
The Prevalence of Multidrug-Resistant Enterobacteriaceae among Neonates in Kuwait

Abstract
:1. Introduction
2. Materials and Methods
2.1. Sample Collection
2.2. Isolation and Identification
2.3. Susceptibility Test by E-Test Method
2.4. Molecular Detection of Resistance Genes
2.5. Sequencing of Resistance Genes
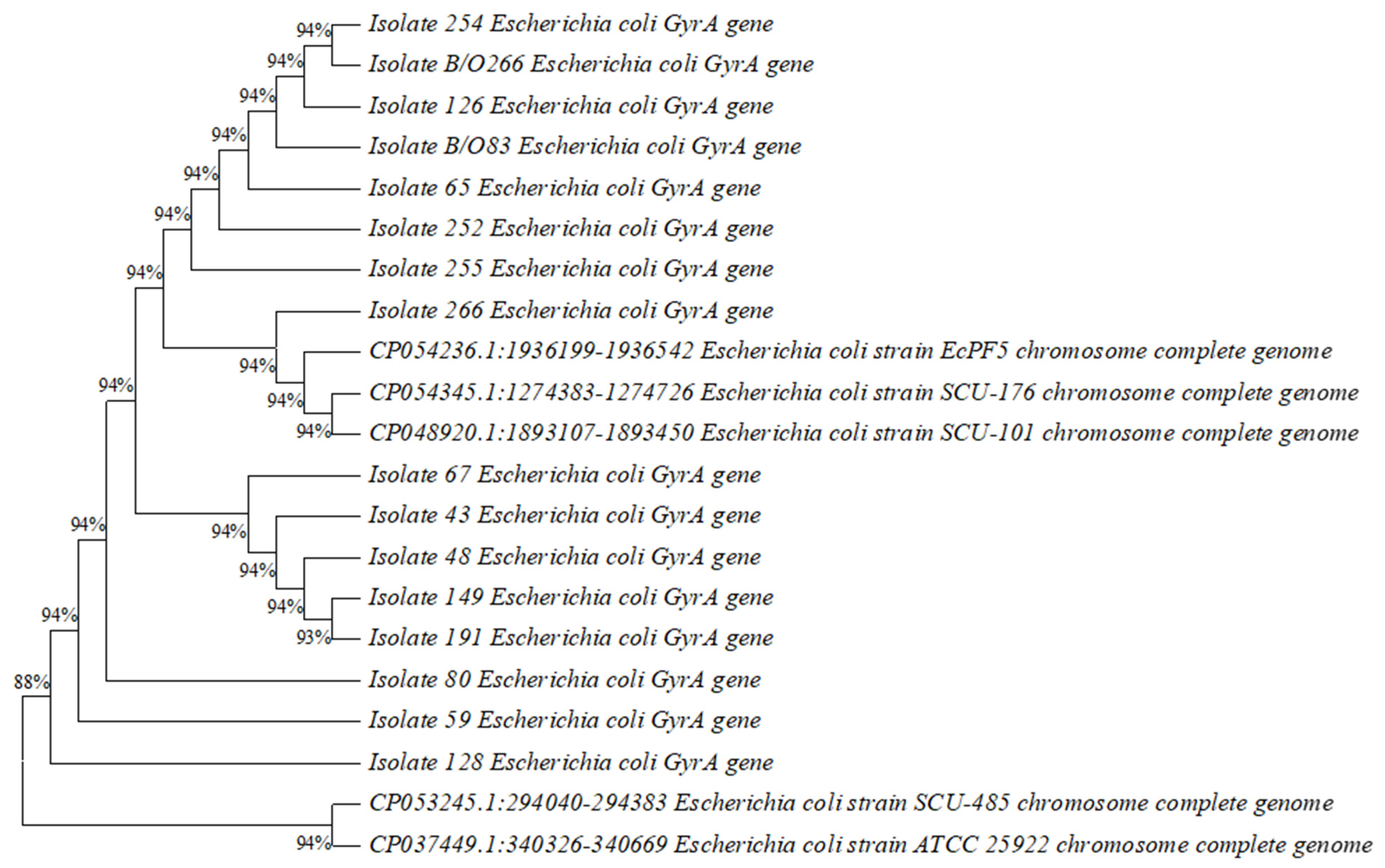
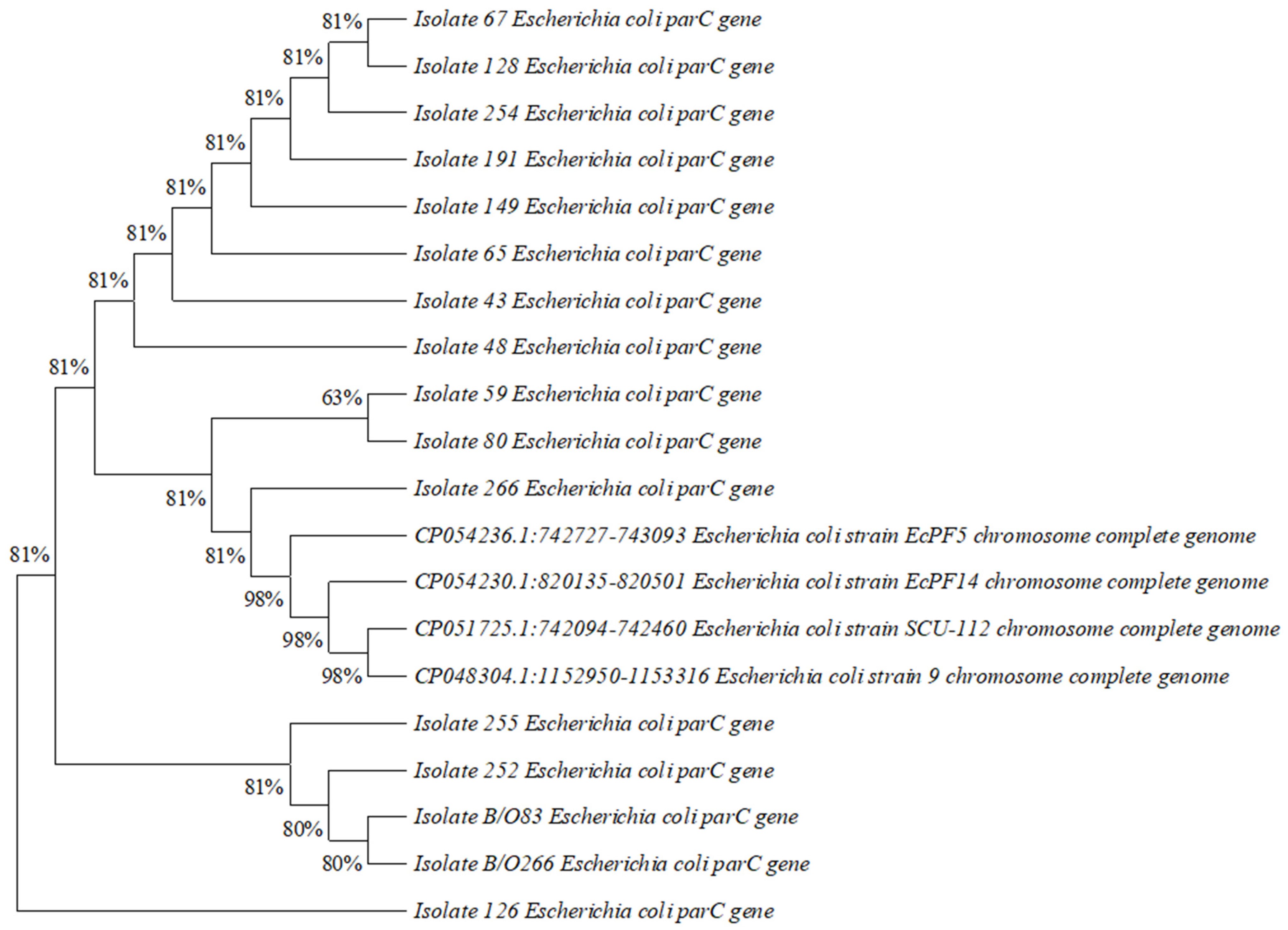
2.6. Construction of Phylogenetic Tree
3. Results
3.1. Isolation and Identification
3.2. Susceptibility Test by E-Test Method
3.3. Molecular Detection of Resistance Genes
3.4. Sequencing of Resistance Genes
3.5. Construction of Phylogenetic Tree
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Acknowledgments
Conflicts of Interest
References
- Riedel, S.; Hobden, J.A.; Miller, S.; Morse, S.A.; Mietzner, T.A.; Detrick, B.; Mitchell, T.G.; Sakanari, J.A.; Hotez, P.; Mejia, R. Enteric gram-negative rods (Enterobacteriaceae). In Jawetz, Melnick & Adelberg’s Medical Microbiology; McGraw Hill: New York, NY, USA, 2019. [Google Scholar]
- Peleg, A.Y.; Hooper, D.C. Hospital-Acquired Infections Due to Gram-Negative Bacteria. N. Engl. J. Med. 2010, 362, 1804–1813. [Google Scholar] [CrossRef]
- Aslam, B.; Wang, W.; Arshad, M.I.; Khurshid, M.; Muzammil, S.; Nisar, M.A.; Alvi, R.F.; Aslam, M.A.; Qamar, M.U.; Salamat, M.K.F.; et al. Antibiotic resistance: A rundown of a global crisis. Infect. Drug Resist. 2018, 11, 1645–1658. [Google Scholar] [CrossRef] [PubMed]
- WHO. Global Antimicrobial Resistance and Use Surveillance System (GLASS) Report 2021; World Health Organization: Geneva, Switzerland, 2021. [Google Scholar]
- WHO. Antibacterial Agents in Clinical and Preclinical Development: An Overview and Analysis 2021; World Health Organization: Geneva, Switzerland, 2022. [Google Scholar]
- Partridge, S.R. Resistance mechanisms in Enterobacteriaceae. Pathology 2015, 47, 276–284. [Google Scholar] [CrossRef] [PubMed]
- Folgori, L.; Bielicki, J. Future Challenges in Pediatric and Neonatal Sepsis: Emerging Pathogens and Antimicrobial Resistance. J. Pediatr. Intensiv. Care 2019, 8, 17–24. [Google Scholar] [CrossRef] [PubMed]
- Andersson, D. The Ways in Which Bacteria Resist Antibiotics; World Health Organization: Geneva, Switzerland, 2005. [Google Scholar]
