Blum, K. Statistical Validation of Risk Alleles in Genetic Addiction Risk Severity (GARS) Test: Early Identification of Risk for Alcohol Use Disorder (AUD) in 74,566 case-control subjects. In Proceedings of the 19 th Annual World Congress for Brain Mapping and Therapeutics of SMBT, Los Angeles, CA, USA, 10–13 March 2022. [Google Scholar]
- Blum, K.; Oscar-Berman, M.; Demetrovics, Z.; Barh, D.; Gold, M.S. Genetic Addiction Risk Score (GARS): Molecular neurogenetic evidence for predisposition to Reward Deficiency Syndrome (RDS). Mol. Neurobiol. 2014, 50, 765–796. [Google Scholar] [CrossRef] [Green Version]
- Treister, R.; Pud, D.; Ebstein, R.P.; Laiba, E.; Raz, Y.; Gershon, E.; Haddad, M.; Eisenberg, E. Association between polymorphisms in serotonin and dopamine-related genes and endogenous pain modulation. J. Pain 2011, 12, 875–883. [Google Scholar] [CrossRef]
- Alijanpour, S.; Zarrindast, M.R. Potentiation of morphine-induced antinociception by harmaline: Involvement of μ-opioid and ventral tegmental area NMDA receptors. Psychopharmacology 2020, 237, 557–570. [Google Scholar] [CrossRef]
- Blum, K.; Hauser, M.; Fratantonio, J.; Badgaiyan, R.D. Molecular Genetic Testing in Pain and Addiction: Facts, Fiction and Clinical Utility. Addict. Genet. 2015, 2, 1–5. [Google Scholar] [CrossRef]
- Chen, A.L.; Chen, T.J.; Waite, R.L.; Reinking, J.; Tung, H.L.; Rhoades, P.; Downs, B.W.; Braverman, E.; Braverman, D.; Kerner, M.; et al. Hypothesizing that brain reward circuitry genes are genetic antecedents of pain sensitivity and critical diagnostic and pharmacogenomic treatment targets for chronic pain conditions. Med. Hypotheses 2009, 72, 14–22. [Google Scholar] [CrossRef] [Green Version]
- Moran, M.; Blum, K.; Ponce, J.V.; Lott, L.; Gondré-Lewis, M.C.; Badgaiyan, S.; Brewer, R.; Downs, B.W.; Fynman, P.; Weingarten, A.; et al. High Genetic Addiction Risk Score (GARS) in Chronically Prescribed Severe Chronic Opioid Probands Attending Multi-pain Clinics: An Open Clinical Pilot Trial. Mol. Neurobiol. 2021, 58, 3335–3346. [Google Scholar] [CrossRef]
- Downs, B.W.; Blum, K.; Bagchi, D.; Kushner, S.; Bagchi, M.; Galvin, J.M.; Lewis, M.; Siwicki, D.; Brewer, R.; Boyett, B.; et al. Molecular neuro-biological and systemic health benefits of achieving dopamine homeostasis in the face of a catastrophic pandemic (COVID-19): A mechanistic exploration. J. Syst. Integr. Neurosci. 2020, 7. [Google Scholar] [CrossRef]
- Bills, K.B.; Obray, J.D.; Clarke, T.; Parsons, M.; Brundage, J.; Yang, C.H.; Kim, H.Y.; Yorgason, J.T.; Blotter, J.D.; Steffensen, S.C. Mechanical stimulation of cervical vertebrae modulates the discharge activity of ventral tegmental area neurons and dopamine release in the nucleus accumbens. Brain Stimul. 2020, 13, 403–411. [Google Scholar] [CrossRef] [Green Version]
- di Volo, M.; Morozova, E.O.; Lapish, C.C.; Kuznetsov, A.; Gutkin, B. Dynamical ventral tegmental area circuit mechanisms of alcohol-dependent dopamine release. Eur. J. Neurosci. 2019, 50, 2282–2296. [Google Scholar] [CrossRef] [Green Version]
- Tye, K.M.; Mirzabekov, J.J.; Warden, M.; Ferenczi, E.A.; Tsai, H.-C.; Finkelstein, J.; Kim, S.-Y.; Adhikari, A.; Thompson, K.R.; Andalman, A.S.; et al. Dopamine neurons modulate neural encoding and expression of depression-related behaviour. Nature 2012, 493, 537–541. [Google Scholar] [CrossRef] [Green Version]
