From the 183 DEPs identified in our study, according to the information we reviewed (UniProtKB, Protein Data Bank, protein atlas, and NCBI), there is a large number of proteins, including their coding genes (more than 100), that we currently cannot correlate to alterations reported in the brain or to other changes previously described in the GASH/Sal.
PPI Network Analysis of DEPs
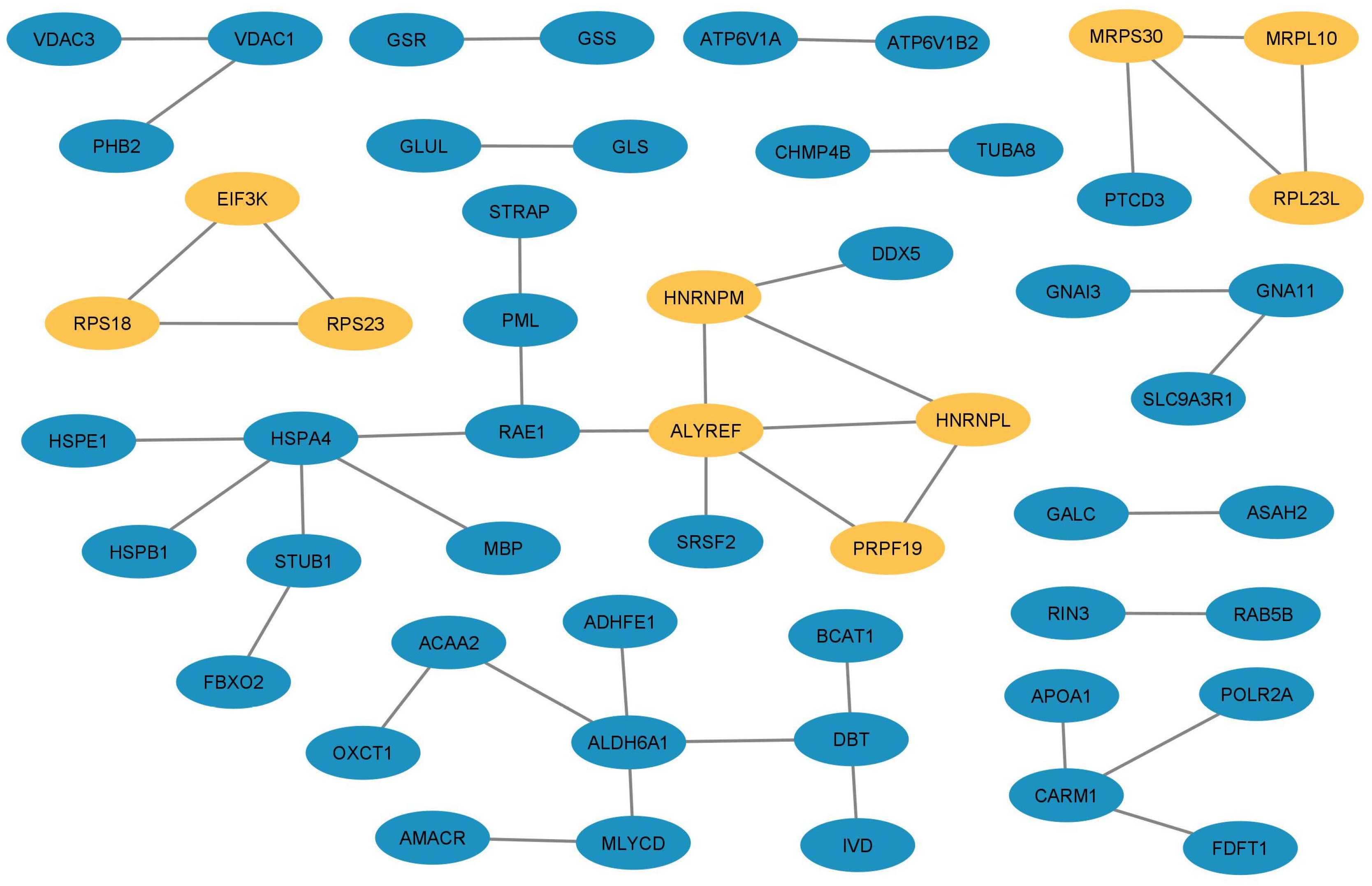
Among all the DEPs, we selected 10 overlapping hub proteins using the topological methods described in
Section 2.7 as potential key proteins in the triggering of seizures in the GASH/Sal strain (
Table 5).