- Antunes, N.T.; Fisher, J.F. Acquired Class D β-Lactamases. Antibiotics 2014, 3, 398–434. [Google Scholar] [CrossRef] [PubMed]
- De Angelis, G.; Del Giacomo, P.; Posteraro, B.; Sanguinetti, M.; Tumbarello, M. Molecular Mechanisms, Epidemiology, and Clinical Importance of β-Lactam Resistance in Enterobacteriaceae. Int. J. Mol. Sci. 2020, 21, 5090. [Google Scholar] [CrossRef] [PubMed]
- Giske, C.; Martínez-Martínez, L.; Glupczynski, Y.; Skov, G.; Cantón, R. EUCAST Guidelines for Detection of Resistance Mechanisms and Specific Resistances of Clinical and/or Epidemiological Importance, Version 2.0. 2017. Available online: https://www.semanticscholar.org/paper/EUCAST-guidelines-for-detection-of-resistance-and-Giske-Mart%C3%ADnez-Mart%C3%ADnez/8fa90d4e48d77c9fb0e33a4e700d45cdde09b232 (accessed on 11 March 2023).
- Toussaint, K.A.; Gallagher, J.C. β-Lactam/β-Lactamase Inhibitor Combinations: From Then to Now. Ann. Pharmacother. 2014, 1, 86–98. [Google Scholar] [CrossRef] [PubMed]
- Di Conzaa, J.A.; Badaraccoa, A.; Ayala, J.; Rodríguez, C.; Famiglietti, A.; Gutkind, G.O. Beta-lactamases produced by amoxi-cillin-clavulanate-resistant Enterobacteria isolated in Buenos Aires, Argentina: A new blaTEM gene. Rev. Argent. Microbiol. 2014, 46, 210–217. [Google Scholar]
- Wang, Y.; Wang, J.; Wang, R.; Cai, Y. Resistance to ceftazidime–avibactam and underlying mechanisms. J. Glob. Antimicrob. Resist. 2019, 22, 18–27. [Google Scholar] [CrossRef]
- Doi, Y.; Wachino, J.I.; Arakawa, Y. Aminoglycoside Resistance: The Emergence of Acquired 16S Ribosomal RNA Methyl-transferases. Infect. Dis. Clin. N. Am. 2016, 30, 523–537. [Google Scholar] [CrossRef]
- Davies, J.; Wright, G.D. Bacterial resistance to aminoglycoside antibiotics. Trends Microbiol. 1997, 5, 234–240. [Google Scholar] [CrossRef]
- Blondeau, J.M. Fluoroquinolones: Mechanism of action, classification, and development of resistance. Surv. Ophthalmol. 2004, 49, S73–S78. [Google Scholar] [CrossRef] [PubMed]
- Leski, T.A.; Bangura, U.; Jimmy, D.H.; Ansumana, R.; Lizewski, S.E.; Stenger, D.A.; Taitt, C.R.; Vora, G.J. Multidrug-resistant tet(X)-containing hospital isolates in Sierra Leone. Int. J. Antimicrob. Agents 2013, 42, 83–86. [Google Scholar] [CrossRef] [PubMed]