- Liu, C.; Kaeser, P.S. Mechanisms and regulation of dopamine release. Curr. Opin. Neurobiol. 2019, 57, 46–53. [Google Scholar] [CrossRef]
- Green, A.I.; Zimmet, S.V.; Strous, R.D.; Schildkraut, J.J. Clozapine for comorbid substance use disorder and schizophrenia: Do patients with schizophrenia have reward-deficiency syndrome that can be ameliorated by clozapine? Harv. Rev. Psychiatry 1999, 6, 287–296. [Google Scholar] [CrossRef]
- Cervinski, M.A.; Foster, J.D.; Vaughan, R.A. Psychoactive substrates stimulate dopamine transporter phosphorylation and down-regulation by cocaine-sensitive and protein kinase C-dependent mechanisms. J. Biol. Chem. 2005, 280, 40442–40449. [Google Scholar] [CrossRef] [Green Version]
- Archer, T.; Oscar-Berman, M.; Blum, K.; Gold, M. Epigenetic Modulation of Mood Disorders. J. Genet. Syndr. Gene Ther. 2013, 4, 1000120. [Google Scholar] [CrossRef] [Green Version]
- Blum, K. Genetic Addiction Risk Severity Test and its Clinical Applications. In Proceedings of the Society Brain Mapping Therapeutics 19 th Annual Meeting, Los Angeles, CA, USA, 10–13 March 2022. [Google Scholar]
- Blum, K.; Chen, T.J.; Meshkin, B.; Downs, B.W.; Gordon, C.A.; Blum, S.; Mangucci, J.F.; Braverman, E.R.; Arcuri, V.; Deutsch, R.; et al. Genotrim, a DNA-customized nutrigenomic product, targets genetic factors of obesity: Hypothesizing a dopamine-glucose correlation demonstrating reward deficiency syndrome (RDS). Med. Hypotheses 2007, 68, 844–852. [Google Scholar] [CrossRef]
- Blum, K.; Chen, T.J.; Meshkin, B.; Downs, B.W.; Gordon, C.A.; Blum, S.; Mengucci, J.F.; Braverman, E.R.; Arcuri, V.; Varshavskiy, M.; et al. Reward deficiency syndrome in obesity: A preliminary cross-sectional trial with a Genotrim variant. Adv. Ther. 2006, 23, 1040–1051. [Google Scholar] [CrossRef] [PubMed]
- Blum, K.; Chen, T.J.; Williams, L.; Chen, A.L.; Downs, W.B.; Waite, R.L.; Huntington, T.; Braverman, E.R. A short term pilot open label study to evaluate efficacy and safety of LG839, a customized DNA directed nutraceutical in obesity: Exploring Nutrigenomics. Gene Ther. Mol. Biol. 2008, 12, 371–382. [Google Scholar]
- Blum, K.; Downs, B.W.; Dushaj, K.; Li, M.; Braverman, E.R.; Fried, L.; Waite, R.; Demotrovics, Z.; Badgaiyan, R.D. The Benefits of Customized DNA Directed Nutrition to Balance the Brain Reward Circuitry and Reduce Addictive Behaviors. Precis. Med. 2016, 1, 18–33. [Google Scholar]
- Blum, K.; Oscar-Berman, M.; Giordano, J.; Downs, B.W.; Simpatico, T.; Han, D.; Femino, J. Neurogenetic Impairments of Brain Reward Circuitry Links to Reward Deficiency Syndrome (RDS): Potential Nutrigenomic Induced Dopaminergic Activation. J. Genet. Syndr. Gene Ther. 2012, 3, 1000e115. [Google Scholar] [CrossRef] [PubMed]
- Blum, K.; Chen, T.J.; Chen, A.L.; Rhoades, P.; Prihoda, T.J.; Downs, B.W.; Bagchi, D.; Bagchi, M.; Blum, S.H.; Williams, L.; et al. Dopamine D2 Receptor Taq A1 allele predicts treatment compliance of LG839 in a subset analysis of pilot study in the Netherlands. Gene Ther. Mol. Biol. 2008, 12, 129–140. [Google Scholar]
- Substance Abuse and Mental Health Services Administration (US); Office of the Surgeon General (US). Facing Addiction in America: The Surgeon General’s Report on Alcohol, Drugs, and Health; US Department of Health and Human Services: Washington, DC, USA, 2016.