ALYREF (THO Complex Subunit 4; DEP with 3.05 FC) is a nuclear protein that functions as a molecular chaperone. It is thought to regulate dimerization, DNA binding, and transcriptional activity of basic region-leucine zipper (bZIP) proteins (
https://www.proteinatlas.org/ENSG00000183684-ALYREF, accessed on 20 October 2022). Among the various functions of ALYREF, as in other RNA-binding proteins, there is direct involvement in the export of miRNAs, because it plays critical roles in nuclear export [
64,
65]. Altered microRNA expression has been observed in the brain and blood of patients with various epilepsy disorders. MicroRNAs are the main regulators of gene expression- because single miRNAs impact multiple proteins with diverse effects within different pathways and networks. Changes both in levels and activity of miRNAs can induce profound effects on cellular function. Regulation of miRNA has high potential use in complex disorders like epilepsy, where numerous cellular pathways and processes may be affected simultaneously [
66]. The understanding is that the miRNA–mRNA interactions will provide insights into epilepsy pathogenesis [
67]. This is because another emerging therapeutic target is miRNA-small noncoding RNAs, which negatively regulate sets of proteins. Therefore, further studies on the ALYREF protein are regarded as having the potential to provide insight into epilepsy and potential novel therapeutic strategies.
PRPF19 (pre-mRNA processing factor 19; overexpressed DEP with 2.29 FC) is a ubiquitin-protein ligase that is a core component of several complexes, mainly involved pre-mRNA splicing and DNA repair, which is required for pre-mRNA splicing as a component of the spliceosome. As a core component of the spliceosome, PRPF19 participates in its assembly and its remodeling and is required for its activity (
https://www.proteinatlas.org/ENSG00000110107-PRPF19, accessed on 21 October 2022). In addition, as part of the PSO4 core complex, this protein has been shown to participate in the DNA damage response. However, the specific role of the complex in DNA damage response pathways is still unclear [
68]. Moreover, studies on mouse brain development suggest that PRPF19 serves as a molecular switch in governing neuron/glia differentiation, inhibiting neuronal differentiation while accelerating the differentiation of astrocytes. However, the neuronal function of human PRPF19 has rarely been investigated, and its involvement in human neurological diseases remains unexplored [
69]. In conclusion, due to the vital role that PRPF19 plays in neural differentiation and in DNA damage and repair, it could be hypothetically related to alterations found in certain types of epilepsy. One example is epilepsy caused by focal cortical dysplasia, which originates from neurodevelopmental alterations.
HNRNPL (heterogeneous nuclear ribonucleoprotein L; overexpressed DEP with 2.02 FC) is a splicing factor binding to exonic or intronic sites and acting as either an activator or repressor of exon inclusion, exhibiting a binding preference for CA-rich elements. As part of a ribonucleoprotein complex that nucleates the complex on chromatin, it also negatively regulates the transcription of genes involved in neuronal differentiation (
https://www.uniprot.org/uniprotkb/F1LQ48/entry, accessed on 21 October 2022) [
70] or associated with alterations caused by trauma [
71]. Both phenomena are closely interlinked with the presence of certain types of epilepsy, which could establish a potential relationship between this disorder and this protein. In addition to the former, the role of HNRNPL in both calcium metabolism [
72] and potassium channel functioning [
73] might be relevant for its relationship with seizures and epilepsy.