- Sheng, Z.-K.; Hu, F.; Wang, W.; Guo, Q.; Chen, Z.; Xu, X.; Zhu, D.; Wang, M. Mechanisms of Tigecycline Resistance among Klebsiella pneumoniae Clinical Isolates. Antimicrob. Agents Chemother. 2014, 58, 6982–6985. [Google Scholar] [CrossRef] [PubMed]
- Grape, M.; Motakefi, A.; Pavuluri, S.; Kahlmeter, G. Standard and real-time multiplex PCR methods for detection of trime-thoprim resistance dfr genes in large collections of bacteria. Eur. J. Clin. Microbiol. Infect. Dis. 2007, 13, 1112–1118. [Google Scholar] [CrossRef]
- Adamus-Białek, W.; Baraniak, A.; Wawszczak, M.; Głuszek, S.; Gad, B.; Wróbel, K.; Bator, P.; Majchrzak, M.; Parniewski, P. The genetic background of antibiotic resistance among clinical uropathogenic Escherichia coli strains. Mol. Biol. Rep. 2018, 45, 1055–1065. [Google Scholar] [CrossRef]
- Fernández-Villa, D.; Aguilar, M.R.; Rojo, L. Folic Acid Antagonists: Antimicrobial and Immunomodulating Mechanisms and Applications. Int. J. Mol. Sci. 2019, 20, 4996. [Google Scholar] [CrossRef]
- Wayne, P.A. Performance Standards for Antimicrobial Susceptibility Testing. In Clinical and Laboratory Standards Institute, 26th ed.; Clinical and Laboratory Standards Institute Supplement: St. Louis, MO, USA, 2016. [Google Scholar]
- Magiorakos, A.-P.; Srinivasan, A.; Carey, R.B.; Carmeli, Y.; Falagas, M.E.; Giske, C.G.; Harbarth, S.; Hindler, J.F.; Kahlmeter, G.; Olsson-Liljequist, B.; et al. Multidrug-resistant, extensively drug-resistant and pandrug-resistant bacteria: An international expert proposal for interim standard definitions for acquired resistance. Clin. Microbiol. Infect. 2012, 18, 268–281. [Google Scholar] [CrossRef]
- Nwafia, I.N.; Ohanu, M.E.; Ebede, S.O.; Ozumba, U.C. Molecular detection and antibiotic resistance pattern of extend-ed-spectrum beta-lactamase producing Escherichia coli in a Tertiary Hospital in Enugu, Nigeria. Ann. Clin. Microbiol. Antimicrob. 2019, 18, 2–4. [Google Scholar] [CrossRef]
- Poirela, L.; Walshb, T.R.; Cuvilliera, V.; Nordmann, P. Multiplex PCR for detection of acquired carbapenemase genes. Diagn. Microbiol. Infect. Dis. 2011, 70, 119–123. [Google Scholar] [CrossRef]