- McLellan, A.T.; Koob, G.F.; Volkow, N.D. Preaddiction-A Missing Concept for Treating Substance Use Disorders. JAMA Psychiatry 2022, ahead of print. [Google Scholar] [CrossRef]
- Diabetes Prevention Program Research Group; Knowler, W.C.; Fowler, S.E.; Hamman, R.F.; Christophi, C.A.; Hoffman, H.J.; Brenneman, A.T.; Brown-Friday, J.O.; Goldberg, R.; Venditti, E.; et al. 10-Year follow-up of diabetes incidence and weight loss in the Diabetes Prevention Program Outcomes Study. Lancet 2009, 374, 1677–1686. [Google Scholar] [CrossRef] [Green Version]
- Glechner, A.; Keuchel, L.; Affengruber, L.; Titscher, V.; Sommer, I.; Matyas, N.; Wagner, G.; Kien, C.; Klerings, I.; Gartlehner, G. Effects of lifestyle changes on adults with prediabetes: A systematic review and meta-analysis. Prim. Care Diabetes 2018, 12, 393–408. [Google Scholar] [CrossRef]
- Blum, K.; Soni, D.; Badgaiyan, R.D.; Baron, D. Overcoming reward deficiency syndrome by the induction of “dopamine homeostasis” instead of opioids for addiction: Illusion or reality? J. Osteopath. Med. 2022, 122, 333–337. [Google Scholar] [CrossRef]
- Blum, K.; Bowirrat, A.; Gondre Lewis, M.C.; Simpatico, T.A.; Ceccanti, M.; Steinberg, B.; Modestino, E.J.; Thanos, P.K.; Baron, D.; McLaughlin, T.; et al. Exploration of Epigenetic State Hyperdopaminergia (Surfeit) and Genetic Trait Hypodopaminergia(Deficit) During Adolescent Brain Development. Curr. Psychopharmacol. 2021, 10. [Google Scholar] [CrossRef]
- Edwards, D.; Roy, A.K., III; Boyett, B.; Badgaiyan, R.D.; Thanos, P.K.; Baron, D.; Hauser, M.; Badgaiyan, S.; Brewer, R.; Siwicki, D.B.; et al. Addiction by Any Other Name is Still Addiction: Embracing Molecular Neurogenetic/Epigenetic Basis of Reward Deficiency. J. Addict. Sci. 2020, 6, 1–4. [Google Scholar] [CrossRef]
- Kótyuk, E.; Urbán, R.; Hende, B.; Richman, M.; Magi, A.; Király, O.; Barta, C.; Griffiths, M.D.; Potenza, M.N.; Badgaiyan, R.D.; et al. Development and validation of the Reward Deficiency Syndrome Questionnaire (RDSQ-29). J. Psychopharmacol. 2022, 36, 409–422. [Google Scholar] [CrossRef]
- Novi, M.; Paraskevopoulou, M.; Van Rooij, D.; Schene, A.H.; Buitelaar, J.K.; Schellekens, A.F.A. Effects of substance misuse and current family history of substance use disorder on brain structure in adolescents and young adults with attention-deficit/hyperactivity disorder. Drug Alcohol Depend. 2021, 228, 109032. [Google Scholar] [CrossRef]