HNRNPM (heterogeneous nuclear ribonucleoprotein M; overexpressed DEP with 2.05 FC) is other abundant nuclear protein that binds to pre-mRNA and is another component of the spliceosome complex. RNA metabolism involves complex and regulated processes, some of which include transcription, intracellular transport, translation, and degradation. The involvement of RNA binding proteins in these processes remains mostly uncharacterized. Regarding brain function, loss of HNRNPM affects the physiological spine in vivo by impairing the morphology of the dendritic spines in the hippocampus [
74]. Additionally, this protein directly binds to the 3’UTR of synaptophysin and PSD95 mRNAs, resulting in the stabilization of these mRNAs. This protein provides novel insight into the regulatory role of HNRNPM in neuronal structure and function [
74]. This suggests that a potential alteration of the HNRNPM hippocampus levels might be associated with altered synaptic processes in epilepsy.
MRPL10 (39S mitochondrial ribosomal protein L10; overexpressed DEP with 2.06 FC) is part of the mammalian mitochondrial ribosomes and play roles in the mitochondrial respiratory chain. MRPL10 is encoded by nuclear genes, is synthesized in the cytoplasm, and then is transported to the mitochondria to be assembled into mitochondrial ribosomes. Thus, MRPL10 is necessary for mitochondrial protein synthesis and mitochondrial activity since the deletion of
MRPL10 reduces the mitochondrial activity and expression of the mitochondrial complex [
75]. In addition, the deletion of this gene may negatively affect the adaptive metabolic response, cell proliferation, and cell survival due to inhibition of the kinase activity of cyclin-dependent kinase CDK1 [
75]. Interestingly, GWAS analysis in humans showed association of the
MRPL10 gene with cytokines, which are also associated with epilepsy [
76].
RPL23L (39S mitochondrial ribosomal protein L23; overexpressed DEP with 2.25 FC), also named MRPL23, is a structural constituent proteinic of ribosome and hence is involved in mitochondrial translation. This protein has been associated with the promotion of carcinoma metastasis and the occurrence of liver cancer [
77]; however, to our knowledge no direct or indirect association with seizures or/and epilepsy has been reported.
MRPS30 (mitochondrial ribosomal protein S30; overexpressed DEP with 2.52 FC) helps in protein synthesis within the mitochondrion. Mutations in its coding gene have been associated with mitochondrial diseases, such us Leigh syndrome, a rare mitochondrial disorder in which patients can develop epilepsy [
78]. Although epilepsy is a characteristic phenotypic feature of mitochondrial disorders, the prevalence of epilepsy is not equal among all patients with mitochondrial disorders [
79]. In the case of patients with the type of syndrome or others, such as West syndrome and epilepsia partialis continua, the incidence of epilepsy is very low [
80].
EIF3K (eukaryotic translation initiation factor 3 subunit K; overexpressed DEP with 2.11 FC) is the largest protein component of the eukaryotic translation initiation factor-3 complex, which is required for several steps in the initiation of protein synthesis [
81]. This protein was found to be a potential biomarker of schizophrenia in a PPI network study of differentially expressed genes [
82]; however, no association with epilepsy or seizures has been reported.
RPS23 (Ribosomal Protein S23; overexpressed DEP with 3.03 FC) is a large ribonucleoprotein complex and is a component of the ribosome responsible for the synthesis of proteins in the cell. This protein plays an important role in translational accuracy; missense mutations in its coding gene do not result in a reduction in the rate of mRNA translation but impair the accuracy of mRNA translation and render cells highly sensitive to oxidative stress [
83]. The genes encode ribosomal proteins, such as
RPS23, among others, which are enriched in biological processes of translation, translation elongation, and RNA processing in disc degeneration [
84].
RPS18 (40S ribosomal protein S18; overexpressed DEP with 2.05 FC) is a component of the 40S ribosomal subunit, and alterations in its expression level could affect translation rate through an effect on ribosome subunit concentrations.