- Hu, X.; Xu, B.; Yang, Y.; Liu, D.; Yang, M.; Wang, J.; Shen, H.; Zhou, X.; Ma, X. A high throughput multiplex PCR assay for simultaneous detection of seven aminoglycoside-resistance genes in Enterobacteriaceae. BMC Microbiol. 2013, 13, 58. [Google Scholar] [CrossRef]
- Cano, M.E.; Rodríguez-Martínez, J.M.; Aguero, J.; Pascual, A.; Calvo, J.; García-Lobo, J.M.; Velasco, C.; Francia, M.V.; Martínez-Martínez, L. Detection of Plasmid-Mediated Quinolone Resistance Genes in Clinical Isolates of Enterobacter spp. in Spain. J. Clin. Microbiol. 2009, 47, 2033–2039. [Google Scholar] [CrossRef]
- Ji, K.; Xu, Y.; Sun, J.; Huang, M.; Jia, X.; Jiang, C.; Feng, Y. Harnessing efficient multiplex PCR methods to detect the expanding Tet(X) family of tigecycline resistance genes. Virulence 2019, 11, 49–56. [Google Scholar] [CrossRef]
- WHO. Antimicrobial Resistance: Global Report on Surveillance; World Health Organization: Geneva, Switzerland, 2014. [Google Scholar]
- Folgori, L.; Bielicki, J.; Heath, P.T.; Sharland, M. Antimicrobial-resistant Gram-negative infections in neonates: Burden of disease and challenges in treatment. Curr. Opin. Infect. Dis. 2017, 30, 281–288. [Google Scholar] [CrossRef]
- Bielicki, J.A.; Lundin, R.; Sharland, M. Antibiotic Resistance Prevalence in Routine Bloodstream Isolates from Children’s Hospitals Varies Substantially from Adult Surveillance Data in Europe. Pediatr. Infect. Dis. J. 2015, 34, 734–741. [Google Scholar] [CrossRef]
- Oluyege, A.O.; Ojo-Bola, O.; Oludada, O.E. Carriage of antibiotic resistant commensal E. coli in infants below 5 months in Ado-Ekiti. Int. J. Curr. Microbiol. Appl. Sci. 2015, 4, 1096–1102. [Google Scholar]
- Dyar, O.J.; Hoa, N.Q.; Trung, N.V.; Phuc, H.D.; Larsson, M.; Chuc, N.T.K.; Lundborg, C.S. High prevalence of antibiotic resistance in commensal Escherichia coli among children in rural Vietnam. BMC Infect. Dis. 2012, 12, 92. [Google Scholar] [CrossRef] [PubMed]
- Taqi, M.; Jamal, W.; Rotimi, V. The Prevalence of Extended-Spectrum β-lactamase (ESBL) and Carbapenem-Resistant Enter-obacteriaceae (CRE) Isolates in Positive Blood Cultures of Patients in A Teaching Hospital in Kuwait Over A 2-year Period. Open Forum Infect. Dis. 2017, 4, S564. [Google Scholar] [CrossRef]