- Blum, K.; Bowirrat, A.; Baron, D.; Lott, L.; Ponce, J.V.; Brewer, R.; Siwicki, D.; Boyett, B.; Gondre-Lewis, M.C.; Smith, D.E.; et al. Biotechnical development of genetic addiction risk score (GARS) and selective evidence for inclusion of polymorphic allelic risk in substance use disorder (SUD). J. Syst. Integr. Neurosci. 2020, 6. [Google Scholar] [CrossRef]
- Fried, L.; Modestino, E.J.; Siwicki, D.; Lott, L.; Thanos, P.K.; Baron, D.; Badgaiyan, R.D.; Ponce, J.V.; Giordano, J.; Downs, W.B.; et al. Hypodopaminergia and Precision Behavioral Management (PBM): It is a Generational Family Affair. Curr. Pharm. Biotechnol. 2020, 21, 528–541. [Google Scholar] [CrossRef]
- Blum, K. Criterion Validity of the Genetic Addiction Risk Severity (GARS) as a Marker of Reward Deficiency in Chemical Substances’ Addiction: A Multi-Center Study, Kenneth Blum. In Proceedings of the 7 th Neurological Disorders Summit, San Francisco, CA, USA, 18–21 July 2022. [Google Scholar]
- Hyman, S.E. The daunting polygenicity of mental illness: Making a new map. Philos. Trans. R. Soc. B Biol. Sci. 2018, 373, 20170031. [Google Scholar] [CrossRef]
- Nahum, L.H. Addiction and preaddiction. Conn. Med. 1971, 35, 560. [Google Scholar]
- Available online: https://www.google.com/search?sxsrf=ALiCzsY4_yGbAAPZN8xupN3x1sBXyRyuoA:1659404973972&q=Operationalizing+%E2%80%98prediction%E2%80%99+using+the+DSM-+5+criteria+for+substance+use+disorder:+Challenges+and+limitations&spell=1&sa=X&ved=2ahUKEwj2-oughaf5AhW-omoFHT3JAmEQBSgAegQIARAz&biw=1904&bih=927&dpr=1 (accessed on 1 June 2022).



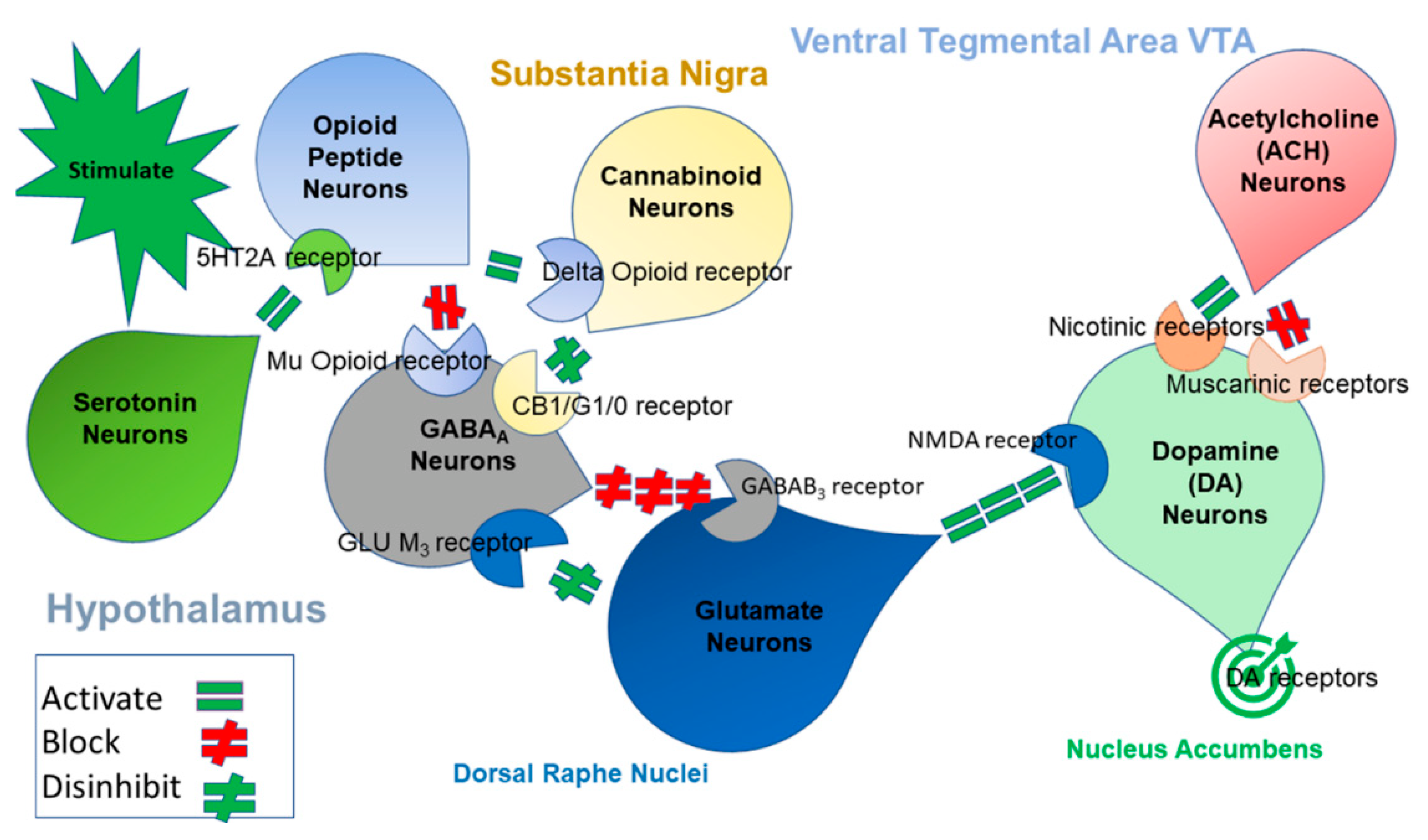{kind=link}
{kind=link}
| Gene/Polymorphism | Number of Studies | Overall Summary |
|---|---|---|
| Dopamine D1 Receptor (DRD1): rs4532—risk allele G | 3 | Several studies supported that genetic variation in DRD1 may influence genetic predisposition to alcoholism. A statistically significant association of DRD1 rs4532 polymorphism with alcohol dependence was found among Indian males (90 cases vs. 122 controls). Other studies also demonstrated that this could be associated with the impulsivity and aggression of AUD patients. |
| Dopamine D2 Receptor (DRD2): rs1800497—risk allele A1 | 118 | The DRD2 rs1800497 was found to be associated with the risk of AUD and several AUD-related conditions in a meta-analysis of numerous case–control studies (total of 18,290 cases vs. 19,809 controls, including US Caucasian, native and African American, British, French, Italian, Swedish, Finnish, Spanish, Mexican, Brazilian, Scandinavian, and Japanese) pooled with the random effect models. |