Protein Outliers
Furthermore, we included in our analysis the five statistical protein outliers; since although they do not present multiple interactions with other proteins, they have a significant representation (over- or underexpressed DEPs) and, therefore, they must have some differential significance in the functionality of the IC.
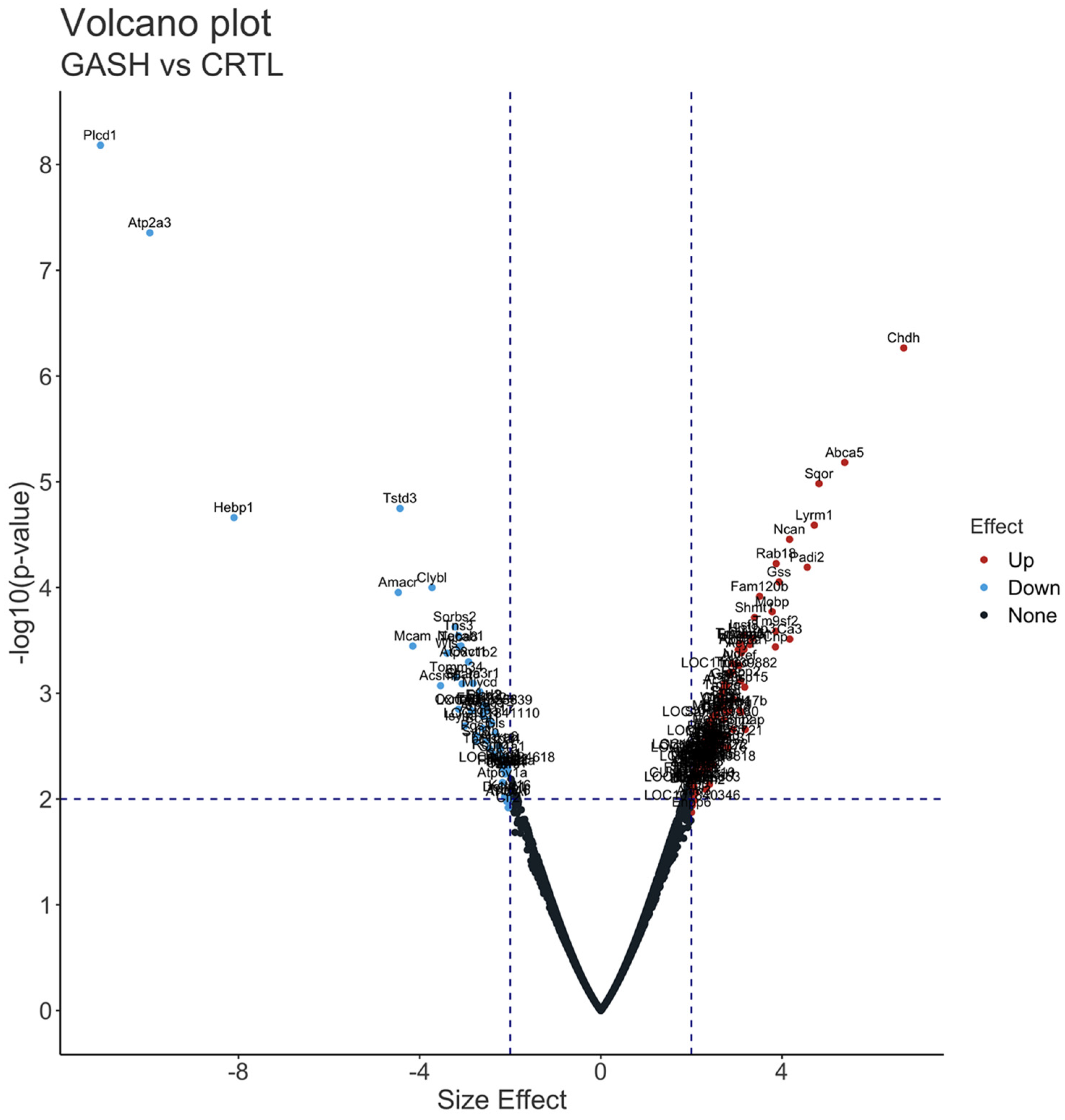
CHDH (choline dehydrogenase; overexpressed DEP with 6.68 FC) is a mitochondrial protein that regulates the concentrations of choline and glycine betaine in the blood and cells. Choline is important for regulation of gene expression as well as the biosynthesis of lipoproteins, membrane phospholipids, and the neurotransmitter acetylcholine [
85]; thus, variations of CHDH can affect susceptibility to choline deficiency. Moreover, a population-based study showed that the metabolic oxidation of choline is related to the risk of developing breast cancer [
86]. On the other hand, glycine betaine plays important roles as a primary intracellular osmoprotectant [
87], and impairments in human CHDH activity have been associated with various pathologies, including male infertility, homocystinuria, metabolic syndrome, cardiovascular diseases, and breast cancer [
87]. Despite all these data, at the moment, there is no association between this enzyme and epilepsy.