- Moghnieh, R.A.; Kanafani, Z.A.; Tabaja, H.Z.; Sharara, S.L.; Awad, L.S.; Kanj, S.S. Epidemiology of common resistant bacterial pathogens in the countries of the Arab League. Lancet Infect. Dis. 2018, 18, e379–e394. [Google Scholar] [CrossRef]
- Bevan, E.R.; Jones, A.M.; Hawkey, P.M. Global epidemiology of CTX-M β-lactamases: Temporal and geographical shifts in genotype. J. Antimicrob. Chemother. 2017, 72, 2145–2155. [Google Scholar] [CrossRef] [PubMed]
- Garneau-Tsodikova, S.; Labby, K.J. Mechanisms of resistance to aminoglycoside antibiotics: Overview and perspectives. MedChemComm 2015, 7, 11–27. [Google Scholar] [CrossRef] [PubMed]



{kind=link}
{kind=link}
| Antibiotic Name | Concentration μg/mL |
|---|---|
| Amoxicillin-clavulanic acid | 0.016–256 μg/mL |
| Ceftolozane-tazobactam | 0.016–256 μg/mL |
| Cefazolin | 0.016–256 μg/mL |
| Cefepime | 0.016–256 μg/mL |
| Ceftaroline | 0.016–256 μg/mL |
| Cefotaxime | 0.016–256 μg/mL |
| Ceftriaxone | 0.016–256 μg/mL |
| Cefoxitin | 0.016–256 μg/mL |
| Cefixime | 0.016–256 μg/mL |
| Ceftazidime | 0.016–256 μg/mL |
| Ceftazidime-avibactam | 0.016–256 μg/mL |
| Ertapenem | 0.002–32 μg/mL |
| Imipenem | 0.002–32 μg/mL |
| Meropenem | 0.002–32 μg/mL |
| Amikacin | 0.016–256 μg/mL |
| Gentamicin | 0.064–1024 μg/mL |
| Ciprofloxacin | 0.002–32 μg/mL |
| Trimethoprim-sulfamethoxazole | 0.002–32 μg/mL |
| Tigecycline | 0.016–256 μg/mL |
| Antibiotic Group | Gene | Denaturation | Annealing | Extension | Reference |
|---|---|---|---|---|---|
| β-lactam-β-lactamase inhibitor combinations (amoxicillin-clavulanic acid) | bla-TEM, bla-SHV, bla-OXA-1, bla-OXA-2, bla-OXA-10 | 95 °C for 1 min/30 cycles | 55 °C for 1 min | 72 °C for 1 min | [13] |
| ESBL’s genes | bla-CTX-M, bla-TEM, bla-SHV | 95 °C for 30 s/35 cycles | 60 °C for 30 s | 72 °C for 1 min | [25] |
| Carbapenems | bla-IMP, bla-VIM, bla-OXA-48, bla-GIM, bla-KPC, bla-NDM | 94 °C for 30 s/36 cycles | 52 °C for 40 s | 72 °C for 50 s | [26] |
| Aminoglycosides | aac(3)-II, aac(6′)-Ib, aac(6′)-II, ant(3″)-I, aph(3′)-VI, armA, rmtB | 94 °C for 30 s/30 cycles | 56 °C for 30 s | 72 °C for 1 min | [27] |
| Fluoroquinolones | qnrA, qnrB, qnrS, qepA, aac(6)-Ib-cr | 94 °C for 1 min/30 cycles | 57 °C for 1 min | 72 °C for 1 min | [28] |