| Dopamine D3 Receptor (DRD3): rs6280—risk allele C (Ser9Gly) | 3 | Several case–control studies investigated the association between the DRD3 rs6280 polymorphism and alcohol dependence. In a Korean study (243 cases vs. 130 controls), the DRD3 rs6280 polymorphism was significantly associated with AUD development. |
| Dopamine D4 Receptor (DRD4): rs1800955—risk allele C (48bp repeat VNTR) | 35 | The DRD4 rs1800955 polymorphism was found to be associated with the risk of developing AUD and AUD-related conditions in a meta-analysis of various case–control studies (total of 2997 cases vs. 2588 controls, including US Caucasian, Mexican American, and Indian) pooled with the random effect models. |
| Dopamine Transporter Receptor (DAT1): SLC6A3 3’-UTR—risk allele A9 (40bp repeat VNTR) | 43 | The central dopaminergic reward pathway is likely involved in alcohol intake and the progression of alcohol dependence. DAT1 is a principal regulator of dopaminergic neurotransmission. From the meta-analysis of numerous case–control studies (total of 3790 cases vs. 3446 controls) pooled with the random effect models, the DAT1 SLC6A3 3’-UTR risk allele was found to be marginally associated with the risk of AUD and AUD-related conditions. |
| Catechol-O-Methyltransferase (COMT): rs4680—risk allele G (Val158Met) | 13 | COMT is a strong candidate gene that contributes to SUD and schizophrenia. A meta-analysis of several case–control studies (total of 1212 cases vs. 933 controls, including US Caucasian, Finnish, Croatian, and Taiwanese) pooled with a random effect model, the association of COMPT rs4680 polymorphism with the risk of developing AUD and AUD-related conditions was found with marginal statistical significance. |
| µ-Opioid Receptor (OPRM1): rs1799971—risk allele G (A118G) | 28 | Opioid receptors play an essential role in ethanol reinforcement and alcohol dependence risk. Polymorphisms in the OPRM1 gene expressing µ-opioid receptors could be significantly associated with some features of alcohol dependence. From the meta-analysis of case–control studies (total of 3096 cases vs. 2896 controls, including US Caucasian, Spanish, Turkish, and Asian), pooled with the random effect model, the results indicated that a functional OPRM variant is associated with the risk of alcohol dependence with marginal statistical significance. |