A type of ATP-binding cassette (ABC) transporter, part of a family of proteins associated with multidrug resistance, is another outlier overexpressed DEP. In particular, the ABCA5 (ATP-binding cassette sub-family A member 5; overexpressed DEP with 5.38 FC) was found to be overexpressed in the brain of patients with refractory epilepsy, suggesting an active drug efflux from brain [
88]. Moreover, ABCA5 was implicated in the neuropathology associated with Alzheimer’s and Parkinson’s disease [
89]. Alzheimer’s patients show specifically an increased expression of ABCA5 in hippocampal neurons, and amyloid-β-peptide levels are significantly reduced. Similar changes were observed with α-synuclein in neurons of the amygdala, where Parkinson’s disease patients showed higher levels of ABCA5. It is assumed that the overexpression of this protein is a protective response in these pathological conditions [
90].
In the case of three outlier underexpressed DEPs, the protein levels of the first of these, PLCD1 (phosphoinositide phospholipase C; underexpressed DEP with −11.04 FC), and its downstream factor transient receptor potential channel 4 (TRPC4), which colocalize with glutamatergic and GABAergic neurons, were elevated in focal cortical dysplasia type II and tuberous sclerosis complexes, which are well-known causes of chronic refractory epilepsy in children [
91]. Moreover, the lower levels of ATP2A3 (ATPase sarcoplasmic/endoplasmic reticulum Ca
2+ transporting 3; underexpressed DEP with −9.95 FC), an ATPase that transports Ca
2+ across membranes to the endoplasmatic reticulum to maintain a low cytoplasmic Ca
2+ level in neurons of substantia nigra pars compacta in Parkinson’s disease, indicates a deficit in organelle function and Ca
2+ sequestration [
92]. Thus, increased levels of cytoplasmic Ca
2+ due to lowered ATP2A3 levels could be detrimental to cells and cause degeneration [
93]. In the case of HEBP1 (heme-binding protein 1; underexpressed DEP with −8.09 FC), Yagensky et al. [
94] identified an increase of expression as a presymptomatic Alzheimer’s disease marker. This protein mediates heme-induced cytotoxicity via an apoptotic pathway, and interestingly, knockdown of
Hebp1 expression in neurons protects them from both heme and Ab42-induced apoptosis. Neurons lacking HEBP1 demonstrated resistance to apoptosis induced by hemin treatment. This resistance was due to the protein not being released into the cytosol, which prevented the activation of caspases 9 and 3/7, even though upstream activating events had occurred. These findings suggest that
HEBP1 may be involved in regulating the formation of apoptosomes, which are essential for cleaving procaspase 9 into its functional form.
Gene–Disease Association Analysis of DEPs
Finally, we included the potential functional significance of the 10 DEPs detected in the present work and directly related to epilepsy.