| Folate pathway inhibitor (trimethoprim-sulfamethoxazole) | sul1, sul2, sul3, dfr1, dfr5, dfr7&17 | 94 °C for 1 min/30 cycles | 57 °C for 1 min | 72 °C for 1 min | [20,21] |
| Tigecycline | tetX, tetX2, tetX3, tetX4, tetX5 | 95 °C for 30 s/30 cycles | 58 °C for 30 s | 72 °C for 30 s | [29] |
| Name of the Enterobacteriaceae Isolate | Number Among Mothers | Number Among Neonates |
|---|---|---|
| Escherichia coli | 200 | 32 |
| Klebsiella pneumoniae | 39 | 9 |
| Klebsiella oxytoca | 3 | - |
| Klebsiella ozaenae | 1 | - |
| Enterobacter cloacae | 16 | 2 |
| Enterobacter aerogenes | 1 | - |
| Citrobacter freundii | 10 | - |
| Citrobacter farmeri | 4 | 1 |
| Citrobacter braakii | 2 | - |
| Citrobacter koseri | 1 | - |
| Citrobacter amalonaticus | 1 | - |
| Kluyvera ascorbata | 4 | - |
| Shigella sonnei | 1 | - |
| Cronobacter sakazakii | 1 | - |
| Total | 284 | 44 |
| Antibiotic Name | Number of Resistant Isolates Among Mothers | Number of Resistant Isolates among Neonates |
|---|---|---|
| Amoxicillin-clavulanic acid | 36 | 3 |
| Ceftolozane-tazobactam | 34 | 3 |
| Cefazolin | 36 | 3 |
| Cefepime | 29 | 3 |
| Ceftaroline | 29 | 2 |
| Cefotaxime | 34 | 3 |
| Ceftriaxone | 34 | 3 |
| Cefoxitin | 35 | 3 |
| Cefixime | 34 | 3 |
| Ceftazidime | 34 | 3 |
| Ceftazidime-avibactam | - | 3 |
| Ertapenem | 2 | - |
| Imipenem | - | - |
| Meropenem | - | - |
| Amikacin | 1 | - |
| Gentamicin | 4 | - |
| Ciprofloxacin | 14 | 2 |
| Trimethoprim-sulfamethoxazole | 30 | 3 |
| Tigecycline | 10 | 2 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Alajmi, R.Z.; Alfouzan, W.A.; Mustafa, A.S. The Prevalence of Multidrug-Resistant Enterobacteriaceae among Neonates in Kuwait. Diagnostics 2023, 13, 1505. https://doi.org/10.3390/diagnostics13081505
Alajmi RZ, Alfouzan WA, Mustafa AS. The Prevalence of Multidrug-Resistant Enterobacteriaceae among Neonates in Kuwait. Diagnostics. 2023; 13(8):1505. https://doi.org/10.3390/diagnostics13081505
Chicago/Turabian StyleAlajmi, Rehab Zafer, Wadha Ahmed Alfouzan, and Abu Salim Mustafa. 2023. "The Prevalence of Multidrug-Resistant Enterobacteriaceae among Neonates in Kuwait" Diagnostics 13, no. 8: 1505. https://doi.org/10.3390/diagnostics13081505