| γ-Aminobutyric Acid (GABA) A Receptor, β-3 Subunit (GABRB3): CA repeat—risk allele 181 | 6 | The GABAergic system has been implicated in alcohol-related behaviors. From case–control studies (171 cases vs. 45 controls), the association of variants of the GABRB3 gene with alcohol dependence is, however, inconclusive. A more extensive controlled study is required for improved results. |
| Monoamine Oxidase A (MAO-A): 3’ 30bp VNTR -risk allele 4R DNRP | 6 | The function of monoamine oxidase (MAO) in alcoholism was determined using several case–control studies (170 cases vs. 177 controls). Although genetic heterogeneity is suspected of underlying alcoholism and MAO-A mutations may play a role in susceptibility to alcoholism, the overall results were not found to be statistically significant. A more extensive controlled study is required to obtain conclusive results. |
| Serotonin Transporter Receptor (5HTT) Linked Promoter Region (5HTTLPR) in SLC6A4: rs25531—risk allele S’ | 20 | Serotonin (5-HT) has been demonstrated to regulate alcohol consumption. Since the activity of the 5-HT transporter protein (5-HTT) regulates 5-HT levels, it may contribute to the risk of alcohol dependence. A meta-analysis of case–control studies (total 9996 cases vs. 9950 controls) pooled with the random effect models showed a significant association between alcohol dependence and the serotonin-transporter-linked promoter region (5-HTTLPR), which is a polymorphic region in the SLC6A4 gene. |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Blum, K.; Han, D.; Bowirrat, A.; Downs, B.W.; Bagchi, D.; Thanos, P.K.; Baron, D.; Braverman, E.R.; Dennen, C.A.; Gupta, A.; et al. Genetic Addiction Risk and Psychological Profiling Analyses for “Preaddiction” Severity Index. J. Pers. Med. 2022, 12, 1772. https://doi.org/10.3390/jpm12111772
Blum K, Han D, Bowirrat A, Downs BW, Bagchi D, Thanos PK, Baron D, Braverman ER, Dennen CA, Gupta A, et al. Genetic Addiction Risk and Psychological Profiling Analyses for “Preaddiction” Severity Index. Journal of Personalized Medicine. 2022; 12(11):1772. https://doi.org/10.3390/jpm12111772
Chicago/Turabian StyleBlum, Kenneth, David Han, Abdalla Bowirrat, Bernard William Downs, Debasis Bagchi, Panayotis K. Thanos, David Baron, Eric R. Braverman, Catherine A. Dennen, Ashim Gupta, and et al. 2022. "Genetic Addiction Risk and Psychological Profiling Analyses for “Preaddiction” Severity Index" Journal of Personalized Medicine 12, no. 11: 1772. https://doi.org/10.3390/jpm12111772