ATP6V1A (H
+-transporting two-sector ATPase; underexpressed DEP with −2.17 FC) is part of a multimeric complex present in a variety of cellular membranes that acts as an ATP-dependent proton pump and plays a key role in pH homeostasis and intracellular signaling pathways.
ATP6V1A variants, mainly clustering within the ATP synthase α/β family-nucleotide-binding domain, include early lethal encephalopathies and developmental encephalopathy in epilepsy [
95,
96,
97]. Interestingly, several mutants of
ATP6V1A, such as the p.Asp100Tyr, are characterized by reduced expression due to increased degradation; these mutations caused a defect in neurite elongation accompanied by loss of excitatory inputs, revealing that altered lysosomal homeostasis markedly affects neurite development and synaptic connectivity [
95]. Furthermore, Persike et al. [
98] used proteomics to determine the differential expression of proteins in the hippocampus of patients with mesial temporal lobe epilepsy (MTLE) compared to control samples, although in this case, ATP6V1A was up-regulated in comparison with our results. On the other hand, we also observed that ATP6V1B2, a subunit of the V-ATPase complex, was down-regulated. Pathogenic variants of
ATP6V1B2 have been associated with various epileptogenic phenotypes, including intellectual disability (ID), seizures, and/or DOORS syndrome (deafness, onychodystrophy, osteodystrophy, ID, and seizures) [
99,
100,
101,
102].
Alterations in the expression levels of BCAN (brevican core protein isoform X2; overexpressed DEP with 2.08 FC) and NCAN (neurocan core protein isoform X2; overexpressed DEP with 4.16 FC) have been reported in epileptic patients, as in the case of focal cortical dysplasia [
103,
104]. Epilepsies with a genetic basis can manifest early in life, and both
BCAN and
NCAN are believed to play crucial roles in terminal differentiation during development, as well as in the adult nervous system during postnatal development. Therefore, our data showing increased levels of expression of BCAN and NCAN in the GASH/Sal strain as compared with the control might explain an imbalance in the extracellular matrix composition that could be implicated in the origin or propagation of seizures.
The enzyme FAAH (fatty acid amide hydrolase), which was found to be overexpressed with a FC of 2.40 in the DEPs, is responsible for the breakdown of various primary and secondary fatty acid amides. This includes endocannabinoids such as anandamide, which have neuro-modulatory properties [
105]. This enzyme, together with 2-arachydonoil glycerol, acts as a key regulator of glutamate and GABA release, providing different forms of synaptic plasticity in excitatory and inhibitory neurotransmission in several brain regions [
106]. In summary, the overexpression of FAAH may lead to a reduced ability to protect against excessive neuronal activity, such as during epileptic seizures.
In addition, another DEP detected in our study is the GJA1 (gap junction protein; overexpressed DEP with 2.03 FC). This protein might be associated with epilepsy as it plays a significant role in phenomena related to cellular communications and provides a route for the diffusion of low-molecular-weight materials from cell to cell. In this regard, it is known that some of the proteins involved in cell-to-cell communication may represent potential targets in epilepsy treatment [
107]. Consistently, our results showed an upregulation of GJA1, which might indicate an abnormal flow of molecules between cells and might potentially contribute to seizure genesis and propagation in the GASH/Sal.
In the case of the enzyme GLS (glutaminase; underexpressed DEP with −2.33 FC), which is involved in the conversion of glutamine into glutamate, Rumping et al. [
108] identified two gene variants, a homozygous p.(Asp232Glufs*2) and compound heterozygous p.(Gln81* and Arg272Lys) in children with neonatal epileptic encephalopathy. These variants likely resulted in glutaminase deficiency, which led to an increase in glutamine levels in the affected children. Moreover, disturbed glutamine–glutamate shuttling is a known cause of epilepsy [
109,
110]. Thus, a loss of function in glutaminase can cause a neurometabolic disorder that leads to lethal early neonatal encephalopathy. Additionally, this disorder is likely to be associated with the epileptic phenotype observed in GASH/Sal hamsters.
GLS gene is highly expressed in the brain and has a pivotal role in creating glutamate abundance in this organ, contrary to the systemic circulation, in which glutamine is the most abundant amino acid [
111,
112].
Another DEP associated with the glutamate metabolism is GLUL (glutamate-ammonia ligase; overexpressed DEP with 2.01 FC). This protein regulates the levels of toxic ammonia and converts neurotoxic glutamate into harmless glutamine, where loss of GLUL in astroglia is reported in the hippocampi of epileptic patients [
113]. The two-fold increase in basal GLUL expression levels could indicate the need for greater availability of this enzyme by the GASH/Sal, a fact that can be correlated with elevated glutamate levels, which is widely described in epilepsy [
114,
115]. On the other hand, ammonium levels also modulate the occurrence of seizures in myoclonic epilepsy and various other related conditions [
116,
117]. GLUL is directly related to the detoxification process of both compounds mentioned above, and both could be found increased during and after convulsive crises, such as those described in the GASH/Sal model.
Furthermore, GSR (glutathione reductase; overexpressed DEP with 2.46 FC) is a central enzyme of cellular antioxidant protection that is closely linked to glutathione metabolism, reducing oxidized glutathione disulfide (GSSG) to the sulfhydryl form GSH, which is a valuable cellular antioxidant. In this sense, GSR maintains high levels of reduced glutathione in the cytosol. As occurred in the GLUL enzyme and other compounds involved in the metabolism of glutathione, an increase of GSR levels may also indicate the intensification of oxidative stress machineries as extensively reported in seizures [
118,
119]. Therefore, we hypothesized that elevated levels of the GSR enzyme could indicate the occurrence of compensatory mechanisms that help reduce harmful elements in the cell. This could be relevant to the seizure susceptibility of GASH/Sal, since noxious elements are generated within neurons, mainly due to the excitatory period they experience.
A type of heat shock protein (HSP) was detected within the DEPs in our study and related with epilepsy: HSPA4 (heat shock 70 kDa protein 4; underexpressed DEP with −2.20 FC). The HSP provides a first line of defense against accumulation of misfolded proteins as well as accomplishing other neuroprotective effects, e.g., inhibition of apoptosis, protection of cytoskeletal structures and immune modulation in stress conditions [
120]. Interestingly, it has also been reported that HSPA4, during the latency phase following status epilepticus, shows a significant upregulation in subregions of the temporal lobe crucial for the development of TLE [
121] and in a pilocarpine model of this type of epilepsy [
122]. It is, therefore, plausible that cytomolecular changes in the neuronal circuitry during epileptogenesis lead to increased endoplasmic reticulum stress with following upregulated expression of HSPH4, which would exert protective effects as a response to stress factors [
123]. Low levels of this enzyme would make GASH/Sal more vulnerable to convulsive episodes. However, it would be necessary to study the modifications of this protein after the convulsive crises.
Finally, in the present work we detected several DEPs linked to the functioning of glial cells, such as the myelin basic protein (MBP; overexpressed DEP with 2.09 FC), the most abundant protein component of the myelin membrane in the CNS. MBP has a key role in both myelin formation and its stabilization; however, studies that discuss the link between MBP and epilepsy are scarce. Thus, for example, children with seizures had normal MBP levels in cerebral spinal fluid, similar to the controls; that is, for individual patients, MBP is of little value as a prognostic indicator [
124]. Another study reported that up-regulation of MBP protein levels in zinc diet-treated animals with seizures may represent a compensatory mechanism for neuronal membrane damage and repair [
125]. Nevertheless, a decrease of MBP levels in the epileptic foci of patients suffering from intractable epilepsy is reported [
126]. We are currently unable to provide a plausible explanation for the result observed in MBP. Thus, it is essential to conduct further studies on the glial population in GASH/Sal hamsters to obtain additional information regarding this under-explored field in this experimental model of epilepsy.

 ,
, 





{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}